- Célula eucariota
-
Célula eucariota
Se denomina eucariotas a todas las células que tienen su material hereditario fundamental (su información genética) encerrado dentro de una doble membrana, la envoltura nuclear, que delimita un núcleo celular. Igualmente estas células vienen a ser microscópicas pero de tamaño grande y variado comparado con las otras células.
La alternativa a la organización eucariótica de la célula la ofrece la llamada célula procariota. En estas células el material hereditario se encuentra dentro de diferentes compartimientos llamados orgánulos, en el seno del citoplasma. Las células eucariotas no cuentan con un compartimiento alrededor de la membrana plasmática (periplasma), como el que tienen las células procariotas.
A los organismos formados por células eucariotas se les denomina eucariontes.
El paso de procariotas a eucariotas significó el gran salto en complejidad de la vida y uno de los más importantes de su evolución.[1] Sin este paso, sin la complejidad que adquirieron las células eucariotas no habrían sido posibles ulteriores pasos como la aparición de los pluricelulares. La vida, probablemente, se habría limitado a constituirse en un conglomerado de bacterias. De hecho, los cuatro reinos restantes procedemos de ese salto cualitativo. El éxito de estas células eucariotas posibilitó las posteriores radiaciones adaptativas de la vida que han desembocado en la gran variedad de especies que existe en la actualidad.
Contenido
Organización
Las células eucariotas presentan un citoplasma muy compartimentado, con orgánulos (membranosos) separados o interconectados, limitados por membranas biológicas que son de la misma naturaleza esencial que la membrana plasmática. El núcleo es solamente el más notable y característico de los compartimentos en que se divide el protoplasma, es decir, la parte activa de la célula. En el protoplasma distinguimos tres componentes principales, a saber, la membrana plasmática, el núcleo y el citoplasma, constituido por todo lo demás. Las células eucariotas están dotadas en su citoplasma de un citoesqueleto complejo, muy estructurado y dinámico, formado por microtúbulos y diversos filamentos proteicos. Además puede haber pared celular, que es lo típico de plantas, hongos y protistas pluricelulares, o algún otro tipo de recubrimiento externo al protoplasma.
Fisiología
Las células eucariotas contienen en principio mitocondrias, orgánulos que habrían adquirido por endosimbiosis de ciertas bacterias primitivas, lo que les dota de la capacidad de desarrollar un metabolismo aerobio. Sin embargo, en algunos eucariotas del reino protistas las mitocondrias han desaparecido secundariamente en el curso de la evolución, en general derivando a otros orgánulos, como los hidrogenosomas.
Algunos eucariontes realizan la fotosíntesis, gracias a la presencia en su citoplasma de orgánulos llamados plastos, los cuales derivan por endosimbiosis de bacterias del grupo denominado cianobacterias (algas azules).
Aunque demuestran una diversidad increíble en su forma, comparten las características fundamentales de su organización celular, arriba resumidas, y una gran homogeneidad en lo relativo a su bioquímica (composición), y metabolismo, que contrasta con la inmensa heterogeneidad que en este terreno presentan los procariontes (bacteria en sentido amplio).
Origen de los eucariotas
El origen de los eucariotas se encuentra en sucesivos procesos simbiogenéticos (procesos simbióticos extremos que desembocan en la transferencia de material genético) entre diferentes bacterias.
Hoy en día existen pruebas concluyentes a favor de la teoría de que la célula eucariota moderna evolucionó en etapas mediante la incorporación estable de las bacterias. Diferentes aportaciones justifican el origen de los croroplastos y las mitocondrias a partir de éstas.
Isabel Esteve, Discurso de presentación de Lynn Margulis en el acto de investidura doctora honoris causa UAB[2]A principios del siglo XX, en 1909, el ruso Kostantin S. Mereschovky presentó la hipótesis según la cual el origen de los cloroplastos tendría su origen en procesos simbióticos.[3] A parecidas conclusiones llegaron Kozo-Polyansky y Andrey Faminstyn (también de la escuela rusa) que consideraban la simbiogénesis “crucial para la generación de novedad biológica".[4] En Francia, el biólogo Paul Portier, en 1918, y Ivan Wallin en Estados Unidos en 1927, llegaron a las mismas conclusiones. Trabajos que o bien pasaron inadvertidos (como los de la escuela rusa) o no fueron tenidos en cuenta (en el caso de Portier y Wallis) costando el prestigio profesional a sus proponentes.
