- Sonic hedgehog
-
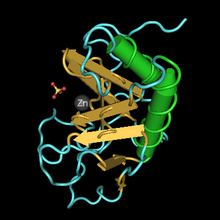 Estructura tridimensional del dominio de señalización de Sonic hedgehog
Estructura tridimensional del dominio de señalización de Sonic hedgehog
El homólogo Sonic hedgehog (Shh) es una de las tres proteínas de la familia hedgehog. SHH es el ligando mejor estudiado de la vía de señalización hedgehog. Juega un papel esencial en la regulación de la organogénesis de los vertebrados, como el crecimiento de las falanges de las extremidades y la organización del cerebro. Sonic hedgehog es un ejemplo canónico de un morfógeno tal como fue definido por el modelo de la bandera francesa de Lewis Wolpert: una molécula que difunde para formar un gradiente de concentración y tiene diferentes efectos en las células del embrión en desarrollo dependiendo de su concentración. Shh sigue siendo importante en el adulto, controlado la división celular de células madre adultas y está implicado en el desarrollo de ciertos cánceres.
Familia Hedgehog
El gen Drosophila hedgehog fue bautizado de tal forma puesto que su perdida de función causa un fenotipo cubierto por dentículos puntiagudos en las moscas de las frutas, similares a las de un erizo. Este gen tiene tres homólogos en vertebrados. Los dos primeros, Desert hedgehog (Dhh), e Indian hedgehog (Ihh), fueron nombrados según especies de erizos existentes. Dhh se expresa en las células de Sertoli en los testículos, e Ihh se expresa a su vez en el cartílago y es importante para el crecimiento de hueso postnatal (Bitgood & McMahon, 1995; Bitgood et al, 1996). Sonic hedgehog (Shh), nombrado en honor al personaje de videojuegos de Sega Genesis, es el homólogo de hedgehog en vertebrados con mayor número de funciones, principalmente en el desarrollo embrionario. Las proteínas de la familia hedgehog son factores paracrinos con actividades morfogenicas. En el embrión inducen la diferenciación celular, y generan barreras entre tejidos. Para que una proteína hedgehog sea funcional debe estar acoplada a una molécula de colesterol. Esta unión tiene como resultado adicional, una mayor capacidad de difusión entre tejidos (Lewis et al, 2001).
Funciones
Neurulación
Shh es esencial para la formación del tubo neural en humanos y otros vertebrados, junto con genes como Pax3 y openbrain. El tubo neural debe polarizarse dorso-ventralmente para dar origen a dos tipos de neuronas: por una lado las neuronas espinales, que reciben señales de las neuronas sensoriales, situadas en la parte dorsal, y por otro lado las neuronas motrices situadas en la parte ventral. El patrón ventral esta determinado por la secreción de la proteína sonic hedgehog en el notocordio. A su vez, el patrón dorsal esta determinado por la secreción en el ectodermo de proteínas de la familia TGF-β como BMP4, BMP7, BMP5, Dorsalin y Activin. Los gradientes formados por estos factores paracrinos determinan la identidad de las células del tubo: dorsalmente se forma la placa del techo y ventralmente la placa del piso (Roelink et al, 1995; Briscoe et al, 1999). La importancia de la secreción de la proteína sonic hedgehog por el notocordio puede ser demostrada por el trasplante de éste último, a los lados del tubo neural. Las células del tubo neural adyacentes al notocordio trasplantado, toman la identidad de las células de la placa del piso (Placzek et al, 1990; Echelard et al, 1993). Adicionalmente, la expresión de Sonic hedgehog en la placa del piso juega un papel importante en la atracción de las neuronas comisurales a la línea media ventral (Charron et al, 2003).
Especificación de las neuronas motrices
Para que la especificación de las neuronas motrices se lleve a cabo se tienen que dar dos procesos. El primero es regulado por la secreción de Sonic hedgehog en el notocordio, y consiste en la especificación de las neuronas del margen ventrolateral en neuronas ventrales. El segundo paso, regulado igualmente por la secreción de Shh, esta vez en la placa del piso del tubo neural, indica a las neuronas ventrales que se conviertan en neuronas motrices, y no en interneuronas (Ericson et al, 1992). Shh puede por lo tanto inducir a células madre neurales obtenidas mediante clonaje terapéutico, a diferenciarse en células madre neurales ventrales.
