- Memoria declarativa
-
La memoria declarativa (en ocasiones, y de una forma más general, conocida como memoria explícita) es uno de los dos tipos de memoria a largo plazo en los seres humanos. Hace referencia a todos aquellos recuerdos que pueden ser evocados de forma consciente, como hechos o eventos específicos.[1] Su contraparte se conoce como memoria no declarativa o memoria procedimental, y engloba todos los recuerdos inconscientes, como determinadas habilidades o destrezas (por ejemplo, la habilidad necesaria para montar en bicicleta). La memoria declarativa puede dividirse en dos categorías: memoria episódica, que almacena las experiencias personales específicas; y la memoria semántica, que almacena información sobre hechos.[2]
Contenido
Tipos
Existen dos tipos de memoria declarativa. La memoria semántica almacena todos aquellos conocimientos sobre hechos generales que son independientes de la experiencia personal. Algunos ejemplos serían los diversos tipos de comidas, las capitales de los países o los conocimientos léxicos (vocabulario).[2] La memoria episódica almacena los recuerdos referidos a eventos específicos de la vida personal, como asistir a una determinada reunión, o un viaje realizado durante las vacaciones. La evocación de estos recuerdos puede entenderse como una forma de revivir mentalmente eventos pasados. Generalmente se considera a la memoria episódica como el sistema que sustenta a la memoria semántica.[2]
Historia
El estudio de la memoria humana se remonta a hace más de 2000 años. Uno de las primeras tentativas de comprender el funcionamiento de la memoria puede encontrrse en el principal tratado de Aristóteles, Acerca del alma, en el que compara a la mente humana con una tabula rasa.[3] En esta obra, Aristóteles teoriza acerca del hecho de que el ser humano nace libre de cualquier tipo de conocimiento, y resulta ser la suma de todas sus experiencias vitales. No obstante, no sería hasta finales del siglo XIX cuando un joven filósofo alemán llamado Hermann Ebbinghaus desarrollara la primera aproximación científica al estudio de la memoria.[4] Algunos de sus descubrimientos, como la curva de aprendizaje han permanecido vigentes hasta el día de hoy. Su máxima aportación al campo de la investigación de la memoria fue el hecho de demostrar que la memoria podía ser objeto de estudio científico.
En el año 1972, Endel Tulving propuso una distinción entre la memoria episódica y la memoria semántica.[2] Este hecho fue rápidamente adoptado por la comunidad científica, y en la actualidad se encuentra ampliamente aceptado. A partir de aquí, Daniel Schacter propuso en el año 1985 una diferenciación más general entre memoria explícita (declarativa) e implícita (procedimental)[5]
Gracias a los recientes avances en las tecnologías de neuroimagen, se ha desarrollado una gran cantidad de estudios que relacional a la memoria declarativa con determinadas áreas cerebrales específicas. A pesar de estos avances en el terreno de la psicología cognitiva, aún queda mucho por descubrirse en lo que respecta a los mecanismos operativos de la memoria declarativa.[6] Aún no está claro si la memoria declarativa está mediatizada por algún tipo particular de "sistema de memoria", o si podría clasificarse con más exactitud como un tipo de conocimiento.[6]
Neuropsicología
Función cerebral normal
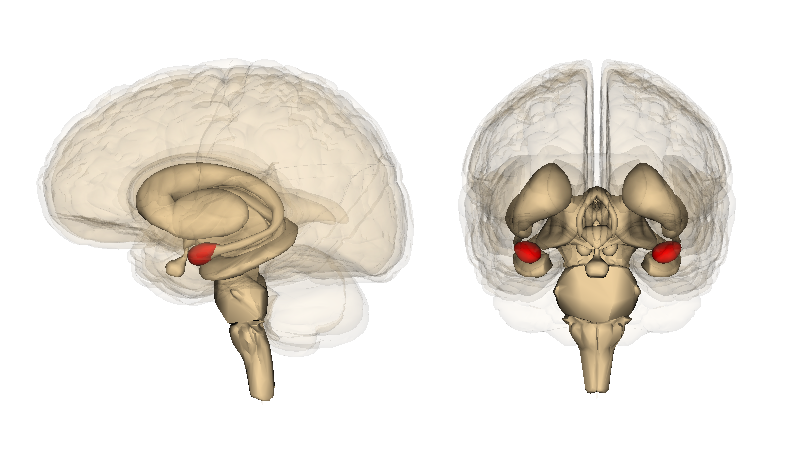
Hipocampo
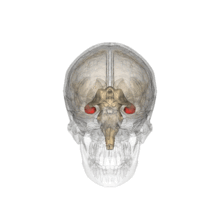 El hipocampo, destacado en rojo.
El hipocampo, destacado en rojo.
Aunque muchos psicólogos opinan que todo el cerebro se encuentra implicado en la función memorística, el hipocampo y las estructuras adyacentes parecen desempeñar el papel más importante, especialmente en lo que a la memoria declarativa se refiere.[7] La capacidad para retener y evocar recuerdos episódicos depende en gran medida del hipocampo,[7] mientras que en la formación de nuevos recuerdos declarativos intervienen tanto el hipocampo como el parahipocampo.[8] Otros estudios han hallado una relación entre la corteza parahipocampal y la memoria de reconocimiento superior.[8]
Eichenbaum et. al (2001) desarrollaron un modelo de tres fases, que propone que el hipocampo interviene en tres ocasiones en relación con la memoria episódica:
- Media en la grabación de recuerdos episódicos.
- Identifica características comunes entre episodios distintos.
- Enlaza estos episodios comunes en un espacio de memoria.
