- Intrón
-
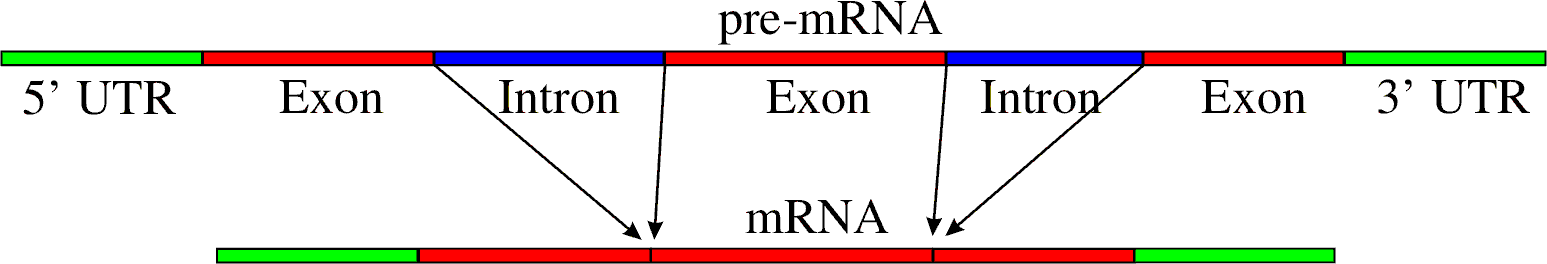
Un intrón es una región del ADN que debe ser eliminada del transcrito primario de ARN, a diferencia de los exones que son regiones que codifican para una determinada proteína. Los intrones son comunes en todos los tipos de ARN eucariota, especialmente en los ARN mensajeros (ARNm), además pueden encontrarse en algunos ARNt y ARNr de procariotas.
El número y longitud de los intrones varía enormemente entre especies, así como entre los genes de una misma especie. Por ejemplo, el pez globo, Takifugu rubripees, tiene pocos intrones en su genoma; mientras que los mamíferos y las angiospermas (plantas con flores) suelen presentar numerosos intrones.
Contenido
Introducción
Los intrones fueron descubiertos por Phillip Allen Sharp y Richard J. Roberts, lo que les supuso ganar el Premio Nobel de Fisiología o Medicina en 1993. El término intrón fue introducido por el bioquímico estadounidense Walter Gilbert en 1978.
Los intrones pueden representar un sitio alternativo de splicing, pudiendo dar diferentes tipos de proteínas. El control del splicing esta regulado por una amplia variedad de señales moleculares. Los intrones también pueden contener “información antigua”, es decir, fragmentos de genes que probablemente se expresaban pero que actualmente no se expresan.
 Ilustración del proceso de ajuste desde pre-ARN a ARN.
Ilustración del proceso de ajuste desde pre-ARN a ARN.
Tradicionalmente se ha afirmado que los intrones son fragmentos de ADN carentes de información. Sin embargo esta afirmación es cuestionada y actualmente goza de pocos adeptos. Se sabe que los intrones contienen varias secuencias pequeñas que son importantes para un ayuste eficiente.
Algunos intrones del grupo I y II son ribozimas capaces de catalizar su propio splicing fuera del ARN. El descubrimiento de estas propiedades auto-catalíticas supuso el Premio Nobel de Química a Thomas R. Cech y Sidney Altman en 1989.
Clasificación de los intrones
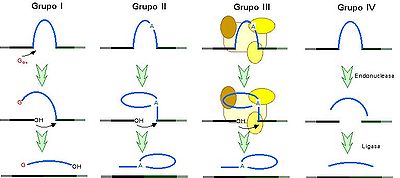 Ilustración que recoge la clasificación de los intrones de acuerdo al método de ayuste, basado en una reacción de transesterificación en los tres primeros casos y en un corte endonucleótido en el cuarto caso. Imagen extraída de Saladrigas V, Claros G (2002): Vocabulario inglés-español de bioquímica y biología molecular (1.ª entrega) Panace@ III (9-10): 13-28. Vocabulario completo en BioROM. Licenciada por Panace@.