Crecimiento cerebral
Alteraciones en el gen Shh, junto con genes como ASPM, NCAM y microcephalin, pueden afectar el crecimiento del cerebro en humanos y otros vertebrados causando microcefalia, ya que se expresan en zonas donde ocurre la neurogénesis (Bond et al. 2002). Estos genes han sido seleccionados positivamente durante la evolución de los humanos, dando como resultados mayor crecimiento del cerebro en comparación a otros primates como chimpancés, orangutanes y gorilas (Koupirna et al. 2004).
Apoptosis
El fenómeno de muerte celular programada, apoptosis, estaá mediado por proteasas llamadas caspasas. Éstas digieren la célula desde el interior, cortando proteínas celulares y fragmentando el ADN. En el tubo neural del pollo, la proteína receptora de hedgehog Patched, activa las caspasas. La unión de sonic heghog suprime patched y por lo tanto las caspasas no se activan, inhibiendo así el proceso de apoptosis (Thibert et al, 2003). Receptores dependientes como patched previenen la formación de células neurales fuera de los tejidos indicados, y la ausencia de dichos receptores esta asociada a cánceres (Porter & Dhakshinamoorty, 2004).
Desarrollo de los dientes
El nudo de enamel, constituye el centro de señalización de los dientes. Su formación es inducida en el epitelio por la mesenquima derivada de la cresta neural. Este nudo secreta una gran variedad de factores paracrinos, entre los cuales están Shh, BMP2, BMP4, BMP7, y FGF4. Shh y FGF4 inducen la proliferación de las células para formar las cúspides (Jernvall et al, 1994; Vaahtokari et al, 1996). Los BMPs se autorregulan, y regulan la producción de sus inhibidores generando un efecto de bloqueo en la proliferación epitelial. Shh y los FGFs tienen una actividad inhibitoria sobre los BMPs. El patrón de actividad localizada de estos factores, genera la gran diversidad de formas y cambios en los dientes de diferentes especies (Salazar-Ciudad & Jernvall, 2004).
Determinación del eje derecha-izquierda
Durante el desarrollo del pollo, Shh y chordin se expresan de manera localizada en la parte anterior de la banda primitiva y en el nódulo de hensen. Cuando la banda primitiva alcanza su máxima longitud, Shh deja de expresarse en el lado derecho del embrión inhibido por la expresión de Activin B, quién a su vez activa la expresión de FGF8. En el lado izquierdo del embrión Shh activa la expresión de Cerberus y de Nodal. Este proceso determina la formación del eje derecha-izquierda (Levin et al, 1995).
Osificación endocondral
La osificación endocondral, un proceso que se da en la columna, costillas, pelvis y extremidades, involucra la formación de tejido cartilaginoso a partir e células mesenquimales y el posterior remplazo del cartílago por hueso. Sonic hedgehog genera la diferenciación de células mesenquimales en células cartilaginosas, mediante la inducción de la expresión de Pax1 en las células esclerotomales adyacentes (Csejersi et al, 1995).
Desarrollo de extremidades
Durante la formación de las extremidades, una zona específica del mesodermo determina la identidad antero-posterior de la extremidad: la zona de actividad polarizante (ZPA). Esta zona esta determinada por la expresión de Shh. Los factores de trascripción dHAND, Hoxd12 y Hoxb8 operan conjuntamente para activar la expresión de Shh y limitarla a la mesénquima posterior (Lettice et al, 2003). Las células de la ZPA donde se secreta Shh, no sufren apoptosis como las células de la cresta ectodérmica apical (AER), otra zona importante en la determinación de la identidad de la extremidad, sino que pasan a formar el hueso, la piel y el músculo de la extremidad. En embriones normales, el gen Shh es activado por los FGFs presentes en el cogollo de la pierna, mientras que los genes de FGF en la AER son activados por Shh. La proteína Shh, induce a Gremlin, un inhibidor de BMPs, previniendo así que los BMPs inhiban los FGFs de la AER (Zuñiga et al, 1999). Esto quiere decir, que una vez se han establecido la ZPA y la AER, estos genes se soportan mutuamente (Niswander et al, 1994). La retroalimentación positiva generada, se detiene una vez que las células de la ZPA forman una barrera entre las células que secretan Shh y las que secretan Gremlin para mantener la AER (Scherz et al, 2004). Actuando típicamente como un morfógeno, Shh puede determinar la identidad de los dedos que se forman en la extremidad, dependiendo de su concentración y tiempo de expresión en cada dedo (Ahn & Joyner, 2004; Harfe et al, 2004). El trasplante de la ZPA, o de células que secretan Shh al margen anterior del cogollo de la extremidad en formación conlleva a la formación de un patrón simétrico de expresión de Hoxd, y una duplicación simétrica en el número de dedos (Riddle et al, 1993). En la extremidad amputada de una salamandra, shh se expresa en el blastema, al igual que se expresaba en la formación de la extremidad (Imokawa & Yoshizato, 1997; Torok et al, 1999).
