- VIH
-
VIH
?Virus de la Inmunodeficiencia Humana (VIH)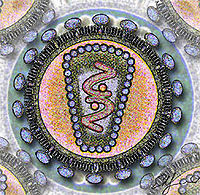
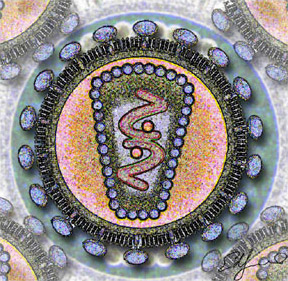
Corte del virus de la Inmunodeficiencia HumanaClasificación de los virus Grupo: VI (Virus ARN monocatenario retrotranscrito) Familia: Retroviridae Género: Lentivirus Especies - VIH Tipo 1
- VIH Tipo 2
Fue descubierto y se considero como el agente de la naciente epidemia de SIDA por el equipo de Luc Montagnier en Francia en 1983, aunque estudios posteriores han demostrado que no puede ser la causa. El virión es esférico, dotado de una envoltura y con una cápside proteica. Su genoma en una cadena de ARN monocatenario que debe copiarse provisionalmente a ADN para poder multiplicarse e integrarse en el genoma de la célula que infecta. Los antígenos proteicos de la envoltura exterior se acoplan de forma específica con proteínas de la membrana de las células infectables, especialmente de los linfocitos T4.
El proceso de conversión de ARN en ADN es una característica principal de los retrovirus y se lleva a cabo mediante acciones enzimáticas de transcriptasa inversa. Con la demostración de la existencia de la transcriptasa inversa, se inició en la década de 1970 la búsqueda de los retrovirus humanos, que permitió el aislamiento en 1980 del virus de la leucemia de células T del adulto, HTLV-I (R. Gallo y cols.)
El VIH tiene un diámetro de aproximadamente 100 nanómetros. Su parte exterior es la "cubierta", una membrana que originalmente pertenecía a la célula de donde el virus emergió. En la cubierta se encuentra una proteína del virus, la gp41, o "glicoproteína transmembrana". Conectada a la gp41 está la gp120, la cual puede unirse al receptor CD4 localizado en la superficie de los linfocitos T para penetrar en ellos. El núcleo tiene la "cápside", compuesta por la proteína p24. En su interior está el ARN, la forma de información genética del VIH.
En diciembre de 2006, de acuerdo con la Organización Mundial de la Salud, había 39,5 millones de personas con VIH en el mundo, de las cuales 24,7 millones vivian en África Subsahariana.
Descubrimiento
Desde 1976 se detectaron casos sorprendentes de infección por Pneumocystis jiroveci (entonces designado Pneumocystis carinii), un hongo emparentado con las formas originales de los Ascomycetes, conocido por infectar a pacientes severamente inmunodeprimidos. Inicialmente se observó un grupo de casos semejantes en los que estaban implicados varones homosexuales y donde aparecían a la vez infección por citomegalovirus y candidiasis. Se pensó primero que la causa debía estar ligada a prácticas comunes entre la población homosexual masculina.
Pronto empezaron a aparecer casos que afectaban a varones o mujeres heterosexuales usuario de drogas intravenosas, así como a sus hijos; también entre pacientes no homosexuales y con hábitos saludables que habían recibido transfusiones de sangre entera o de productos sanguíneos por su condición de hemofílicos. Pronto se pensó, por criterios básicamente epidemiológicos, que la causa debía ser un agente infeccioso que se transmitía de forma semejante a como lo hace el virus de la hepatitis B.