Para sustentar este modelo, se utilizó una versión de las tareas de inferencia transitiva de Jean Piaget para mostrar que el hipocampo, de hecho, cumple la función de espacio de memoria.[7] El hipocampo no sirve de almacén de recuerdos, pero cumple la función de establecer asociaciones o enlaces entre los recuerdos de forma que puedan estar disponibles para acceder a ellos de forma consciente; es decir, una especie de "índice" de recuerdos.
Cuando se experimenta un evento por primera vez, se crea un enlace en el hipocampo que nos permitirá recordar ese evento en el futuro. Además, se van creando otros enlaces para las características relacionadas con ese evento. Por ejemplo, al conocer a una persona, se crea un enlace para ella, además de otros enlaces conectados por los que se puede recordar el color de su camisa, el tiempo que hacía en el momento de conocerla, etc. Los eventos concretos y específicos son más fáciles de recordar y evocar cuantas más veces nos expongamos a ellos, lo que aumenta la fuerza de la asociación en el espacio de los recuerdos, permitiendo una recuperación más rápida de la información.[7]
Las distintas células hipocampales (neuronas) se activan en función de la información a la que nos exponemos en cada momento. Algunas células responden específicamente a la información espacial, a ciertos estímulos (olores, etc.) o ante determinados comportamientos, como se ha visto en la Tarea del Laberinto Radial.[7] Por lo tanto, el hipocampo nos permite reconocer ciertas situaciones o ambientes como diferentes o similares a otras. No obstante, este modelo de tres fases no hace referencia a la importancia de otras estructuras corticales en la función memorística.
Córtex prefrontal
El córtex prefrontal (CPF) lateral es esencial para el recuerdo de los detalles contextuales de una experiencia determinada, más que para la formación misma de recuerdos.[8] También está relacionado en mayor medida con la memoria episódica que con la memoria semántica, en la que en cualquier caso también desempeña un pequeño papel.[9]
Mediante estudios realizados con TEP y estimulación cerebral a través de palabras, Endel Tulving descubrió que el recuerdo es un proceso automático.[10] También es un hecho bien documentado que se encuentra una asimetría hemisférica relacionada con el córtex prefrontal: al codificar los recuerdos, se actuiva el CPF dorsolateral izquierdo, y durante la recuperación de recuerdos el que se activa es el CPF dorsolateral derecho.[10]
Otros estudios han demostrado también que el CPF se encuentra muy involucrado en la conciencia autonoética,[11] responsable de las experiencias de recuerdo que incluyen al sí mismo a través del tiempo, es decir, capacidades de "viaje mental a través del tiempo" características de la memoria episódica, como recordar una experiencia personal del pasado y sus circunstancis específicas.
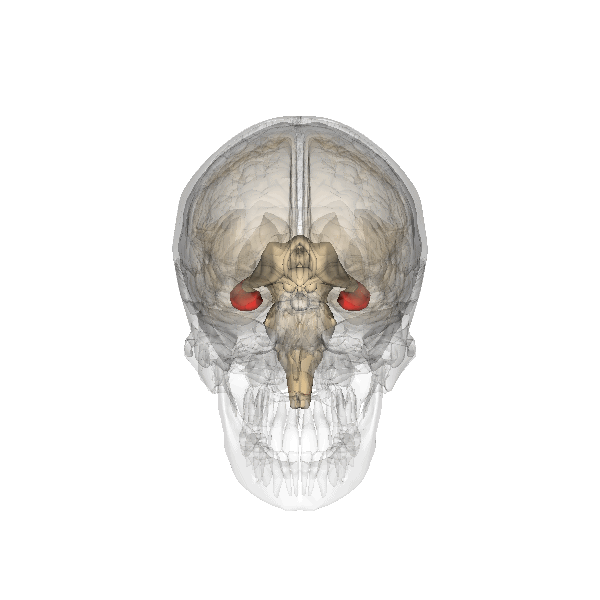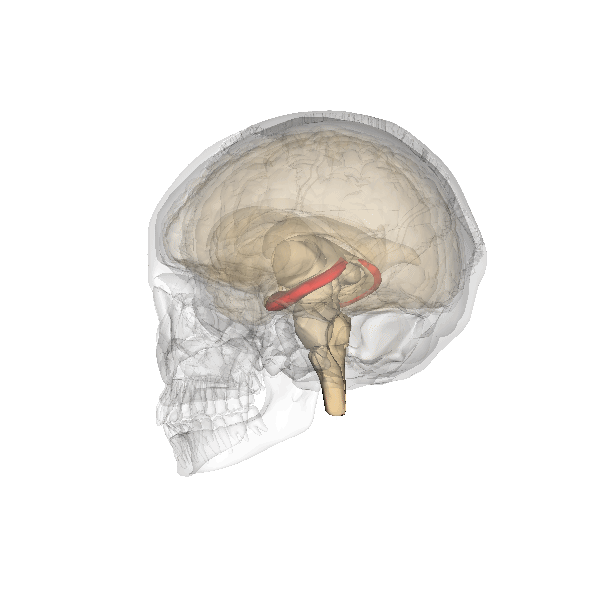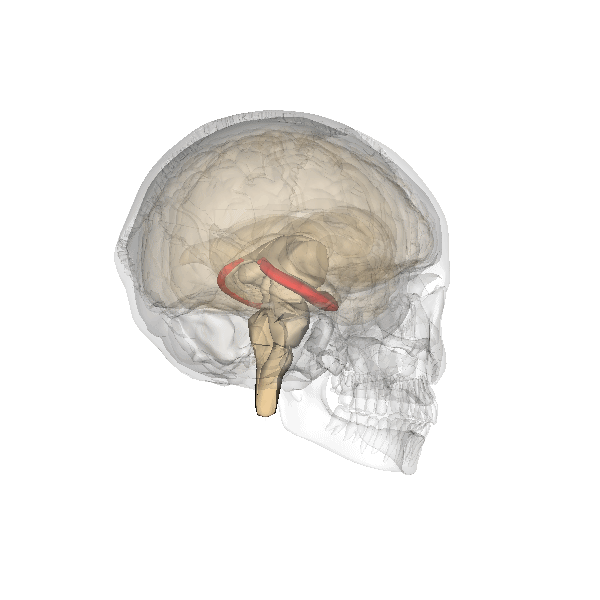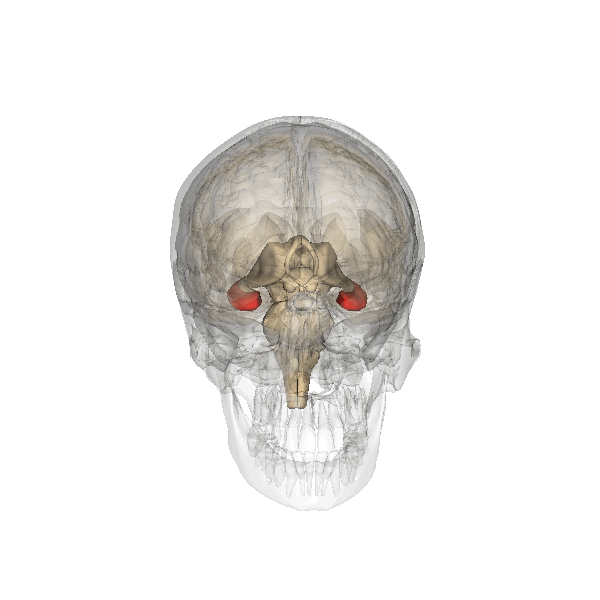
 La amígdala cerebral, destacada en rojo.
La amígdala cerebral, destacada en rojo.Amígdala
Se cree que la amígdala se encuentra implicada en la recuperación de los recuerdos que tienen una carga emocional asociada. La mayor parte de las pruebas al respecto proceden de la investigación de un fenómeno conocido como «recuerdos tipo “flashbul”». Este tipo de recuerdos, relacionados con eventos con un gran contenido emocional, poseen niveles de detalle y persistencia mucho mayores que en el caso de los recuerdos normales. Estos recuerdos se han asociado a un alto grado de activación de la amígdala.[12] Estudios recientes elaborados sobre casos de pacientes con la amígdala dañada sugieren que esta estructura está más relacionada con los recuerdos asociados al conocimiento general que a la información más específica.[13] [14]
Otras estructuras implicadas
Se ha encontrado una activación cerebral en el área del diencéfalo en el momento es que se recupera un recuerdo remoto.[9] Por otra parte, el lóbulo occipital, el lóbulo temporal ventral y el giro fusiforme desempeñan un papel en la formación de recuerdos.[8]
Estudios sobre lesiones
Los estudios de lesiones cerebrales son una práctica habitual de la investigación en neurociencia cognitiva. Las lesiones pueden ocurrir de forma natural por causas de trauma o enfermedad, o pueden ser quirúrgicamente inducidas por los investigadores. En el estudio de la memoria declarativa, el hipocampo y la amígdala son dos estructuras frecuentemente examinadas mediante esta metodología.
Estudios de lesiones hipocampales
 El laberinto de agua de Morris.