Lynn Margulis rescata estos trabajos y en 1967 en el artículo On origen of mitosing cells presenta la que llegaría a conocerse como Serial Endosymbiosis Theory (SET) (Teoría de la endosimbiosis seriada) en la que describe con concreción, mediante procesos simbiogenéticos, los pasos seguidos por las procariotas hasta la eclosión de las diferentes células eucariotas. Los tres pasos descritos por Margulis son:
Primera incorporación simbiogenética:
- Una bacteria consumidora de azufre, que utilizaba el azufre y el calor como fuente de energía (arquea fermentadora o termoacidófila), se habría fusionado con una bacteria nadadora (espiroqueta) habiendo pasado a formar un nuevo organismo y sumaría sus características iniciales de forma sinérgica (en la que el resultado de la incorporación de dos o más unidades adquiere mayor valor que la suma de sus componentes). El resultado sería el primer eucarionte (unicelular eucariota) y ancestro único de todos los pluricelulares. El núcleoplasma de la células de animales, plantas y hongos sería el resultado de la unión de estas dos bacterias.
- A las características iniciales de ambas células se le sumaría una nueva morfología más compleja con una nueva y llamativa resistencia al intercambio genético horizontal. El ADN quedaría confinado en un núcleo interno separado del resto de la célula por una membrana.[5]
Segunda incorporación simbiogenética:
- Este nuevo organismo todavía era anaeróbico, incapaz de metabolizar el oxígeno, ya que este gas suponía un veneno para él, por lo que viviría en medios donde este oxigeno, cada vez más presente, fuese escaso. En este punto, una nueva incorporación dotaría a este primigenio eucarionte de la capacidad para metabolizar oxigeno. Este nuevo endosombionte, originariamente bacteria respiradora de oxigeno de vida libre, se convertiría en las actuales mitocondrias y peroxisomas presentes en las células eucariotas de los pluricelulares, posibilitando su éxito en un medio rico en oxígeno como ha llegado a convertirse el planeta Tierra. Los animales y hongos somos el resultado de esta segunda incorporación.[6]
Tercera incorporación simbiogenética:
- Esta tercera incorporación originó el Reino vegetal, las recientemente adquiridas células respiradoras de oxígeno fagocitarían bacterias fotosintéticas y algunas de ellas, haciéndose resistentes, pasarían a formar parte del organismo, originando a su vez un nuevo organismo capaz de sintetizar la energía procedente del Sol. Estos nuevos pluricelulares, las plantas, con su éxito, contribuyeron y contribuyen al éxito de animales y hongos.[7]
El primer paso, al día de hoy, no se considera demostrado. A finales de los años ochenta y principio de los noventa diversos trabajos no admitían las homologías propuestas entre los flagelos de los eucariontes y de las espiroquetas.[8] [9] [10] [11] Margulis defiende que las asociaciones entre espiroquetas y protistas apoyan su teoría, y "la comparación de genes y genomas arqueobaterianos con secuencias de eucariontes han demostrado la relación filogenética de ambos grupos".[12] No obstante, desde su formulación por Margulis, han surgido innumerables interrogantes. Margulis admite que este es el punto de su teoría con más dificultades para defenderse y Antonio Lazcano, en 2002, previene que para comprender el origen de este primer paso, se acepte o no su origen simbiogenético, "es indispensable secuenciar no sólo los genomas de una gama representativa de protistas sino también reconocer la importancia del estudio de la biología de estos organismos".[12]
Ya en los años setenta surgió, como alternativa al origen simbiogenético de este primer paso, la hipótesis de que éste se hubiese producido mediante invaginaciones,[13] propuesta que no contradice el paradigma neodarviniano y que, aún hoy, se considera plausible por amplios sectores del mundo académico.
