- Osmundaceae
-
?Osmundáceas

Clasificación científica Reino: Plantae Clados superiores: Plantae (clado) Viridiplantae
Streptophyta
Streptophytina
Embryophyta
Tracheophyta
Euphyllophyta
MonilophytaClase: Polypodiopsida Orden: Osmundales Familia: Osmundaceae genera - Osmundastrum
- Osmundastrum cinnamomeum (=Osmunda cinnamomea)
- Leptopteris
- Todea
- Osmunda sensu stricto
Osmundales y su única familia Osmundaceae es el más pequeño pero más anciano orden de helechos leptosporangiados (Polypodiopsida) y ocupa una importante posición filogenética como hermano de todo el resto de leptosporangiados vivientes (Hasebe et al. 1995,[1] Pryer et al. 2004,[2] Schuettpelz y Pryer 2007[3] ). Osmundaceae se caracteriza por poseer rizomas con una distintiva anatomía en el corte transversal, consistente en toda la familia y única en los helechos (Hewitson 1962,[4] Gifford y Foster 1989[5] p. 267), esporangios que no están organizados en soros, esporas verdes, y un único set de caracteres reproductivos que parecen intermedios entre los helechos leptosporangiados y los eusporangiados (Kramer 1990[6] ): los leptosporangios de Osmundaceae se desarrollan de células iniciales múltiples y producen cientos de esporas, ambos caracteres asociados al desarrollo de los helechos eusporangiados (Bierhost 1971,[7] Ogura 1972[8] ). Los esporangios también tienen un característico anillo como un parche o escudo, que causa una dehiscencia longitudinal, distinta de todas las demás morfologías de anillo presentes en los demás helechos leptosporangiados (Bierhost 1971[7] ).
Osmundaceae fue largamente reconocido como monofilético. De climas templados y tropicales, su registro fósil comienza en el Pérmico. Poseen poca importancia económica, su importancia es sistemática.
Contenido
Caracteres
 Diagrama esquemático del ciclo de vida de las plantas vasculares sin semilla.
Diagrama esquemático del ciclo de vida de las plantas vasculares sin semilla.
Referencias:
n : generación haploide,
2n : generación diploide,
m! : mitosis,
M! : meiosis,
F! : fecundación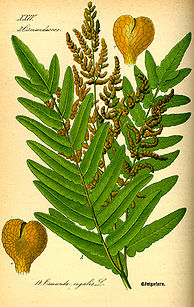 Ilustración de Osmunda regalis. Obsérvese el anillo lateral (escudo de dehiscencia) en el esporangio.
Ilustración de Osmunda regalis. Obsérvese el anillo lateral (escudo de dehiscencia) en el esporangio.Las osmundáceas son plantas vasculares con ciclo de vida haplodiplonte donde la alternancia de generaciones es bien manifiesta, con esporófito y gametofito multicelulares e independientes, con esporas como unidad de dispersión y de resistencia. El gametofito es un "talo" (cuerpo sin organización), y el esporofito es un "cormo" (con raíz, vástago y sistema vascular). Debido a estas características tradicionalmente se las agrupa con las "pteridofitas".
Poseen esporofitos con megafilos o "frondes" (Euphyllophyta).
La anatomía del tallo es muy característica: la estela es una sifonostela ectofloica, con un anillo de hebras de xilema discretas, que normalmente aparecen conduplicadas o dos veces conduplicadas en un corte transversal.
Hojas con estípulas en la base de los pecíolos. Las hojas son dimórficas o con porciones fértiles distintas de las estériles (véase ilustración).
Esporangios no organizados en soros. Los esporangios son leptosporangios pero a diferencia de los leptosporangios del resto de las familias de helechos leptosporangiados, se inician de células iniciales múltiples en lugar de una sola, y producen cientos de esporas, ambos caracteres típicos de los eusporangios. Los esporangios son grandes, con 128-512 esporas cada uno. El anillo es un anillo lateral, con forma de parche o escudo, a veces llamado "escudo de dehiscencia", que causa una dehiscencia longitudinal por una hendidura apical (véase ilustración).
Esporas verdes, subglobosas, con marca trilete.
Gametofitos grandes, verdes, cordados, superficiales.
Número de cromosomas: x = 22.
Ecología y Evolución
El registro fósil comienza en el Pérmico, donde se encuentran numerosos representantes (Tidwell y Ash 1994[9] ), y representantes del género Osmunda son conocidos desde el Triásico (Phipps et al. 1998,[10] Vavrek et al. 1996[11] ).
De climas templados y tropicales.
Filogenia
Monofilético (Hasebe et al. 1995,[1] Yatabe et al. 1999,[12] Pryer et al. 2001a[13] y 2004b[2] ).