El laberinto de agua de Morris.La tarea de navegación de Morris se utiliza para comprobar los mecanismos implicados en el aprendizaje espacial en ratas.[15] En esta prueba, las ratas aprenden a escapar de una piscina nadando hacia una plataforma sumergida bajo la superficie del agua. Las referncias visuales que rodean la piscina (por ejemplo, una silla o una ventana) ayudan a la rata a localizar la plataforma a medida que se van sucediendo los ensayos. El uso que hacen las ratas de estas referencias visuales son formas de memoria declarativa.[16] Se suelen observar dos grupos de ratas: un grupo control sin lesiones, y un grupo experimental con lesiones hipocampales. En esta tarea, creada por Morris et. al., las ratas se sitúan en la piscina en la misma posición a lo largo de doce ensayos. Cada ensayo es cronometrado, y se graba el recorrido realizado por las ratas hasta encontrar la plataforma y salir del laberinto. En estas condiciones, tanto las ratas sanas como las que tienen lesiones hipocampales aprenden a encontrar la plataforma. Pero si se cambia el lugar de salida, las ratas lesionadas suelen fracasar en la localización de la salida. Por el contrario, las ratas del grupo control son capaces de encontrar fácilmente la salida ayudándose de las pistas que dan las referencias visuales externas adquiridas durante los ensayos de aprendizaje.[15] Esto demuestra la implicación del hipotálamo en la memoria declarativa.[16]
En la Tarea de Reconocimiento Olor-olor desarrollada por Bunsey y Eichenbaum, se propicia un encuentro entre dos ratas (un "sujeto" y un "demostrador"). El demostrador, tras haber ingerido un tipo específico de comida, interactúa con la rata sujeto, que percibe el olor de la comida en el aliento de su compañera. En ese momento, los experimentadores le ofrecen a la rata sujeto una elección entre dos comidas diferentes, una de las cuales es la que ha comido previamente la rata demostrador. Los resultados obtenidos fueron que, en los casos en los que no hay una demora temporal entre la reunión con la rata y la elección de la comida, tanto las ratas sanas como las que presentan lesiones hipocampales se decantan por la comida que les resulta familiar. No obstante, si la tarea de elección de comida se presenta 24 horas después de haberse producido el encuentro, las ratas control siguen presentando una preferencia por la comida cuyo olor recuerdan, mientras que las ratas lesionadas no muestran preferencia alguna, decantándose por una u otra en un número similar de ocasiones.[17] Este fenómeno puede atribuirse a la incapacidad de almacenar recuerdos episódicos debida a la lesión hipocampal. Los efectos observados en este estudio también pueden encontrarse en los casos de seres humanos con amnesia, lo que indica el papel del hipocampo en el desarrollo de recuerdos episódicos que pueden generalizarse a nuevas situaciones percibidas como similares.[16]
El paciente H.M. presentaba una extirpación parcial del lóbulo temporal medial, lo que dio como resultado una amnesia anterógrada, es decir, una pérdida de la capacidad de crear recuerdos nuevos.[18] La memoria declarativa a largo plazo se vio muy afectada, incluyendo la capacidad para adquirir nuevos recuerdos o conocimientos semánticos.[19] La disociación encontrada en este caso entre la adquisición de recuerdos declarativos y otros tipos de aprendizaje se apreció en primer lugar en el caso del aprendizaje motor.[20] La memoria declarativa de H.M. no era funcional, como se pudo comprobar cuando el paciente realizó la tarea de primado (o priming) de repetición. Su rendimiento fue mejorando a lo largo de los sucesivos ensayos, pero sus resultados siempre fueron inferiores a los obtenidos por los sujetos control (sin lesiones).[21] En el caso de H.M., se observaron los mismos resultados al investigar otros tipos de funciones memorísticas básicas, como el recuerdo, la evocación y el reconocimiento.[18] Las lesiones no peden considerarse en términos de todo o nada; en el caso del paciente H.M. no se produjo una pérdida de toda memoria o reconocimiento: aunque la memoria declarativa se vio seriamente dañada, el paciente mantenía un sentido del yo y recuerdos no declarativos desarrollados después de la lesión.[22]
Estudios de lesiones de la amígdala
Adolph, Cahill y Schul realizaron un estudio que demostró que la activación emocional facilita la codificación de material en la memoria declarativa a largo plazo.[23] Seleccionaron a dos sujetos con daño bilateral de la amígdala, así como seis sujetos control, y seis sujetos con otro tipo de daño cerebral. A todos los sujetos participantes se les presentó una serie de doce imágenes narradas. Estas imágenes variaban en el grado de emoción que elicitaban. Así, las imágenes 1 a 4, y las imágenes 9 a 12 presentaban un contenido no emocional. Las imágenes 5 a 8 contenían material emotivo, especialmente la número 7 (una imagen de las piernas quirúrgicamente reparadas de la víctima de un accidente de coche).[23]
Los sujetos con daño bilateral de la amígdala no recordaron mejor la imagen número 7 que el resto de imágenes. En cambio, el resto de participantes la recordaban de forma notablemente superior y con mucho mayor detalle que el resto de imágenes más neutras.[23] Esto demuestra que la amígdala es importante en la facilitación de la codificación de conocimientos declarativos relacionados con estímulos de alto contenido emocional, pero no tiene esa influencia en la codificación de conocimientos relacionados con estímulos emocionalmente neutros.[24]