Recurrentemente se han propuesto diferentes hipótesis, también simbiogéneticas, en las que el propio núcleo sería resultado de la incorporación de otro simbionte, como en el caso de las mitocondrias y los cloroplastos.[14]
A Margulis le ha costado más de 30 años hacer valer su teoría hasta lograr demostrar la incorporación de tres de los cuatro simbiontes, o si se quiere, dos de los tres pasos propuestos (la incorporación de las espiroquetas no se considera probada).
El mundo académico se vio forzado a aceptar la parte de la teoría de Margulis que hoy se enseña en todos los libros de texto: que las mitocondrias y los cloroplastos provienen, por simbiosis, de antiguas bacterias de vida libre. La idea convencional, sin embargo, persiste aún gracias a que la teoría de Margulis se suele presentar en una versión edulcorada que no capta el fondo de la cuestión.
Javier Sampedro, Deconstruyendo a Darwin, p. 40Afortunadamente, gracias a la genial bióloga estadounidense Lynn Margulis, hoy tenemos la solución a este desconcertante enigma: una explicación científica mucho más sensata, lúcida y creativa que la que se ha empeñado en sostener la ortodoxia neodarwinista durante los últimos 35 años, pese a tener la solución, publicada por Margulis en 1967, literalmente delante de sus narices. La ortodoxia se ha resistido con uñas y dientes —en gran medida sigue resistiéndose— a aceptar la teoría de Margulis por el sencillo hecho de que no encaja con sus prejuicios darwinistas. Pero si usted logra liberarse de ese lastre irracional y anticientífico, verá inmediatamente que la idea de Margulis no sólo es la correcta, sino que está dotada de un luminoso poder explicativo. El modelo de Margulis sobre el origen de la célula eucariota no es gradual, pero no le hace ninguna falta para ser factible. Implica un suceso brusco y altamente creativo, pero también enteramente materialista, ciego y mecánico.
Javier Sampedro, Deconstruyendo a Darwin.[15]Margulis opina que el primer paso, la incorporación de la espiroqueta, es el que más dificultades encuentra para su demostración, no obstante, al día de hoy son numerosas las voces desde la biología que defienden el origen simbiotico también para este primer paso.
Organismos eucariontes
Los organismos eucariontes forman el dominio Eukarya que incluye a los organismos más conocidos, repartidos en cuatro reinos: Animalia (animales), Plantae (plantas), Fungi y Protista Incluyen a la gran mayoría de los organismos extintos morfológicamente reconocibles que estudian los paleontólogos. Los ejemplos de la disparidad eucariótica van desde un dinoflagelado (un protista unicelular fotosintetizador), un árbol como la sequoia, un calamar, o un racimo de setas (órganos reproductivos de hongos), cada uno con células distintas y, en el caso de los pluricelulares, a menudo muy variadas.
Diferencias entre células eucariotas
Existen diversos tipos de células eucariotas entre las que destacan las células de animales y plantas. Los hongos y muchos protistas tienen, sin embargo, algunas diferencias substanciales.
Células animales
-
El artículo principal de esta categoría es Célula animal.
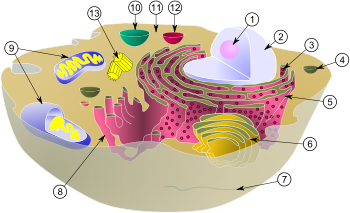 Estructura de una célula animal típica: 1. Nucléolo, 2. Núcleo, 3. Ribosoma, 4. Vesícula, 5. Retículo endoplasmático rugoso, 6. Aparato de Golgi, 7. Citoesqueleto (microtúbulos), 8. Retículo endoplasmático liso, 9. Mitocondria, 10. Peroxisoma, 11. Citoplasma, 12. Lisosoma. 13. Centriolo.
Estructura de una célula animal típica: 1. Nucléolo, 2. Núcleo, 3. Ribosoma, 4. Vesícula, 5. Retículo endoplasmático rugoso, 6. Aparato de Golgi, 7. Citoesqueleto (microtúbulos), 8. Retículo endoplasmático liso, 9. Mitocondria, 10. Peroxisoma, 11. Citoplasma, 12. Lisosoma. 13. Centriolo.
