- Trirachodon
-
 Trirachodon
TrirachodonRango temporal: Triásico Inferior y Triásico Medio 
Clasificación científica Reino: Animalia Filo: Chordata Clase: Synapsida Orden: Therapsida Suborden: Cynodontia Infraorden: Eucynodontia (sin clasif.): Cynognathia Superfamilia: Tritylodontoidea Familia: Trirachodontidae Género: Trirachodon
Seeley, 1895Especies - T. berryi Seeley, 1895 (tipo)
- T. kannemeyeri Seely, 1895
- T. minor Broom, 1905
- T. browni Broom, 1915
Trirachodon es un género extinto de terápsido cinodonto perteneciente a la familia Tritylodontoidea. Sus fósiles se encontraron en Sudáfrica provenientes del Triásico Inferior y Medio.[1]
Contenido
Descripción
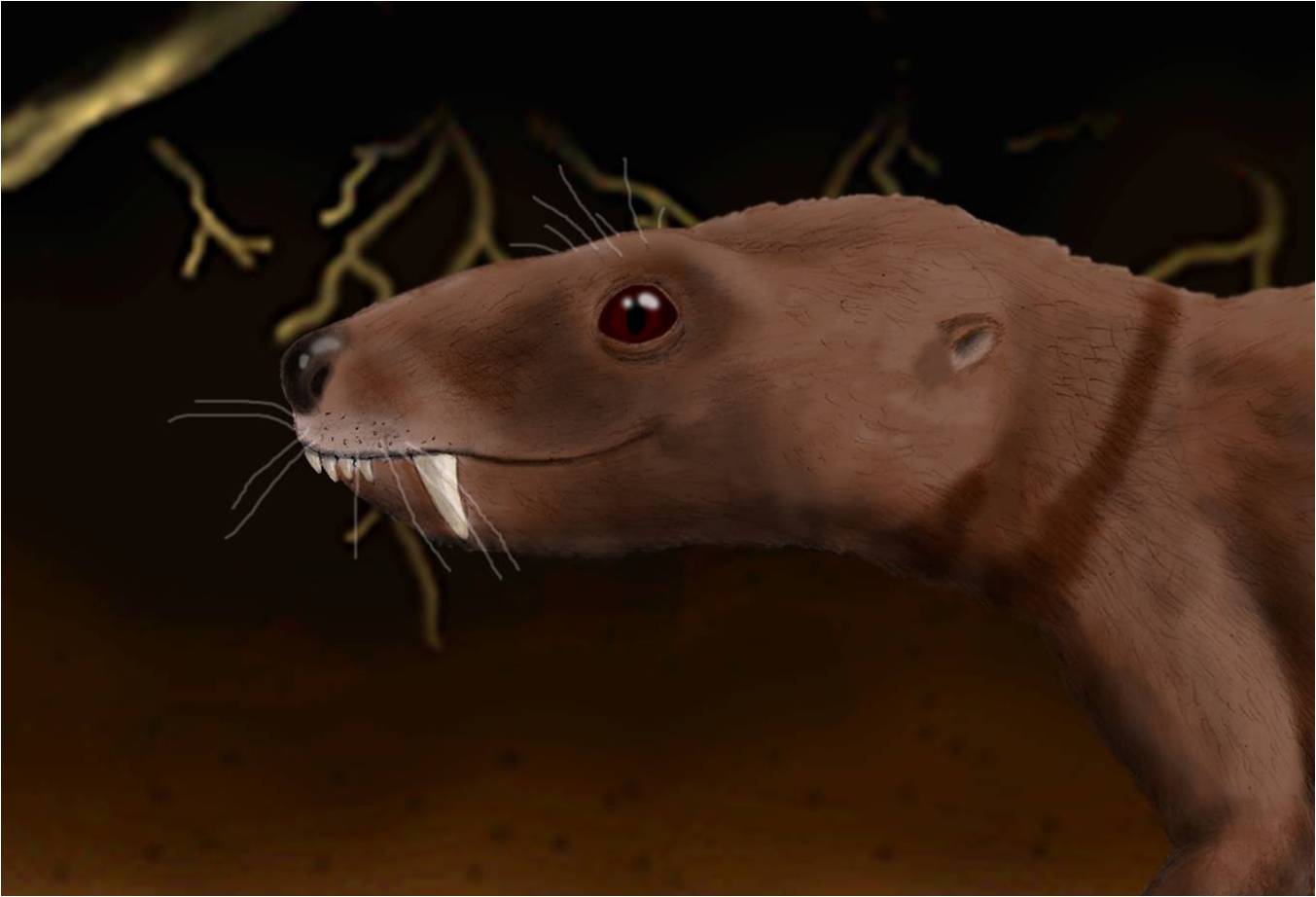
El cráneo de Trirachodon tenía un hocico corto y angosto con una región orbital amplia. El arco cigomático era relativamente delgado.[2] [3] Trirachodon era bastante pequeño para ser un cinodonto, alcanzando solo 50 cm de longitud. Tenía menos diente molariformes que su pariente contemporáneo más cercano Diademodon. Estos dientes tendían a ser más anchos transversalmente que en Diademodon.[4] [5] Un paladas secundario oseo y la oclusión precisa de los postcaninos son las características de Trirachodon que lo hacen similar a los mamíferos.
Especies
La especie tipo es T. berryi, descrita en 1895 a partir de la base de un cráneo. Otros tres especímenes fueron posteriormente asignados a la especie T. kannemeyeri, la cual se distinguía de la especie tipo por la longitud de la base del hocico y el número de dientes postcaninos. Estas diferencias se consideraron demasiado superfluas para ser asignados a una especie diferente por lo cual T. kannemeyeri se encuentra fuera de uso debido a la posible sinonimia.