Factores que afectan a la memoria declarativa
Estrés
El estrés provoca un gran impacto en la formación de recuerdos declarativos. Lupien et. al. realizaron un estudio en tres fases. La primera fase consistía en la memorización de series de palabras; la segunda fase suponía participar en una situación estresante (hablar en público) o bien en una no estresante (una tarea de atención); y en la tercera fase los participantes debían recordar las palabras que habían aprendido durante la primera fase. Si los participantes recordaban las palabras significaría que se habían formado recuerdos declarativos. Los participantes que en la segunda fase habían sido sometidos a la situación estresante mostraron un rendimiento inferior en esta tercera fase de recuerdo. Esto demostró que el estrés había empeorado la capacidad de los participantes para almacenar conocimientos declarativos concretos.[25] Los participantes que habían participado en la situación no estresante recordaban con facilidad las series de palabras aprendidas durante la fase 1.
El trastorno de estrés post-traumático surge tras una exposición a sucesos traumáticos que provoquen miedo, horror y sentimientos de indefensión, como los sucesos de daño corporal o amenazas de sufrirlo, así como las experiencias de riesgo para la vida de uno mismo u otra persona cercana.[26] El estrés crónico en el TSPT contribuye a una reducción del volumen del hipocampo, así como a déficits observados en la memoria declarativa.[27]
Factores neuroquímicos cerebrales del estrés
En el cerebro, los glucocorticoides (GCs) modulan la capacidad del hipocampo y el córtex prefrontal para procesar recuerdos.[28] El Cortisol es uno de los GC más comunes en el cuerpo humano, y la hidrocortisona (un derivado del cortisol) disminuye la actividad cerebral en estas áreas durante el proceso de recuperación de recuerdos declarativos.[28]
Durante la fase de estrés se producen aumentos en los niveles de cortisol, y por este motivo, los casos de estrés a largo plazo empeoran el rendimiento de la memoria declarativa.[28] Un estudio realizado por Damoiseaux et. al. evaluó el efecto de los glucocorticoides en el lóbulo temporal medial y el córtex prefrontal en varones jóvenes. Encontraron que administrando GCs a los participantes una hora antes de la tarea, se encontraba un empeoramiento en el rendimiento a la hora de realizar una prueba de recuerdo libre de palabras, sin importar si la administración de los GCs tenía lugar antes o después del aprendizaje del material.[28] A pesar de que no se conoce exactamente cómo influyen los GCs en el rendimiento memorístico, existen receptores de glucocorticoides en el hipocampo y en el córtex prefrontal que indican que estos centros son objetivo de la circulación hormonal.[28] No obstante, se sabe que la cortisona empeora la función mnésica a través de la reducción del flujo sanguíneo en el giro parahipocampal derecho, el córtex visual izquierdo y el cerebelo.[28]
Es necesario reseñar que este estudio sólo incluyó la participación de sujetos masculinos, lo que puede ser significativo, ya que los esteroides sexuales pueden tener efectos diferentes en respuesta a la administración de cortisol. Hombres y mujeres responden de formas diferentes a los estímulos emocionales, y esto puede afectar a los niveles de cortisol. Además, este estudio fue el primer estudio de imagen por resonancia magnética funcional que incluía la participación de GCs, y es necesario que se realicen más investigaciones para respaldar los resultados obtenidos.[28]
Consolidación de la memoria declarativa durante el sueño
Muchos investigadores opinan que el sueño juega un papel activo en la consolidación de la memoria declarativa. Concretamente, las propiedades únicas del sueño favorecen la reactivación de los recuerdos más recientemente aprendidos. Por ejemplo, se ha sugerido que el mecanismo central para la consolidación de la memoria declarativa durante el sueño es la reactivación de las representaciones de los recuerdos en el hipocampo.[29] Algunos estudios realizados con ratas en tareas de aprendizaje de rutas de escape de laberintos demostraron que las agrupaciones neuronales hipocámpicas implicadas en la codificación de información espacial, son reactivadas en el mismo orden temporal.[30] Del mismo modo, la tomografía por emisión de positrones (TEP) ha revelado una reactivación del hipocampo durante la fase de sueño de ondas lentas (fases 3 y 4 del sueño NMOR) después de una tarea de aprendizaje espacial.[31] En conjunto, estos estudios demuestran que los aprendizajes recientes se reactivan durante el sueño, ayudando a la consolidación de nuevas huellas de memoria.[32] En suma, se han identificado tres fases del sueño durante las cuales tiene lugar la consolidación de los aprendizajes declarativos: el sueño de ondas lentas, el sueño de transición, y el sueño MOR.
