- Caspasa
-
Caspasa
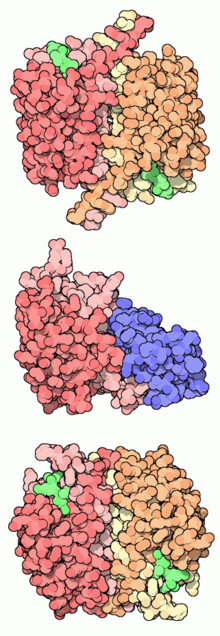 Arriba, caspasa1 (en verde, inhibidores asociados): centro, caspasa9 (en azul, una proteína inhibidora); abajo, caspasa3.
Arriba, caspasa1 (en verde, inhibidores asociados): centro, caspasa9 (en azul, una proteína inhibidora); abajo, caspasa3.
Las caspasas son un grupo de proteínas perteneciente al grupo de las cisteín-proteasas, caracterizadas por presentar un residuo de cisteína que media la ruptura de otras proteínas. En el caso de las caspasas el corte se produce al nivel de un residuo de aspartato de lo que deriva su nombre (cisteinil-aspartato proteasas). Las caspasas son mediadores esenciales de los procesos de apoptosis, la muerte celular programada, de especial relevancia en los procesos morfogenéticos del desarrollo embrionario. Algunas caspasas también están implicadas en procesos de maduración proteica como en el caso de mediadores del sistema inmune del tipo de la interleuquinas. Por estos motivos, fallos en los procesos mediados por caspasas son algunos de los principales responsables del desarrollo de tumores y enfermedades autoinmunes, así como una excesiva activación se cree vinculada con enfermedades como el Alzheimer.
Contenido
Clasificación y estructura
La familia de las caspasas está compuesta por 14 proteínas de las que 11 se corresponderían con proteínas humanas. La primera en descubrirse fue la enzima convertidora de interleuquina-1-b (ICE), conocida desde entonces como caspasa-1 y responsable de la maduración de la pro-interleuquina-1-b a su forma pro-inflamatoria y biológicamente activa. La importancia de las caspasas como responsables de los procesos de apoptosis fue establecida por Robert Horvitz y colaboradores al encontrar que el producto del gen ced-3, una cisteín-proteasa análoga de ICE, estaba implicado en los procesos de muerte celular durante el desarrollo de C. elegans. Estudios posteriores permitieron clasificar el resto de la familia de las caspasas en función de su orden de descubrimiento y filogenéticamente en dos grupos, el grupo inflamatorio que estaría formado por los homólogos de ICE como la caspasa-11, y el grupo apoptótico, formado por las relacionadas con ced-3 como las caspasas-3 y -7. Aunque ambos grupos presentan similitudes en cuanto a la especificidad de sustrato, el primero mediaría la maduración de citoquinas pro-inflamatorias y el segundo el procesamiento de productos que desencadenarían cambios celulares incluyendo degradación del ADN, condensación de cromatina y desintegración de la membrana plasmática.
Las caspasas contienen tres dominios: un prodominio N-terminal, una subunidad grande (p20) que contiene el centro activo con cisteína dentro de un motivo conservado QACXG, y una subunidad pequeña (p10) en el C-terminal. Las caspasas son unas de las proteasas más específicas con un requerimiento inusual y absoluto de cortar después de un residuo de ácido aspártico (Asp). El prodominio y la subunidad grande están separados por un lugar de corte con Asp, y la subunidad grande está separada de la pequeña por uno o dos motivos de este tipo. La presencia de Asp en los motivos de corte para la maduración es consistente con la habilidad de las caspasas de autoactivarse o de ser activadas por otras caspasas como parte de una cascada de amplificación.
 Secuencia conservada de las caspasas apoptóticas de Drosophila
Secuencia conservada de las caspasas apoptóticas de DrosophilaAparte de clasificarse en base a su filogenia y función general, las caspasas se pueden clasificar en dos tipos según su función en las diversas cascadas de señalización intracelular en las que median: caspasas iniciadoras y caspasas efectoras. Las caspasas iniciadoras como las caspasas-8 y -9 procesan las formas inactivas de las caspasas efectoras como las caspasas-3 y -7, activándolas. Las caspasas efectoras una vez activadas procesan a su vez otros sustratos proteicos que mediarán en las distintas vías de apoptosis. La iniciación de estas reacciones en cascada está regulada por inhibidores de caspasas.