Una nueva especie, T. minor, fue descrita por Robert Broom en 1905 para describir un hocico escasamente preservado. Broom posteriormente lo denominó T. browni en 1915, distinguiéndolo de las otras especies por la diferencia de longitud de los molares. En 1932, Broom propuso que T. berryi se asignara an nuevo género, Trirachodontoides. Otra especie de Trirachodon denominada T. angustifrons fue descrita en 1946 a partir de un cráneo angosto hallado en Tanzania, pero se probó posteriormente que correspondía al traversodóntido Scalenodon. En 1972 se sugirió que todas las especies de Trirachodon son sinónimos de la especie tipo excepto T. browni, el cual fue renombrado como Diademodon tetragonus.[6]
Paleobiología
Se cree que Trirachodon tenía una forma de vida fosorial. Rastro de excavaciones complejas encontrados en Sudáfrica y Namibia se le han atribuido al género.[7] Al menos 20 individuos se encontraron en uno de los complejos. La entrada tiene un desnivel poco pronunciado y cuentan con suelos y techos abovedados bilobulados. El piso en los niveles inferiores son menos bilobulados. Las cavernas normalmente terminan en ductos angostos. Los túneles tienden a hacerse más curvos a mayor profundidad con cámaras que conectan en ángulo recto con el túnel principal. Trirachodon presentaba una postura semierecta en los miembros posteriores como adaptación para mantener una locomoción eficiente dentro de los túneles.[8] [9] Las cortezas relativamente gruesas observadas en los huesos también le proporcionaban resistencia extra a miembros durante las excavaciones. Se cree que la madriguera con su ocupantes fue preservada a causa de una inundación repentina que ocupó los túneles sin permitir a sus ocupantes evacuar a tiempo.
Muchas características de las madrigueras sugieren que fueron usadas como albergue para una colonia. La entrada espaciosa pudo ser ocupada por muchos individuos y los túneles accesorios y sus cámaras respectivas pudieron ser construidas por un solo animal. Los pisos compactos y los túneles bilobulados sugieren que fueron utilizados por muchos individuos que transitaban continuamente a través de ellos.[10]
Una forma de vida colonial para Trirachodon sugiere comportamientos sociales complejos que se creían únicos entre los sinápsidos del Cenozoico, y uno de las primeras evidencias de cohabitación en un complejo de túneles por parte de algún tetrápodo (un molde parcial de madriguera asociado con Thrinaxodon liorhinus, a mostrado que estos predaron estas cuevas por algunos millones de años.[11] ) Se han sugerido muchas razones para explicar el comportamiento de Trirachodon, las cuales incluyen protección de los depredadores, sitios de reproducción y crianza de los jóvenes, y termoregulación.
Estudios histológicos óseos recientes de muchos especímenes de Trirachodon han mejorado la compresión del origen y estilo de vida de estos animales. Existe evidencia de anillos de crecimiento óseo con tasas de crecimiento fuertemente influenciadas por las fluctuaciones de las condiciones climáticas de su medio ambiente.[12]
Referencias
- ↑ Rubidge, B.S. (ed.) 1995. Biostratigraphy of the Beaufort Group (Karoo Supergroup). 46 pp. Council for Geoscience, Pretoria.
- ↑ Seeley, H. G. (1895). Researches on the structure, organization and classification of the fossil Reptilia. On Diademdon. Philosophical Transactions of the Royal Society of London B 185:1029–1041.
- ↑ Kemp, T. S. (1982). Mammal−like Reptiles and the Origin of Mammals. 363 pp. Academic Press, London.
- ↑ Seeley, H. G. (1895). On the structure, organization and classification of the fossil Reptilia III. On Trirachodon. Philosophical Transactions of the Royal Society of London B 186:48–57.
- ↑ Crompton, A. W. and Ellenberger, F. (1957). On a new cynodont from Molteno Beds and origin of tritylodontids. Annals of the South African Museum 44:1–14.
- ↑ Abdala, F., Neveling, J. and Welman, J. (2006). A new trirachodontid cynodont from the lower levels of the Burgersdorp Formation (Lower Triassic) of the Beaufort Group, South Africa and the cladistic relationships of Gondwanan gomphodonts. Zoological Journal of the Linnean Society 147:383-413.
- ↑ Smith, R. and Swart, R. (2002). Changing fluvial environments and vertebrate taphonomy in response to climatic drying in a Mid−Triassic rift valley fill: The Omingonde Formation (Karoo Supergroup) of Central Namibia. Palaios 17(3):249–267.
- ↑ Carrier, D. R. (1987). The evolution of locomotor stamina in tetrapods: Circumventing a mechanical constraint. Paleobiology 13:326–341.
- ↑ Pough, F. H., Heiser, J. B., and McFarland, W. N. (1996). Vertebrate Life. 798 pp. Prentice−Hall, New Jersey.
- ↑ Groenewald, G. H., Welman, J., and Maceachern, J. A. (2001). Vertebrate Burrow Complexes from the Early Triassic Cynognathus Zone (Driekoppen Formation, Beaufort Group) of the Karoo Basin, South Africa. Palaios 16(2):148-160.
- ↑ Damiani, R., Modesto, S., Yates, A., and Neveling, J. (2003). Earliest evidence of cynodont burrowing. Proceedings of the Royal Society B 270:1747–1751
- ↑ Botha, J. and Chinsamy, A. (2004). Growth and life habits of the Triassic cynodont Trirachodon, inferred from bone histology. Acta Palaeontologica Polonica 49(4):619-627.
Enlace externos
- Trirachodon in the Paleobiology Database
Categorías:- Cynodontia
- Sinápsidos del Triásico
Wikimedia foundation. 2010.