Sueño de ondas lentas
El sueño de ondas lentas, a menudo llamado sueño profundo, desempeña el papel principal en la consolidación de la memoria declarativa, y existe una gran cantidad de evidencias al respecto. Como muestra, un estudio encontró que durante las primeras tres horas y media de sueño se producen los mayores avances en el desempeño posterior en tareas de recuerdo, porque se trata de una fase dominada por el sueño de ondas lentas. El incremento de horas de sueño a partir de este punto no redunda en mejora alguna sobre el nivel inicial de recuerdo. Así, este estudio sugiere que el sueño completo puede no resultar importante para un desempeño óptimo de la memoria.[33] Otro estudio muestra que la gente que experimenta sueño de ondas lentas durante la primera mitad de su ciclo de sueño, comparada con los sujetos que no lo hacen, muestran un mejor recuerdo de la información.[34]
Otra evidencia al respecto de la influencia del sueño de ondas lentas en la consolidación de la memoria declarativa es el hallazgo de que las personas con patologías del sueño como insomnio, muestran una reducción en el tiempo de sueño de ondas lentas y un empeoramiento de la consolidación de los recuerdos declarativos durante el sueño.[35] Otro estudio ha puesto de manifiesto el hecho de que la gente de mediana edad presentan un peor desempeño en la recuperación de recuerdos que los individuos más jóvenes, lo que indica que el empeoramiento del rendimiento no tiene tanto que ver con la edad en sí misma como con la disminución de tiempo de sueño de ondas lentas que va asociado al envejecimiento.[36] En suma, estos estudios, junto a otros muchos, demuestran la importancia del sueño de ondas lentas en la consolidación de la memoria declarativa.
Husos del sueño
Algunos investigadores opinan que el periodo de transición entre el sueño ligero y el profundo que tiene lugar durante la Fase 2 del ciclo de sueño y se caracteriza por la presencia de unas ondas cerebrales denominadas spindles o husos del sueño, también desempeña un papel importante en la consolidación de recuerdos declarativos.[37] Algunas críticas a este respecto tienen que ver con la correlación que existe entre la presencia de husos del sueño e inteligencia.[38] A pesar de todo, Schaubs y Gruber apuntan que la actividad de los husos del sueño sólo influye sobre el desempeño memorístico relacionado con los aprendizajes más recientes, y no sobre el rendimiento general. Esto sustenta la hipótersis de que los husos del sueño influyen sobre la consolidación de las huellas de memoria recientemente generadas, pero no sobre el rendimiento memorístico general.[39]
Sueño MOR
Existe un relativamente pequeño (aunque creciente) cuerpo de evidencias que apoyan la idea de que el sueño MOR ayuda a la consolidación de recuerdos declarativos asociados a un componente emocional. Por ejemplo, Wagner et. al. compararon la retención memorística de textos emotivos y textos neutros bajo dos condiciones; una de sueño temprano (con predominio de sueño de ondas lentas) y otra de sueño tardía (con predominio de sueño MOR).[40] El resultado fue que el sueño mejoró el rendimiento en retención memorística de textos con contenido emotivo sólo durante la fase de sueño tardío (con predominio de sueño MOR). Análogamente, Hu & Stylos-Allen et al. realizaron un estudio con imágenes emotivas y neutras, y llegaron a la misma conclusión: el sueño MOR facilita la consolidación de los recuerdos declarativos de contenido emocional.[41]
Algunas críticas
No obstante, la teoría de que el sueño desempeña un papel activo en la consolidación de la memoria declarativa no es compartida por todos los investigadores. Por ejemplo, Ellebogen et al. argumentan que el sueño protege a la memoria declarativa de posibles interferencias, facilitando así la consolidación.[42] Por su parte, Wixted opina que el único papel del sueño en la consolidación de recuerdos declarativos se limita a crear las condiciones ideales para que esta consolidación tenga lugar.[43] Por ejemplo, al despertar, una persona se ve asaltada por multitud de estímulos que pueden interferir en la consolidación de la información previamente aprendida. En cambio, durante el sueño, esta interferencia es mínima, y los recuerdos pueden ser consolidados sin obstáculo alguno. En suma, este punto de vista sugiere que el sueño proporciona las condiciones ideales para que se de una consolidación a decuada de los recuerdos, pero no juega un papel activo en este proceso.[32]
Véase también
Referencias
- ↑ Ullman MT. (2004). «Contributions of memory circuits to language: the declarative/procedural model» (en inglés, PDF). Cognition 92 (1-2): pp. 231–270. doi:. ISSN 0010-0277. http://brainlang.georgetown.edu/PUBS/Ullman_Cognition_04.pdf. Consultado el 2 de abril de 2011.