Desarrollo del intestino
El intestino sufre un proceso de especialización regional durante el desarrollo del pollo. Los límites entre regiones están dados por la interacción con el mesodermo. Cuando el tubo intestinal empieza a formarse en los extremos anterior y posterior, induce la especialización regional del mesodermo esplácnico, donde Shh está involucrado. En las etapas mas tempranas del desarrollo la expresión de Shh esta limitada al intestino posterior y a la faringe. A medida que el tubo intestinal se extiende hacia el centro del embrión, el área de expresión de Shh se extiende de igual manera. Su expresión desigual a través de todo el tubo delimita las zonas del mesodermo que rodea el intestino (Roberts et al, 1995).
Determinación del esclerotoma
La especialización de los somitas se lleva a cabo por la interacción entre varios tejidos. La parte ventral media del somita, es inducida a diferenciarse en esclerotoma por factor paracrinos entre los cuales se destaca Sonic hedgehog. Este es secretado por el notocordio y la placa del piso del tubo neural (Fan & Tessier-Lavigne 1994; Johnson et al, 1994). El esclerotoma dará origen posteriormente a partes de la columna vertebral y la mayoría del cráneo.
Desarrollo del páncreas
Mientras que la presencia de células cardíacas induce la formación del hígado, la presencia de células del notocordio la previene. El caso contrario se da durante el desarrollo del páncreas. La presencia del notocordio y la ausencia del corazón generan la formación del páncreas a partir del intestino. Esta activación se da por que el notocordio inhibe de la expresión de Shh en el endodermo (Appelqvist et al, 1997; Hebrok et al, 1998). Shh se expresa a lo largo de todo el endodermo intestinal, excepto en la región del páncreas. El notocordio secreta FGF2 y activin que inhiben a shh. Si se induce la expresión de shh experimentalmente, la formación del páncreas no se da y el tejido sigue siendo tejido intestinal (Offield et al, 1996; Jonnson et al, 1994; Ahlgren et al, 1996).
Desarrollo de ojos
La formación de dos ojos bilaterales a partir de el ojo central primordial, depende de la expresión de Shh. Éste tiene una acción antagónica al gen Pax6. De ser inhibido Shh, el ojo central no se separa resultando en la condición conocida como ciclopía: un único ojo ubicado generalmente bajo la nariz. Por el contrario una sobreexpresión de Shh suprime demasiado a Pax6, impidiendo la formación de los ojos (Chiang et al, 1996; kelley et al, 1996;Roessler et al, 1996). Este es el fenómeno que lleva al arresto de la formación de los ojos en las poblaciones troglófilas de Astayanax mexicanum (Yamamoto et al. 2004).
Desarrollo de plumas y escamas
Shh y BMP2 se expresan durante la formación de las escamas en reptiles y de las plumas aves. El cambio que pudo haber dado origen a las plumas a partir de un ancestro reptil con escamas, es la localización de la expresión de Shh y BMP2 en la parte distal de la estructura, estimulando así la proliferación de células mesenquimales en el eje vertical de la pluma, para formar el raquis. (Harris et al, 2002; Yu et al, 2002).
Metamorfosis en ranas
Durante la metamorfosis de las ranas, el intestino de los renacuajos adaptado a una dieta herbívora, debe acortarse para adaptarse a una dieta carnívora. La absorción de las células en exceso y la reprogramación de las células restantes esta mediada por la acción de la metaloproteinasa stromelysin-3 y por la transcripción de BMP4 y Shh (Stolow & Shi, 1995).
Referencias
• Ahlgren, U., J. Jonnson and H. Edlund. 1996. The morphogenesis of the pancreatic mesenchyme is uncoupled from that of the pancreatic epithelium in IPF/PDX1-deficient mice. Development 122: 1409– 1416.
• Ahn, S. and A. L. Joyner. 2004. Dynamic changes in the response of cells to positive hedgehog signaling during mouse limb patterning. Cell 118: 505–616.
• Apelqvist, A., U. Ahlgren and H. Edlund. 1997. Sonic hedgehog directs _emodeling_ mesoderm differentiation in the intestine and pancreas. Curr. Biol. 7: 801–804.
• Bitgood, M. J. and A. P. McMahon. 1995. Hedgehog and BMP genes are coexpressed at many diverse sites of cell-cell interaction in the mouse embryo. Dev. Biol. 172:126–158.