Ilustración que recoge la clasificación de los intrones de acuerdo al método de ayuste, basado en una reacción de transesterificación en los tres primeros casos y en un corte endonucleótido en el cuarto caso. Imagen extraída de Saladrigas V, Claros G (2002): Vocabulario inglés-español de bioquímica y biología molecular (1.ª entrega) Panace@ III (9-10): 13-28. Vocabulario completo en BioROM. Licenciada por Panace@.Actualmente se reconocen cuatro clases de intrones:
- Intrones del grupo I
- Intrones del grupo II
- Intrones del grupo III
- Intrones nucleares, spliceosomales o intrones del grupo IV
Los intrones del grupo I, II y III son intrones que sufren de autosplicing mediante reacciones de transesterificación. La frecuencia con la que encontramos estos intrones en el genoma es relativamente rara si la comparamos con la frecuencia de los intrones spliceosomales.
Los intrones del grupo II y III son muy similares y presentan una estructura secundaria altamente conservada. De hecho a veces los intrones del grupo III son identificados como intrones del grupo II debido a su similitud funcional y estructural.
Los intrones del grupo I están presentes en los genes de ARNr de algunos eucariotas inferiores y en los genes mitocondriales de hongos. Se caracterizan por eliminarse mediante un proceso autocatalítico que requiere de una guanosina o un nucleótido de guanosina libre; así como por carecer de secuencias consenso en los puntos de empalme, aunque pueden tenerlas en su interior.
Los del grupo II y III se eliminan mediante un proceso autocatalítico que requiere de una adenina o de un spliceosoma, respectivamente. En ambos grupos, durante el proceso de empalme de los exones, se forma una estructura en lazo característica denominada lariat.
Los del grupo IV están presentes en los ARNt de los eucariotas y se caracterizan por ser los únicos que se eliminan mediante un corte endonucleótido seguido de un ligamiento en lugar de la reacción de transesterificación
Evolución de los intrones
Existen dos modelos, contrapuestos, que explican el origen y la evolución de los intrones nucleares o ayustosomales. Estos modelos se conocen como intrones tempranos (IE) o intrones tardíos (IL).
El modelo IE propone que los intrones eran extremadamente numerosos en los ancestros de procariotas y eucariotas; y se fueron perdiendo a lo largo de la evolución. Este modelo se basa en la hipótesis de que los intrones fueron mediadores que facilitaron la combinación de exones, facilitando por tanto la evolución de nuevos genes.
El modelo IL propone que los intrones aparecieron tras la divergencia de procariotas y eucariotas. Este modelo se basa en la observación de que los intrones ayustosomales únicamente se han encontrado en eucariotas
Véase también
-
- Splicing
- Splicing alternativo
- Splicing de ARN
- ADN no codificante
- Exón
Referencias
- Saladrigas V, Claros G (2002): Vocabulario inglés-español de bioquímica y biología molecular (1.ª entrega) Panace@ III (9-10): 13-28. Vocabulario completo en BioROM.
- Gilbert, Walter (1978): Why genes in pieces. Nature 271(5645): 501. doi 10.1038/271501a0
- Roy, Scott William & Gilbert, Walter (2006): The evolution of spliceosomal introns: patterns, puzzles and progress. Nature Reviews Genetics 7: 211-221. doi 10.1038/nrg1807 PDF fulltext
- Gogarten, J. Peter & Hilario, Elena (2006): Inteins, introns, and homing endonucleases: recent revelations about the life cycle of parasitic genetic elements. BMC Evolutionary Biology 6: 94 doi 10.1186/1471-2148-6-94 PDF fulltext
- Yandell, Mark; Mungall, Chris J.; Smith, Chris; Prochnik, Simon; Kaminker, Joshua; Hartzell, George; Lewis, Suzanna & Rubin, Gerald M. (2006): Large-Scale Trends in the Evolution of Gene Structures within 11 Animal Genomes. PLoS Comput. Biol. 2(3): 113-125. doi 10.1371/journal.pcbi.0020015 PDF fulltext Supporting Information
Enlaces
Categorías:- ADN
- Genes

Wikimedia foundation. 2010.