- ↑ a b c d Tulving E. (1972). E. Tulving, W. Donaldson. ed. «Episodic and semantic memory» (en inglés, PDF). Organization of Memory: pp. 381–403. http://www.garfield.library.upenn.edu/classics1987/A1987K827500001.pdf. Consultado el 2 de abril de 2011.
- ↑ Aristóteles (1936). «Acerca del alma (De Anima)». En Jeffrey Henderson. Aristóteles, Volumen 8. Londres: Loeb Classical Library. http://www.amazon.com/Aristotle-Naturalia-Breath-Classical-Library/dp/0674993187/ref=cm_lmf_tit_9#reader_0674993187. Consultado el 2 de abril de 2011.
- ↑ Ebbinghaus, H. (1885). Memory: A Contribution to Experimental Psychology. Nueva York: Teachers College, Columbia University. http://psychclassics.yorku.ca/Ebbinghaus/index.htm. Consultado el 2 de abril de 2011.
- ↑ Graf, P., & Schacter, D. L. (1985). «Implicit and explicit memory for new associations in normal and amnesic subjects» (en inglés, PDF). Journal of Experimental Psychology: Learning, Memory, and Cognition 11 (3): pp. 501-518. doi:. PMID 3160813. http://www.deepdyve.com/lp/psycarticles-reg/implicit-and-explicit-memory-for-new-associations-in-normal-and-Rgr6d0RJku. Consultado el 2 de abril de 2011.
- ↑ a b Eichenbaum, Howard (1997). «Declarative memory: Insights from cognitive neurobiology» (en inglés). Annual Review of Psychology 48: pp. 547-572. doi:. ISSN 0066-4308. PMID 9046568.
- ↑ a b c d e Eichenbaum, Howard (2001). «The Hippocampus and Declarative Memory: Cognitive Mechanisms and Neural Codes» (en inglés, PDF). Behavioural Brain Research 127 (1-2): pp. 199-207. doi:. PMID 11718892. http://www-cogneuro.bu.edu/publications/Eichenbaum%20%282001%292.pdf. Consultado el 3 de abril de 2011.
- ↑ a b c d Noa Ofen, Yun-Ching Kao, Peter Sokol-Hessner, Heesoo Kim, Susan Whitfield-Gabrieli y John D. E. Gabrieli (2007). «Development of the Declarative Memory System in the Human Brain» (en inglés). Nature Neuroscience 10 (9): pp. 1198-1205. doi:. PMID 17676059.
- ↑ a b Graham, S., Levine, B. (noviembre 2004). «The Fundamental Neuroanatomy of Episodic and Semantic Autobiographical Remembering: A Prospective Functional MRI Study» (en inglés). Journal of Cognitive Neuroscience 16 (9): pp. 1633-1646. doi:.
- ↑ a b Craik, F.I.M., Houle, S. (1994). «Role of Prefrontal Cortex in Human Episodic Memory: Lessons From PET Studies» (en inglés). Biol. Psychiatry 42: pp. 75S-76S.
- ↑ Stuss, D.T., Tulving, E. (mayo 1997). «Toward a Theory of Episodic Memory: The Frontal Lobes & Autonoetic Consciousness» (en inglés, PDF). Psychological Bulletin 121 (3): pp. 331-354. ISSN 00332909. PMID 9136640. http://cogweb.ucla.edu/Discourse/Narrative/Wheeler-97.pdf. Consultado el 3 de abril de 2011.
- ↑ Sharot T., Martorella E.A., Delgado M.R., Phelps E.A. (enero 2007). «How personal experience modulates the neural circuitry of memories of September 11» (en inglés, PDF). Proc. Natl. Acad. Sci. USA 104 (1): pp. 389–394. doi:. ISSN 0027-8424. PMC 1713166. http://www.psych.nyu.edu/phelpslab/papers/2007_PNAS_V104No1.pdf. Consultado el 3 de abril de 2011.
- ↑ Adolphs R., Tranel D., Buchanan T.W. (Abril 2005). (en inglés, PDF)Nature Neuroscience 8 (4): pp. 512–518. doi:. PMID 15735643. http://www.slu.edu/colleges/AS/cnslab/publications/AltNarr_NN.pdf. Consultado el 3 de abril de 2011.
- ↑ Adolphs R., Denburg N.L., Tranel D. (Octubre 2001). «The amygdala’s role in long-term declarative memory for gist and detail» (en inglés). Behav. Neurosci. 115 (5): pp. 983–992. doi:. PMID 11584931.
- ↑ a b Eichenbaum, H., Stewart, C. & Morris, R. G. M. (Noviembre 1990). «Hippocampal representation in spatial learning» (en inglés, PDF). the Journal of Neuroscience 10 (11): pp. 3531–3542. ISSN 0270-6474. PMID 2230943. http://www.jneurosci.org/content/10/11/3531.full.pdf+html. Consultado el 3 de abril de 2011..
