- Chlamydiae
-
 Chlamydiae
Chlamydiae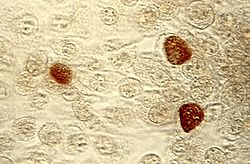
Chlamydia trachomatis intracelular (marrón)Clasificación científica Dominio: Bacteria Filo: Chlamydiae Clase: Chlamydiae Orden: Chlamydiales Familias Las clamidias (Chlamydiae) son un filo de bacterias cuyos miembros son endosimbiontes o patógenos intracelulares obligados.[1] Muchas Chlamydiae coexisten en sus hospedadores específicos cuyo estado asintomático, se cree, proporciona un reservorio natural para estas bacterias.[2]
Todas las Chlamydiae crecen infectando células eucariotas y dentro del hospedero las Chlamydiae toman una forma intracelular replicativa; y en el exterior sobreviven solamente en una forma infecciosa extracelular. Chlamydiae sólo puede proliferar donde haya células hospedadoras, lo que implica dos cosas en el laboratorio: no pueden crecer en medios de cultivo bacterianos y se aíslan más fácilmente mientras están aún dentro de la célula hospedera.
Se han descrito tres especies del filo Chlamydiae que infectan comúnmente a los seres humanos:
- Chlamydia trachomatis, que causa la enfermedad ocular tracoma y linfogranuloma venéreo, una infección de transmisión sexual;
- Chlamydophila pneumoniae, que causa una neumonía atípica;
- Chlamydophila psittaci, que causa psitacosis en loros y enfermedad zoonótica.
Ciclo vital
Chlamydiae agrupa bacterias que pueden ser parásitas o endosimbiontes, dependiendo de la especie y del hospedero eucariota.
La forma infecciosa extracelular se denomina cuerpo elemental (EB, un término prestado de la virología), es denso a los electrones y tiene típicamente un diámetro de 0,2-0,6 μm. La pared del EB se liga con enlaces de disulfuro. Los EBs que han sido ingeridos por las células eucariotas permanecen en general en forma de inclusiones vacuolares (un término de la virología), en donde se reducen los enlaces de disulfuro y los EBs se transforman en cuerpos reticulados (RBs, donde "reticulado" quiere decir homogéneo).
Los RBs tienen un tamaño de hasta 1,5 μm, toman nutrientes de la célula huésped y experimentan múltiples divisiones binarias. La división binaria puede implicar estructuras anulares compuestas por un tipo transitorio de peptidoglicano.[3] Usando la microscopía electrónica, EBs y RBs se pueden ver durante la inclusión y la réplicación. Las inclusiones no experimentan acidificación o fusión lisosomial, no correspondiendo con las vesículas endocíticas canónicas, estando esencialmente disociadas del ciclo endocítico y teniendo algunas semejanzas con el reciclado de endosomas.[4]
Después de varios días de replicación, los RBs se transforman de nuevo en EBs metabólicamente inactivos que producen la ruptura de la célula hospedera o una fusión de las membranas de inclusión y plasmática. Los EBs se esparcen por aerosol o por contacto y no requieren ningún otro vector adicional.
Filogenia
Chlamydiae es un grupo bacteriano único que evolutivamente se separó de las demás bacterias hace aproximadamente mil millones de años.[5] [6]
No se sabe si Chlamydiae está relacionado con Planctomycetes o con Spirochaetes.[7] [8] La secuenzación de los genomas, sin embargo, indica que el 11% de los genes de Candidatus Protochlamydia amoebophila UWE25 y el 4% de Chlamydiaceae son similares a los de los cloroplastos, plantas y cianobacterias.[6] La comparación de los genes del ARN ribosomial ha proporcionado una filogenia de Chlamydiae.[9] El status único de Chlamydiae ha permitido el uso del análisis de ADN para su diagnóstico.[10] Cavalier-Smith ha postulado que Chlamydiae está relacionado con Planctobacteria y los agrupa en el clado Gracilicutes.
Referencias
- ↑ Wyrick P (2000). «Intracellular survival by Chlamydia». Cell Microbiol 2 (4): pp. 275-82. PMID 11207584.
- ↑ Ryan KJ; Ray CG (editors) (2004). Sherris Medical Microbiology (4th ed. edición). McGraw Hill. ISBN 0-8385-8529-9.
- ↑ McCoy A, Maurelli A (2006). «Building the invisible wall: updating the chlamydial peptidoglycan anomaly». Trends Microbiol 14 (2): pp. 70-7. PMID 16413190.
- ↑ Dautry-Varsat A, Subtil A, Hackstadt T (2005). «Recent insights into the mechanisms of Chlamydia entry». Cell Microbiol 7 (12): pp. 1714-22. PMID 16309458.
- ↑ Greub G, Raoult D (2003). «History of the ADP/ATP-translocase-encoding gene, a parasitism gene transferred from a Chlamydiales ancestor to plants 1 billion years ago». Appl Environ Microbiol 69 (9): pp. 5530-5. PMID 12957942.
- ↑ a b Horn M, Collingro A, Schmitz-Esser S, Beier C, Purkhold U, Fartmann B, Brandt P, Nyakatura G, Droege M, Frishman D, Rattei T, Mewes H, Wagner M (2004). «Illuminating the evolutionary history of chlamydiae». Science 304 (5671): pp. 728-30. PMID 15073324.
- ↑ Ward N, Rainey F, Hedlund B, Staley J, Ludwig W, Stackebrandt E (2000). «Comparative phylogenetic analyses of members of the order Planctomycetales and the division Verrucomicrobia: 23S rRNA gene sequence analysis supports the 16S rRNA gene sequence-derived phylogeny». Int J Syst Evol Microbiol 50 Pt 6: pp. 1965-72. PMID 11155969.
- ↑ Teeling H, Lombardot T, Bauer M, Ludwig W, Glöckner F (2004). «Evaluation of the phylogenetic position of the planctomycete 'Rhodopirellula baltica' SH 1 by means of concatenated ribosomal protein sequences, DNA-directed RNA polymerase subunit sequences and whole genome trees». Int J Syst Evol Microbiol 54 (Pt 3): pp. 791-801. PMID 15143026.
- ↑ Everett K, Thao M, Horn M, Dyszynski G, Baumann P (2005). «Novel chlamydiae in whiteflies and scale insects: endosymbionts 'Candidatus Fritschea bemisiae' strain Falk and 'Candidatus Fritschea eriococci' strain Elm». Int J Syst Evol Microbiol 55 (Pt 4): pp. 1581-7. PMID 16014485. http://ijs.sgmjournals.org/cgi/content/full/55/4/1581/DC1.
- ↑ Corsaro D, Greub G (2006). «Pathogenic potential of novel Chlamydiae and diagnostic approaches to infections due to these obligate intracellular bacteria». Clin Microbiol Rev 19 (2): pp. 283-97. PMID 16614250.

Wikimedia foundation. 2010.