Las células animales componen los tejidos de los animales y se distinguen de las células vegetales en que carecen de paredes celulares y de cloroplastos y poseen centríolos y vacuolas más pequeñas y, generalmente, más abundantes. Debido a la carencia de pared celular rígida, las células animales pueden adoptar variedad de formas e incluso pueden fagocitar otras estructuras.
Células vegetales
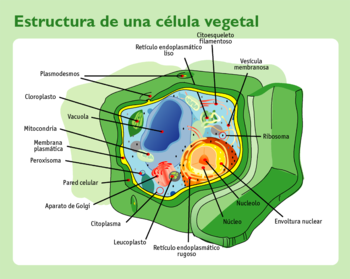 Estructura de una célula vegetal típica: 1. Núcleo, 2. Nucléolo, 3. Membrana nuclear, 4. Retículo endoplasmático rugoso, 5. Leucoplasto, 6. Citoplasma, 7. Dictiosoma / Aparato de Golgi, 8. Pared celular, 9. Peroxisoma, 10. Membrana plasmática, 11. Mitocondria, 12. Vacuola central, 13. Cloroplasto, 14. Plasmodesmos, 15. Retículo endoplasmático liso, 16. Citoesqueleto, 17. Vesícula, 18. Ribosomas.
Estructura de una célula vegetal típica: 1. Núcleo, 2. Nucléolo, 3. Membrana nuclear, 4. Retículo endoplasmático rugoso, 5. Leucoplasto, 6. Citoplasma, 7. Dictiosoma / Aparato de Golgi, 8. Pared celular, 9. Peroxisoma, 10. Membrana plasmática, 11. Mitocondria, 12. Vacuola central, 13. Cloroplasto, 14. Plasmodesmos, 15. Retículo endoplasmático liso, 16. Citoesqueleto, 17. Vesícula, 18. Ribosomas.Las características distintivas de las células de las plantas son:
- Una vacuola central grande (delimitada por una membrana, el tonoplasto), que mantiene la forma de la célula y controla el movimiento de moléculas entre citosol y savia.
- Una pared celular compuesta de celulosa y proteínas, y en muchos casos, lignina, que es depositada por el protoplasto en el exterior de la membrana celular. Esto contrasta con las paredes celulares de los hongos, que están hechas de quitina, y la de los procariontes, que están hechas de peptidoglicano.
- Los plasmodesmos, poros de enlace en la pared celular que permiten que las células de la plantas se comuniquen con las células adyacentes. Esto es diferente a la red de hifas usada por los hongos.
- Los plastos, especialmente cloroplastos que contienen clorofila, el pigmento que da a la plantas su color verde y que permite que realicen la fotosíntesis.
- Los grupos de plantas sin flagelos (incluidas coníferas y plantas con flor) también carecen de los centriolos que están presentes en las células animales. Estos también se pueden encontrar en los animales de todos los tipos es decir en un mamifero en una ave o en un reptil
Células de los hongos
Las células de los hongos, en su mayor parte, son similares a las células animales, con las excepciones siguientes:
- Una pared celular hecha de quitina.
- Menor definición entre células. Las células de los hongos superiores tienen separaciones porosas llamados septos que permiten el paso de citoplasma, orgánulos, y a veces, núcleos. Los hongos primitivos no tienen tales divisiones, y cada organismo es esencialmente una supercélula gigante. Estos hongos se conocen como coenocíticos.
- Solamente los hongos más primitivos, Chytridiomycota, tienen flagelos.
Comparación de estructuras en células animales y vegetales Célula animal típica Célula vegetal típica Estructuras básicas Orgánulos Estructuras adicionales - Flagelo (sólo en gametos)
- Pared celular
- Plasmodesmos
Véase también
- Eukarya
- Orgánulo
- Endosimbiosis
- Célula
- Célula procariota
- Célula vegetal
- organismos unicelulares
Referencias
- Célula Eucariota: orgánulos, Universidad Nacional del Nordeste
Notas
- ↑
Su origen puede ser considerado como el avance más significativo de la evolución orgánica.