- ↑ a b c Eichenbaum, H. (Octubre 2000). «A cortical-hippocampal system for declarative memory» (en inglés, PDF). Nature Reviews Neuroscience 1 (1): pp. 41-50. doi:. PMID 11252767. http://www.bu.edu/cogneuro/publications/Eichenbaum%20%282000%29.pdf. Consultado el 3 de abril de 2011.
- ↑ Bunsey, M. & Eichenbaum, H. (1995). «Selective damage to the hippocampal region blocks long term retention of a natural and nonspatial stimulus-stimulus association» (en inglés). Hippocampus 5 (6): pp. 546–556. doi:. PMID 8646281..
- ↑ a b Corkin, S. (Febrero 2002). (en inglés, PDF)Nature Reviews. Neuroscience 3 (2): pp. 153-160. doi:. ISSN 1471-003X. PMID 11836523. http://homepage.mac.com/sanagnos/corkin2002.pdf. Consultado el 3 de abril de 2011.
- ↑ Gabrieli, J. D. E., Cohen, N. J. y Corkin, S (Abril 1988). «The impaired learning of semantic knowledge following bilateral medial temporal-lobe resection» (en inglés). Brain and Cognition 7 (2): pp. 157–177. doi:. ISSN 0278-2626. PMID 3377896.
- ↑ Corkin, S. (Septiembre 1968). «Acquisition of motor skill after bilateral medial temporal-lobe excision» (en inglés, PDF). Neuropsychologia 6 (3): pp. 225–264. doi:. http://web.mit.edu/bnl/pdf/Corkin%201968.pdf. Consultado el 3 de abril de 2011.
- ↑ Milner, B., Corkin, S. y Teuber, H.-L. (Septiembre 1968). «Further analysis of the hippocampal amnesic syndrome: 14-year follow-up study of H.M.» (en inglés, PDF). Neuropsychologia 6 (3): pp. 215–234. doi:. http://www.psychology.uiowa.edu/Faculty/Freeman/Milner_68.pdf. Consultado el 3 de abril de 2011.
- ↑ Aggleton, J. P. y Brown, M. W. (Junio 1999). «Episodic memory, amnesia, and the hippocampal–anterior thalamic axis» (en inglés, PDF). Behavioral and Brain Sciences 22 (3): pp. 425–489. doi:. PMID 11301518. http://www.cf.ac.uk/psych/home2/papers/aggleton/Behav%20&%20Brain%20Sci%20-%2022%20-%20425-444.pdf. Consultado el 3 de abril de 2011.
- ↑ a b c Adolphs, R., Cahill, L., Schul, R., & Babinsky, R. (Septiembre-Octubre 1997). «Impaired declarative memory for emotional material following bilateral amygdala damage in humans» (en inglés, PDF). Learning and Memory 4 (3): pp. 291-300. doi:. PMID 10456070. http://learnmem.cshlp.org/content/4/3/291.full.pdf+html. Consultado el 3 de abril de 2011.
- ↑ Babinsky, R., Calabrese, P., Durwen, H., Markowitsch, H., & Brechtelsbuauer, D. (1993). «The possible contribution of the amygdala to memory» (en inglés). Behavioural Neurology 6 (3): pp. 167-170. ISSN 0953-4180.
- ↑ Lupien, S., Gaudreau, S., Tchiteya, B., Maheu, F., Sharma, S., Nair, N., et al. (Julio 1997). «Stress-Induced Declarative Memory Impairment in Healthy Elderly Subjects: Relationship to Cortisol Reactivity» (en inglés). The Journal of Clinical Endocrinology & Metabolism 82 (7): pp. 2070-2075. doi:. PMID 9215274. http://jcem.endojournals.org/cgi/content/full/82/7/2070. Consultado el 3 de abril de 2011.
- ↑ Cabeza, R., y LaBar, K. S. (enero 2006). «Cognitive neuroscience of emotional memory» (en inglés, PDF). Nature Reviews Neuroscience 7 (1): pp. 54-64. doi:. ISSN 1471-003X. PMID 16371950. http://www.people.vcu.edu/~mreimers/SysNeuro/LaBar%20-%20Emotional%20memory%20neuroscience.pdf. Consultado el 3 de abril de 2011.
- ↑ Baker, D. G. et al. (Mayo 2005). «Higher levels of basal serial CSF cortisol in combat veterans with posttraumatic stress disorder» (en inglés). The American Journal of Psychiatry 162 (5): pp. 992–994. PMID 15863803. http://ajp.psychiatryonline.org/cgi/content/full/162/5/992. Consultado el 3 de abril de 2011.
- ↑ a b c d e f g Damoiseaux, J.S., Elzinga, B.M. (Junio 2007). «Glucocorticoids Decrease Hippocampal and Prefrontal Activation during Declarative Memory Retrieval in Young Men» (en inglés, PDF). Brain Imaging and Behaviour 1 (1-2): pp. 31-41. doi:. PMID 19946603. https://openaccess.leidenuniv.nl/bitstream/1887/16156/9/02.pdf. Consultado el 3 de abril de 2011.
- ↑ McClelland, J.L., McNaughton, B.L., y O’Reilly, R.C. (Julio 1995). «Why there are complementary learning systems in the hippocampus and neocortex: Insights from the successes and failures of connectionist models of learning and memory» (en inglés, PDF). Psychol. Rev. 102 (3): pp. 419–457. PMID 7624455. ftp://grey.colorado.edu/pub/oreilly/papers/McClellandMcNaughtonOReilly95.pdf. Consultado el 3 de abril de 2011.