• Bitgood, M. J., L. Shen and A. P. McMahon. 1996. Sertoli cell signalling by desert hedgehog regulates the male germline. Curr. Biol. 6: 298–304.
• Bond, J. and 11 others. 2002. ASPM is a major determinant of cerebral cortical size. Nature Genet. 32: 316–320.
• Briscoe, J., L. Sussel, D. Hartigan-O’Connor, T. M. Jessell, J. L. R. Rubenstein and J. Ericson. 1999. Homeobox gene Nkx2.2 and specification of neuronal identity by graded Sonic hedgehog signaling. Nature 398: 622–627.
• Charron, F., E. Stein, J. Jeong, A. P. McMahon, and M. Tessier-Lavigne. 2003. The morphogen sonic hedgehog is an axonal chemoattractant that collaborates with Netrin-1 in midline axon guidance. Cell 113: 11–23.
• Chiang, C., Y. Litingung, E. Lee, K. K. Young, J. E. Corden, H. Westphal and P. A. Beachy. 1996. Cyclopia and defective axial patterning in mice lacking sonic hedgehog gene function. Nature 383: 407– 413.
• Cserjesi, P. and 7 others. 1995. A basic helix-loop-helix protein that prefigures skeletal formation during mouse embryogenesis. Development 121: 1099–1110.
• Echelard, Y., D. J. Epstein, B. St.-Jacques, L. Shen, J. Mohler, J. A. McMahon and A. McMahon. 1993. Sonic hedgehog, a member of a family of putative signaling molecules, is implicated in the regulation of CNS polarity. Cell 75: 1417–1430.
• Ericson, J., S. Morton, A. Kawakami, H. Roelink and T. M. Jessell. 1996. Two critical periods of Sonic hedgehog signaling required for the specification of motor neuron identity. Cell 87: 661–673.
• Fan, C. M. and M. Tessier-Lavigne. 1994. Patterning of mammalian somites by surface ectoderm and notochord: Evidence for sclerotome induction by a hedgehog homolog. Cell 79: 1175–1186.
• Hall, T.M., Porter, J.A., Beachy, P.A., Leahy, D.J. A potential catalytic site revealed by the 1.7-A crystal structure of the amino-terminal signalling domain of Sonic hedgehog. Nature v378 pp.212-216, 1995. • Harfe, B. D., P. J. Scherz, S. Nissim, H. Tian, A. P. McMahon,and C. J. tabin. 2004. Evidence for an expansion-based tempral Shh gradient in specifying vertebrate digit identities. Cell 118: 517–528.
• Harris, M. P., J. F. Fallon and R. O. Prum. 2002. Shh-Bmp2 signaling module and the evolutionary origin and diversification of feathers. J. Exp. Zool. 294: 160–176.
• Hebrok, M., S. Kim and D. A. Melton. 1998. Notochord repression of endodermal sonic hedgehog permits pancreas development. Genes Dev. 12: 1705–1713.
• Imokawa, Y. and K. Yoshizato. 1997. Expression of sonic hedgehog gene in regenerating newt limb blastema recapitulates that in developing limb buds. Proc. Natl. Acad. Sci. USA 94: 9159–9164.
• Jernvall, J., P. Kettunen, I. Karavanova, L. B. Martin and I. Theseleff. 1994. Evidence for the role of the enamel knot as a control center in mammalian tooth cusp formation: Non-dividing cells express growth stimulating Fgf-4 gene. Int. J. Dev. Biol. 38: 463–469.
• Johnson, R. L., E. Laufer, R. D. Riddle and C. Tabin. 1994. Ectopic expression of Sonic hedgehog alters dorsal-ventral patterning of somites. Cell 79: 1165–1173.
• Jonnson, J., L. Carlsson, T. Edlund and H. Edlund. 1994. Insulin-promote-factor 1 is required for pancreas development in mice. Nature 371: 606–608.
• Kelley, R. I. and 7 others. 1996. Holoprosencephaly in RSH/Smith-Lemli-Opitz syndrome: Does abnormal cholesterol metabolism affect the function of Sonic hedgehog? Am. J. Med. Genet. 66: 478–484.
• Kouprina, N. and 12 others. 2004. Accelerated evolution of the ASPM gene controlling brain size begins prior to human brain expansion. PloS Biol. 2: 0653–0663.
• Levin, M. 2003. Motor protein control of ion flux is an early step in embryonic left-right asymmetry. BioEssays 25: 1002–1010.