Stebbins, 1980, p. 380 - ↑ Acto de investidura doctora honoris causa por la Universidad Autònoma de Barcelona UAB. 1/02/2006
- ↑ Margulis, 2002, p. 35.
- ↑ La bibliografía rusa, interpretada por el historiador de la ciencia Liya N. Khakhina, no estuvo disponible en inglés hasta el año 1922. Fueron necesarias dos generaciones de académicos para resumir la gran bibliografía de los botánicos rusos. Parece hoy como si esta bibliografta fuera ignorada por esta misma razón. La literatura antigua escrita por botánicos rusos carece de atractivo para el mercado anglófono.|Margulis, Sagan (2003), p. 142.}}
- ↑
En primer lugar, un tipo de bacteria amante del azufre y del calor, llamada arqueobacteria fermentadora (o termoacidófila), se fusionó con una bacteria nadadora. Juntos, los dos componentes integrados de la fusión se convirtieron en el nucleocitoplasma, la sustancia base de los ancestros de las células animales, vegetales y fúngicas. Este temprano protista nadador era, como sus descendientes actuales, un organismo anaerobio. Envenenado por el oxígeno, vivía en arenas y lodos donde abundaba la materia orgánica, en grietas de las rocas, en charcos y estanques donde este elemento estaba ausente o era escaso.
Lynn Margulis, Una revolución en la Evolución, Cap.: Individualidad por incorporación.En las plantas y en los animales el núcleo delimitado por una membrana desaparece cada vez que la célula se multiplica por división; la membrana se disuelve y los cromosomas se hacen visibles. La cromatina, el material que se tiñe de rojo y del que están hechos los cromosomas, se enrolla formando estructuras fácilmente observables. La explicación de libro de texto de este proceso es que la cromatina se condensa formando cromosomas visibles y contables cuyo número es representativo de la especie en cuestión. La danza de los cromosomas, que más tarde desaparecen pasando a ser cromatina débilmente enrollada a medida que reaparece la membrana nuclear, constituye los pasos del proceso de división celular de la mitosis. La mitosis, con sus numerosas variaciones en las células nucleadas de los protoctistas y los hongos, evolucionó en los más tempranos organismos con núcleo.
Margulis, Una revolución en la Evolución, p. 98. - ↑
Después de que evolucionara la mitosis en los protistas nadadores, otro tipo de microorganismo de vida libre fue incorporado a la fusión: una bacteria que respiraba oxígeno. Surgieron células todavía más grandes, más complejas. El triplemente complejo respirador de oxígeno (amante del calor y del ácido, nadador y respirador de oxígeno) se volvió capaz de engullir alimento en forma de partículas. Estas células con núcleo, seres complejos y asombrosos que nadaban y respiraban oxígeno, aparecieron por primera vez sobre la Tierra quizá tan pronto como hace unos 2.000 millones de años. Esta segunda fusión, en la que el anaerobio nadador adquirió un respirador de oxígeno, condujo a células con tres componentes cada vez más preparadas para soportar los niveles de oxígeno libre que se acumulaban en el aire. Juntos, el delicado nadador, la arqueobacteria tolerante al calor y al ácido y el respirador de oxígeno, formaban ahora un único y prolífico individuo que produjo nubes de prole.
Margulis, Una revolución en la Evolución, Cap.: Individualidad por incorporación. - ↑
En la adquisición final de la serie generadora de células complejas, los respiradores de oxígeno engulleron, ingirieron, pero no pudieron digerir bacterias fotosintéticas de color verde brillante. La «incorporación» literal tuvo lugar tras una gran lucha en la que las bacterias verdes no digeridas sobrevivieron y la fusión completa prevaleció. Con el tiempo las bacterias verdes se convirtieron en cloroplastos (paso 4, figura 1.1). Como cuarto miembro, estos productivos amantes del sol se integraron con los demás socios anteriormente independientes. Esta fusión final dio lugar a las algas verdes nadadoras. Estas antiguas algas verdes nadadoras no sólo son los ancestros de las células vegetales actuales; todos sus componentes individuales todavía están vivos y en buena forma, nadando, fermentando y respirando oxígeno.