- ↑ Daoyun Ji y Matthew A. Wilson (2007) (en inglés, PDF). Coordinated memory replay in the visual cortex and hippocampus during sleep. 10. pp. 100-107. doi:. ISSN 10976256. PMID 17173043. http://www.phys.mcw.edu/documents/RewindingthetapeduringsleeptoPresent.pdf. Consultado el 3 de abril de 2011.
- ↑ P. Peigneux et al. (2004). «Memory processing during sleep mechanisms and evidence from neuroimaging studies» (en inglés, PDF). Psychologica belgica 44 (1-2): pp. 121-142. ISSN 0033-2879. http://dev.ulb.ac.be/ur2nf/reprints/Peigneux%28B%29_PyschologicaBelgica_44%281-2%2904.pdf. Consultado el 3 de abril de 2011.
- ↑ a b Ellenbogen J., Payne D., et al. (Diciembre 2006). «The role of sleep in declarative memory consolidation: passive, permissive, active or none?» (en inglés, PDF). Neurobiology 16 (6): pp. 716–722. doi:. PMID 17085038. http://dept.wofford.edu/neuroscience/NeuroSeminar/pdfSpring2009/Ellenbogen_2006_Current-Opinion-in-Neurobiology.pdf. Consultado el 4 de abril de 2011.
- ↑ Tucker A., Fishbein W. (Septiembre 2009). «The Impact of sleep duration and subject intelligence on declarative and motor memory performance: how much is enough?» (en inglés, PDF). J. Sleep Res. 18 (3): pp. 304-312. doi:. PMID 19702788. http://moodle.ncku.edu.tw/file.php/30504/Tucker_and_Fishbein_-_2009_-_The_impact_of_sleep_duration_and_subject_intellige.pdf. Consultado el 4 de abril de 2011.
- ↑ Plihal W., Born J. (Julio 1997). «Effects of early and late nocturnal sleep on declarative and procedural memory» (en inglés). Journal of Cognitive Neuroscience 9 (4): pp. 534-547. doi:. ISSN 0898-929X.
- ↑ Backhaus, J., Junghanns, K., Born, J., Hohaus, K., Faasch, F., and Hohagen, F. (Diciembre 2006). «Impaired declarative memory consolidation during sleep in patients with primary insomnia: Influence of sleep architecture and nocturnal cortisol release» (en inglés). Biologycal Psychiatry 60 (12): pp. 1324–1330. doi:. ISSN 0006-3223. PMID 16876140.
- ↑ Backhaus J., Born J., et al. (Mayo 2007). «Midlife decline in declarative memory consolidation is correlated with a decline in slow wave sleep» (en inglés, PDF). Learning & Memory 14 (5): pp. 336-341. doi:. PMID 17522024. PMC 1876757. http://learnmem.cshlp.org/content/14/5/336.full.pdf+html. Consultado el 4 de abril de 2011.
- ↑ Gais S., Molle M., Helms K., Born J. (Agosto 2002). «Learning-dependent increases in sleep spindle density» (en inglés, PDF). J. Neurosci. 22 (15): pp. 6830-6834. PMID 12151563. http://www.jneurosci.org/content/22/15/6830.full.pdf. Consultado el 4 de abril de 2011.
- ↑ Schabus M., Hodlmoser K., Gruber G., Sauter C., Anderer P., Klosch G., Parapatics S., Saletu B., Klimesch W., Zeitlhofer J. (Abril 2006). «Sleep spindle-related activity in the human EEG and its relation to general cognitive and learning abilities» (en inglés, PDF). Eur J Neurosci. 23 (7): pp. 1738-1746. doi:. PMID 16623830. http://web.mit.edu/dmalt/Public/9.10/j.1460-9568.2006.04694.pdf. Consultado el 4 de abril de 2011.
- ↑ Schabus M.; Gruber G.; Parapatics S. et al. (Diciembre 2004). «Sleep spindles and their significance for declarative memory consolidation» (en inglés, PDF). Sleep 27 (8): pp. 1479-1485. ISSN 0161-8105. PMID 15683137. http://www.journalsleep.org/Articles/270804.pdf. Consultado el 4 de abril de 2011.
- ↑ Wagner U., Gais S., Born J. (Marzo-Abril 2001). «Emotional memory formation is enhanced across sleep intervals with high amounts of rapid eye movement sleep» (en inglés, PDF). Learn Mem 8 (2): pp. 112-119. doi:. PMID 11274257. http://learnmem.cshlp.org/content/8/2/112.full.pdf+html. Consultado el 4 de abril de 2011.
- ↑ Hu P., Stylos-Allen M., Walker M.P. (Octubre 2006). «Sleep facilitates consolidation of emotionally arousing declarative memory» (en inglés, PDF). Psychologycal Science 17 (10): pp. 891-898. PMID 17100790. http://webs.wofford.edu/boppkl/courseFiles/Memory/articles/Hu_SleepEmotDeclMem.pdf. Consultado el 4 de abril de 2011.
- ↑ Ellenbogen J.; Hulbert J.; et al. (Julio 2006). «Interfering with Theories of Sleep and Memory: Sleep, Declarative Memory, and Associative Interference» (en inglés, PDF). Current Biology 16 (13): pp. 1290-1294. doi:. PMID 16824917. http://www.sas.upenn.edu/psych/stslab/assets/pdf/Ellenbogen2006.pdf. Consultado el 4 de abril de 2011.
- ↑ Wixted J.T. (2004). «The psychology and neuroscience of forgetting» (en inglés, PDF). Annual Review of Psychology 55: pp. 235-269. doi:. PMID 14744216. http://www.camden.rutgers.edu/~bwhitlow/Courses/Learning/forgetting.pdf. Consultado el 4 de abril de 2011.

Wikimedia foundation. 2010.