Regulación y mecanismos de acción
Como ya se ha comentado, las caspasas están reguladas a nivel postraduccional, asegurando así que puedan ser activadas rápidamente. En un primer momento son sintetizadas como zimógenos inactivos (pro-caspasas) con su estructura clásica consistente en un prodominio, una subunidad pequeña y una subunidad grande. Las caspasas iniciadoras poseen un prodominio mayor que las caspasas efectoras que contiene dominios como los de reclutamiento y activación de caspasas (CARD) en el caso de caspasa-2 o caspasa-9 (ver Apaf-1) o efectores de muerte celular (DED) en el caso de caspasa-8 y -10, que le permite interactuar con otras moléculas que regulan su activación. Estas moléculas responden a estímulos, ocasionando el agrupamiento de las caspasas iniciadoras, lo que les permite autoactivarse y así proceder a activar a las caspasas efectoras. En todos los casos estudiados, la enzima madura es un heterotetrámero que contiene dos heterodímeros p20/p10 y dos centros activos. Existen tres mecanismos generales de activación de caspasas:
- Activación por otra caspasa: en base a su estructura en la que un punto de corte de Asp separa el prodominio de p20 y uno o dos separan p20 de p10 se puede suponer que una posibilidad de activación sea la autocatálisis, es decir, la activación de una pro-caspasa exponiéndola a otra previamente activada. Esta estrategia de activación denominada cascada de caspasas es muy utilizada por las células para la activación de las caspasas efectoras de prodominio corto. La cascada de caspasas es un método útil para amplificar e integrar las señales proapoptóticas, pero no pueden explicar cómo se activó la primera caspasa. Existen al menos dos aproximaciones que explican dicha activación.
- Activación inducida por proximidad: la caspasa-8 es la caspasa iniciadora clave en la vía de los receptores de muerte. Después de la unión del ligando, los receptores de muerte como Fas se agregan y forman un complejo de señalización de membrana. Estos complejos reclutan, a través de sus proteínas adaptadoras, varias moléculas de procaspasa-8 con lo que se aumenta la concentración local de zimógeno. En estas condiciones, la baja e intrínseca actividad proteasa de la procaspasa-8 es suficiente para permitir que varias moléculas inactivas se corten mutuamente y se activen unas a otras.
- Asociación con una subunidad reguladora: el mecanismo de activación más complejo es el utilizado por la caspasa-9. Al contrario que en otras caspasas, el procesamiento proteolítico de la procaspasa-9 tiene un efecto mínimo en su activación. El requerimiento clave para la activación de la caspasa-9 es su asociación con un cofactor de proteínas, Apaf-1. También es necesario el citocromo c liberado por la mitocondria. El citocromo c y Apaf-1 se asocian en un proceso ATP dependiente. La oligomerización de Apaf-1 recluta procaspasas-9 formando el apoptosoma. La activación de la caspasa-9 es debida a un cambio conformacional, no a exclusivamente a la proteolisis.
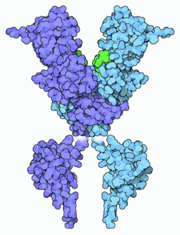 Desoxirribonucleasa activada por una caspasa.
Desoxirribonucleasa activada por una caspasa.En resumen, las caspasas efectoras se activan proteolíticamente por otras caspasas mientras que las caspasas iniciadoras son activadas por interacciones reguladas proteína-proteína. La cascada de caspasas puede ser activada por la granzima B liberada por los linfocitos T citotóxicos (CD8+) y que activa las caspasas-3 y -7, por receptores de muerte celular como Fas, TRAIL o TNF que activan las caspasas-8 y -10 o por el apoptosoma, regulado por la familia Bcl-2 y el citocromo c que activa la caspasa-9. Una vez activada esta cascada, procesos de retroalimentación positiva aseguran que la célula inevitablemente sufrirá apoptosis. Por ejemplo, la caspasa-9 activada por el apoptosoma procesa y activa la caspasa-3 que, además de procesar sus proteínas diana, procesa a la propia caspasa-9, aumentando la concentración de su forma activa.
Algunas de las dianas finales de las caspasas incluyen las láminas nucleares, el sistema de fragmentación de ADN ICAD/DFF45, la poli-ADP-ribosa polimerasa (PARP) y la kinasa PAK2. La contribución exacta de las diferentes caspasas y el procesamiento de sus respectivas dianas a la apoptosis es todavía incierta. Sin embargo, sí se sabe que la inactivación del complejo ICAD/DFF45 mediada por caspasas permite a la proteína CAD entrar en el núcleo y fragmentar el ADN, originando la característica "escalera de ADN" que se observa en células apoptóticas.
Véase también
Referencias
- Cerretti DP, Kozlosky CJ, Mosley B, Nelson N, Van Ness K, Greenstreet TA, March CJ, Kronheim SR, Druck T, Cannizzaro LA, et al. Molecular cloning of the interleukin-1 beta converting enzyme Science. 1992 Apr 3;256(5053):97-100. PMID 8242740.
- Yuan J, Shaham S, Ledoux S, Ellis HM, Horvitz HR. The C. elegans cell death gene ced-3 encodes a protein similar to mammalian interleukin-1 beta-converting enzyme. Cell. 1993 Nov 19;75(4):641-52. PMID 9748481.
- Stennicke HR, Salvesen GS. Properties of the caspases. Biochim Biophys Acta. 1998 Sep 8;1387(1-2):17-31. PMID 9748481.
- Chen M, Wang J. Initiator caspases in apoptosis signaling pathways. Apoptosis. 2002 Aug;7(4):313-9. PMID 12101390.
- Shi Y. Caspase activation: revisiting the induced proximity model. Cell. 2004 Jun 25;117(7):855-8. PMID 15210107.
- Yuan J, Horvitz HR. A first insight into the molecular mechanisms of apoptosis. Cell. 2004 Jan 23;116(2 Suppl):S53-6, 1 p following S59. PMID 15055582.
Categorías: Apoptosis | Proteínas

Wikimedia foundation. 2010.