Margulis, Una revolución en la Evolución, Cap.: Individualidad por incorporación. - ↑ Bermudes, D., Fracek, S. P. Jr., Laursen, R. A., Margulis, L., Obar, R. and Tzertzinis, G. (1987). "Tubulinlike protein from Spirochaeta bajacaliforniensis", in Annals of the New York Academy of Sciences: Endocytobiology III. New York, The New York Academy of Sciences. 503: 515-527.
- ↑ Barth, A. L., Stricker, J. A. and Margulis, L. (1991). "Search for Eukaryotic Motility Proteins in Spirochetes - Immunological Detection of a Tektin-Like Protein in Spirochaeta-Halophila". BioSystems, V24(N4): 313-319.
- ↑ Bermudes, D., Hinkle, G. and Margulis, L. (1994). "Do Prokaryotes Contain Microtubules". Microbiological Reviews, V58(N3): 387-400.
- ↑ Munson, D., Obar, R., Tzertzinis, G. and Margulis, L. (1993). "The Tubulin-Like S1 Protein of Spirochaeta Is a Member of the Hsp65 Stress Protein Family". BioSystems, V31(N2-3): 161-167.
- ↑ a b Lazcano, 2002, p. 197.
- ↑
Las membranas internas de las células eucariotas, sobre todo la membrana nuclear y el retículo endoplasmático, se explican mejor como invaginaciones. Como el cromosoma de los procariotas se halla unido a la membrana celular al menos durante su división (Stanier, Doudoroff & Adelberg, 1970), una invaginación de esta parte de la membrana celular situaría automáticamente a los cromosomas en el interior del núcleo. Las conexiones y la semejanza molecular entre los flagelos y centríolos podrían explicarse suponiendo que en los primeros eucariotas, la membrana nuclear continuaría unida a la membrana celular externa durante la evolución simultánea del aparato mitótico y de los flagelos. Podría explicarse la existencia de DNA en el cuerpo basal del flagelo por adición de un plásmido, tal como han sugerido Raff & Mahler.
Stebbins, 1980. - ↑
La idea de que el núcleo mismo sea el resultado de una simbiosis es una hipótesis recurrente, que se propuso, en forma vaga y poco precisa, en 1909 por el mismo Merezhkovsky (Margulis 1993). Aunque la continuidad funcional y anatómica entre el medio intranuclear y el citoplasma eucarionte está perfectamente demostrada, de cuando en cuando se revive en la bibliografía la posibilidad de que el núcleo provenga, al igual que las mitocondrias y los cloroplastos, de un simbionte microbiano (Pickett-Heaps 1974, Hartman 1975, 1984, Hartman y Fedorov 2002, Sogin 1991, Lake y Rivera 1994, Gupta y Golding 1996, Vellai et al. 1998, Horiike et al. 2001). Lazcano, 2002, p. 191.
- ↑ P. 35.
Bibliografía
- Margulis, Lynn (2002). Planeta Simbiótico. Un nuevo punto de vista sobre la evolución., Victoria Laporta Gonzalo (trad.), Madrid: Editorial Debate.
- Margulis, Lynn; Dorion Sagan (2003). Captando Genomas. Una teoría sobre el origen de las especies., Ernst Mayr (prólogo). David Sempau (trad.), 1ª edición, Barcelona: Editorial Kairós. ISBN 84-7245-551-3.
- Margulis, Lynn, (2003) Una Revolución en la Evolución (escritos seleccionados) Colección Honoris Causa, Universitat de Valencia.
- Sampedro, Javier (2002). Deconstruyendo a Darwin, Ginés Morata (prólogo)., 1ª edición, Barcelona: Editorial Crítica.
- Michel E. N. Majerus, 1991, Simbiontes hereditarios causantes de efectos deletéreos en los antropos.
Categoría: Célula

Wikimedia foundation. 2010.