Un análisis de ADN sobre la secuencia rbcL de Osmundaceae encontró que Osmunda cinnamomea es hermana de todo el resto de la familia, incluyendo a Leptopteris y Todea (Yatabe et al. 1999[12] ). Este estudio también encontró que Osmunda claytoniana es hermana de un clado conteniendo al subgénero Plenasium y al subgénero Osmunda. Si bien esta relación no tenía mucho sostén, Yatabe et al. (2005[14] ) propusieron un nuevo subgénero, Claytosmunda Yatabe, Murakami & Iwatsuki, para acomodarlo a la posición filogenética de O. claytoniana.
Osmunda en sentido amplio es parafilético (Metzgar et al. 2008[15] ) y hoy se ha dividido en los géneros Osmundastrum y Osmunda en sentido estricto (sensu stricto) tal como lo proponía Yatabe et al. (2005[14] ).
Taxonomía
Ubicación taxonómica: Plantae (clado), Viridiplantae, Streptophyta, Streptophytina, Embryophyta, Tracheophyta, Euphyllophyta, Monilophyta, Clase Polypodiopsida, Orden Osmundales, familia Osmundaceae.
Numerosos tratamientos taxonómicos fueron recomendados para Osmundaceae (Hewitson 1962,[4] Miller 1971,[16] Kramer 1990,[6] Yatabe et al. 2005[14] ).
Sensu Smith et al. 2006 son 3 géneros, cerca de 20 spp.:
- Leptopteris C. Presl.
- Todea Willd. ex Bernh.
- Osmunda en 3 subgéneros (Kramer 1990[6] ): subg. Osmunda L. con 3 especies, subg. Osmundastrum (C. Presl.) C. Presl. con 2 especies, y subg. Plenasium (C. Presl.) J. Smith con 3 o 4 especies.
Sensu Yatabe et al. (2005[14] ) son 4 géneros, avalados por los actuales conocimientos de filogenia (Metzgar et al. 2008[15] ):
- Leptopteris C. Presl.
- Todea Willd. ex Bernh.
- Osmundastrum que en la clasificación anterior se ubica dentro de Osmunda, con su única especie, O. cinnamomeum.
- Osmunda con 3 subgéneros: subg. Claytosmunda con su especie O. claytoniana, subg. Plenasium y subg. Osmunda.
Leptopteris y Todea comparten muchos caracteres, entre los que se encuentran las hojas monomórficas y los esporangios que siguen a las venas en pinas fértiles no contráctiles (Kramer 1990[6] ), pero los dos géneros son distinguibles.
Leptopteris, con unas 6 especies, tiene hojas tipo film que no tienen estomas, y pocos esporangios desparramados en la superficie abaxial.
Todea, con 2 especies, tiene hojas coriáceas con estomas, y esporangios cubriendo densamente la superficie abaxial (Hennipman 1968,[17] Brownsey 1981[18] ).
Osmunda en sentido amplio (sensu lato) se ha distinguido de estos dos géneros por sus pinas fértiles contraídas y contiene 8 a 9 especies.
-

Osmunda regalis Pinas fértiles contraídas.
-

Osmunda claytoniana Pinas fértiles contraídas.
Importancia económica
Poseen poca importancia económica, su importancia es sistemática.
Algunas especies de Osmunda se cultivan como ornamentales.
La masa densa de raíces que forman los helechos del género Osmunda se usa para producir "fibra de Osmunda", que es empleada como soporte de crecimiento de orquídeas cultivadas y otras plantas epífitas.
Véase también
- Clasificación de los organismos vegetales
- Para ver una lista completa de los taxones ver Sistema de clasificación de monilophytas de Smith 2006
- Para una descripción básica de morfología y anatomía de los helechos ver Pteridophyta
Enlaces externos
- Stevens, P. F. (2001 en adeltante) Angiosperm Phylogeny Website Versión 8, junio de 2007 y actualizado desde entonces. http://www.mobot.org/MOBOT/research/APweb/ Fecha de acceso: 5 de mayo del 2008.
- Pryer's lab con los pdfs de las publicaciones de la autora.
 Wikimedia Commons alberga contenido multimedia sobre Osmundaceae. Commons
Wikimedia Commons alberga contenido multimedia sobre Osmundaceae. Commons Wikiespecies tiene un artículo sobre Osmundaceae. Wikispecies
Wikiespecies tiene un artículo sobre Osmundaceae. Wikispecies
Bibliografía
- A. R. Smith, K. M. Pryer, E. Schuettpelz, P. Korall, H. Schneider, P. G. Wolf. 2006. "A classification for extant ferns". Taxon 55(3), 705-731 (pdf aquí)
Lecturas recomendadas
- Pryer, Kathleen M., Harald Schneider, Alan R. Smith, Raymond Cranfill, Paul G. Wolf, Jeffrey S. Hunt y Sedonia D. Sipes. 2001. "Horsetails and ferns are a monophyletic group and the closest living relatives to seed plants". Nature 409: 618-622 (resumen en inglés aquí).