• Lettice, L. A., Heaney, S. J. H., Purdie, L. A., de Beer, P., Oostra, B. A., Goode, D., Elgar, G., Hill, R. E. and de Graaff, E. 2003. A long-range Shh enhancer regulates expression in the developing limb and fin and is associated with preaxial polydactyly. Human Molecular Genetics 12: 1725-1735.
• Lewis, P. M., M. P. Dunn, J. A. McMahon, M. Logan, J. F. Martin, B. St.-Jacques and A. P. McMahon. 2001. Cholesterol modification of Sonic hedgehog is required for long-range signaling activity and effective modulation of signaling by Ptc1. Cell 105: 599–612.
• Niswander, L., S. Jeffrey, G. R. Martin and C. Tickle. 1994. A positive feedback loop coordinates growth and patterning in the vertebrate limb. Nature 371: 609–612.
• Offield, M. F. and 7 others. 1996. PDX-1 is required for pancreatic outgrowth and differentiation of the rostral duodenum. Development 122: 983–995.
• Placzek, M., M. Tessier-Lavigne, T. Yamada, T. Jessell and J. Dodd. 1990. Mesodermal control of neural cell identity: Floor plate induction by the notochord. Science 250: 985–988.
• Porter, A. G. and S. Dhakshinamoorty. 2004. Apoptosis initiated by dependence reseptors: a new paradigm for cell death? BioEssays 26; 656–664.
• Riddle, R. D., R. L. Johnson, E. Laufer and C. Tabin. 1993. Sonic hedgehog mediates the polarizing activity of the ZPA. Cell 75: 1401–1416.
• Roberts, D. J., R. L. Johnson, A. C. Burke, C. E. Nelson, B. A. Morgan and C. Tabin. 1995. Sonic hedgehog is an endodermal signal inducing Bmp-4 and Hox genes during induction and regionalization of the chick hindgut. Development 121: 3163–3174.
• Roelink, H., J. A. Porter, C. Chiang, Y. Tanabe, D. T. Chang, P. A. Beachy and T. M. Jessell. 1995. Floor plate and motor neuron induction by different concentrations of amino-terminal cleavage product of Sonic hedgehog autoproteolysis. Cell 81: 445–455.
• Roessler, E. and 7 others. 1996. Mutations in the human sonic hedgehog gene cause holoprosencephaly. Nature Genet. 14: 357–360.
• Salazar-Ciudad, I. and J. Jernvall. 2002. A gene network model accounting for development and evolution of mammalian teeth. Proc. Natl. Acad. Sci. USA 99: 8116–8120.
• Salazar-Ciudad, I. and J. Jernvall. 2004. How different types of pattern formation mechanisms affect the evolution of form and development. Evol. Dev. 6: 6–16.
• Scherz, P. J., B. D. Harfe, A. P. McMahon and C. J. Tabin. 2004. The limb Shh-Fgf feedback loop is terminated by the expansion of former ZPA cells. Science 305: 396.
• Stolow, M. A. and Y. B. Shi. 1995. Xenopus sonic hedgehog as a potential morphogen during embryogenesis and thyroid hormone dependent metamorphosis. Nucleic Acids Res. 23: 2555–2562.
• Thibert, C., M.-A. Teillet, F. Lapointe, L. Mazelin, N. M. Le Douarin, and P. Mehlen. 2003. Inhibition of neuroepithelial Patched-induced apoptosis by sonic hedgehog. Science 301: 843–846.
• Torok, M. A., D. M. Gardiner, J.-C. Izpisúa-Belmonte and S. V. Bryant. 1999. Sonic hedgehog (Shh) expression in developing and regenerating axolotl limbs. J. Exp. Zool. 284: 197–206.
• Vaahtokari, A., T. Aberg, J. Jernvall, S. Keränen and I. Thesleff. 1996a. The enamel knot as a signalling center in the developing mouse tooth. Mech. Dev. 54: 39–43.
• Yamamoto, Y., D. Stock and W. R. Jeffery. 2004. Hedgehog signaling controls eye degeneration in blind cavefish. Nature 431: 844–8477.
• Yu, M., P. Wu, R. B. Widelitz and C. M. Chuong. 2002. The morphogenesis of feathers. Nature 420: 308– 312.
• Zúñiga, A., A. P. Haramis, A. P. McMahon and R. Zeller. 1999. Signal relay by BMP antagonism controls the Shh/Fgf4 feedback loop in vertebrate limb buds. Nature 401: 598–602.
Categorías:- Proteínas
- Biología del desarrollo
- Genética
Wikimedia foundation. 2010.