- Pryer, Kathleen M., Eric Schuettpelz, Paul G. Wolf, Harald Schneider, Alan R. Smith y Raymond Cranfill. 2004. "Phylogeny and evolution of ferns (monilophytes) with a focus on the early leptosporangiate divergences". American Journal of Botany 91:1582-1598 (resumen en inglés aquí).
Referencias citadas
- ↑ a b Hasebe, M.; P. G. Wolf, K. M. Pryer, K. Ueda, M. Ito, R. Sano, G. J. Gastony, J. Yokoyama, J. R. Manhart, N. Murakami, E. J. Crane, C. H. Hayfler, y W. D. Hauk. (1995). «Fern phylogeny based on rbcL nucleotide sequences.». American Fern Journal 85: pp. 134-181.
- ↑ a b Pryer, K.M.; E. Schuettpelz, P.G. Wolf, H. Schneider, A.R. Smith, R. Cranfill (2004). «Phylogeny and evolution of ferns (monilophytes) with a focus on the early leptosporangiate divergences.» (pdf). American Journal of Botany 91: pp. 1582-1598. http://www.pryerlab.net/publication/fichier494.pdf. Consultado el 03/05/2008.
- ↑ Schuettpelz E.; K.M. Pryer (2007). «Fern phylogeny inferred from 400 leptosporangiate species and three plastid genes.» (pdf). Taxon 56: pp. 1037-1050. http://www.pryerlab.net/publication/fichier1085.pdf. Consultado el 03/05/2008.
- ↑ a b Hewitson, W. (1962). «Comparative morphology of the Osmundaceae.». Annals of the Missouri Botanical Garden 49: pp. 57-93.
- ↑ Gifford, E. M.; Foster, A. S. (1989). Morphology and evolution of vascular plants. Ed. 3.. New York: W. H. Freeman & Co..
- ↑ a b c d Kramer, K. U. (1990). K. Kubitzki. ed. The families and genera of vascular plants vol. 1.. Berlin: Springer Verlag.
- ↑ a b Bierhost, D. W. (1971). Morphology of vascular plants.. London: The Macmillan Company.
- ↑ Ogura, Y. (1972). Comparative anatomy of vegetative organs of the pteridophytes. Handbuck der Pflanzanatomie vol. 7, part 3.. Berlin: Gebrüder Borntraeger.
- ↑ Tidwell, W. D.; Ash, S. R. (1994). «A review of selected Triassic to early Cretaceous ferns.». Journal of Plant Research 107: pp. 417-442.
- ↑ Phipps, C. J.; T. N. Taylor, E. L. Taylor, N. R.Cuneo, L. D. Boucher, y X. Yao. (1998). «Osmunda (Osmundaceae) from the Triassic of Antarctica: an example of evolutionary stasis.». American Journal of Botany 85: pp. 888-895.
- ↑ Vavrek, M. J.; R. A. Stockey, y G. W. Rothwell (2006). «Osmunda vancouverensis sp. nov. (Osmundaceae), permineralizad fertile frond segments from the Lower Cretaceous of British Columbia, Canada.». International Journal of Plant Sciences 167: pp. 631-637.
- ↑ a b Yatabe, Y.; H. Nishida, y N. Murakami. (1999). «Phylogeny of Osmundaceae inferred from rbcL nucleotide sequences and comparison to the fossil evidences.». Journal of Plant Research 112: pp. 397-404.
- ↑ Pryer, K.M.; H. Schneider, A.R. Smith, R. Cranfill, P.G. Wolf, J.S. Hunt and S.D. Sipes (2001). «Horsetails and ferns are a monophyletic group and the closest living relatives to seed plants.». Nature 409: pp. 618-622.
- ↑ a b c d Yatabe, Y.; N. Murakami, y K. Iwatsuki (2005). «Claytosmunda, a new subgenus of Osmunda (Osmundaceae).». Acta Phytotaxonomica et Geobotanica 56: pp. 127-128.
- ↑ a b Metzgar, J. S.; Skog, J. S., Zimmer, E. A., Pryer, K. A. (2008). «The paraphyly of Osmunda is confirmed by phylogenetic analyses of seven plastid loci.» (pdf). Systematic Botany 33 (1): pp. 31-36. http://www.pryerlab.net/publication/fichier1090.pdf. Consultado el 03/05/2008.
- ↑ Miller, C. N. Jr. (1971). «Evolution of the fern family Osmundaceae based on anatomical studies.». Contributions from the Museum of Paleontology, University of Michigan 23: pp. 105-169.
- ↑ Hennipman, E. (1968). «A new Todea from New Guinea with rmarks onthe generic delimitation in recent Osmundaceae (Filices).». Blumea 16: pp. 105-108.
- ↑ Brownsey, P. J. (1981). «A biosystematic study of a wild population of Leptopteris hybrids in New Zealand.». New Zealand Journal of Botany 19: pp. 343-352.
Categorías:- Pteridopsida
- Polypodiopsida
- Familias de plantas
- Osmundastrum




Wikimedia foundation. 2010.