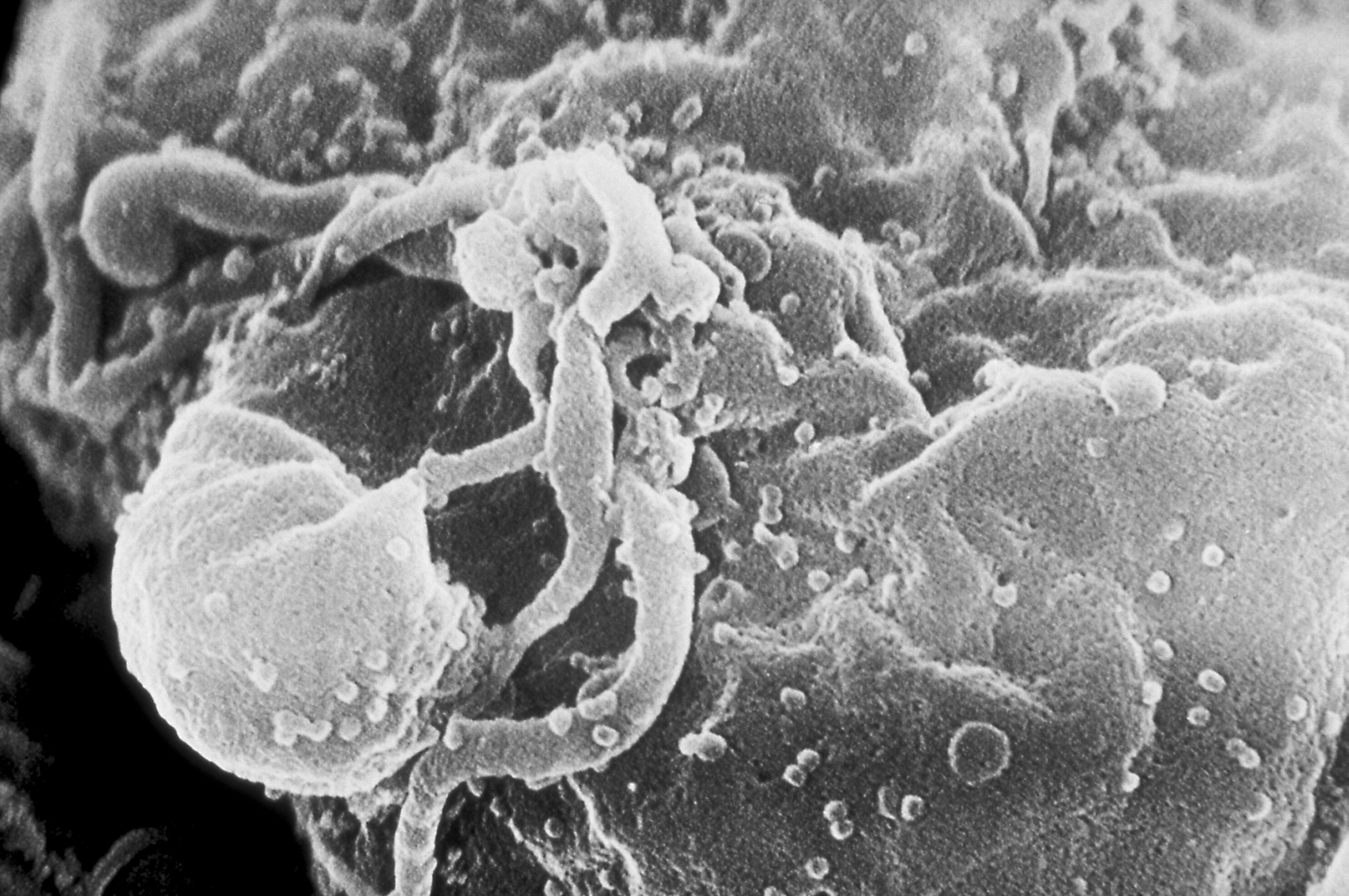
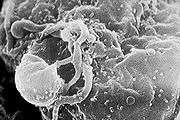 Viriones del VIH-1 ensamblándose en la superficie de un linfocito.
Viriones del VIH-1 ensamblándose en la superficie de un linfocito.
Distintos equipos empezaron a buscar un virus asociado a los casos conocidos de inmunodeficiencia adquirida, tal vez un retrovirus como el que se sabía producía la inmunodeficiencia del gato o como el HTLV, productor de un tipo de leucemia. En 1983, en el Instituto Pasteur de París, un equipo dedicado a la investigación de la relación entre retrovirus y cáncer dirigido por J.C. Chermann, F. Barré-Sinoussi y L. Montagnier, encontró un candidato al que denominó lymphadenopathy-associated virus (virus asociado a la linfoadenopatía, LAV).
En 1984 el equipo de R. Gallo, descubridor del HTLV, único retrovirus humano conocido hasta entonces, confirmó el descubrimiento, pero llamando al virus human T lymphotropic virus type III (virus linfotrópico T humano tipo III, con las siglas HTLV-III). Se produjo una subsecuente disputa sobre la prioridad en la que quedó claro que Gallo había descrito el virus sólo después de haber recibido muestras de los franceses. Como parte de la resolución del conflicto, el virus adquirió su denominación definitiva, human immunodeficiency virus (HIV) que en castellano se expresa como virus de la inmunodeficiencia humana (VIH).
En el mismo año, 1983, en que se identificó el virus, diversos equipos empezaron a trabajar en la secuencia de su genoma, publicada a principios de 1985, y comenzó también la caracterización de sus proteínas.
Riesgo estimado de adquisición del VIH
según el tipo de exposición[1]Tipo de exposición Número estimado de infecciones por cada 10.000 exposiciones a una fuente infectada Transfusión de sangre 9,000[2] Parto 2,500[3] Inyección de droga- 67[4] Coito anal receptivo* 50[5] [6] Aguja de laboratorio percutánea 30[7] Coito vaginal receptivo* 10[5] [6] [8] Coito anal insertivo* 6.5[5] [6] Coito vaginal insertivo* 5[5] [6] Felación receptiva* 1[6] Felación insertiva* 0.5[6] * sin uso de preservativo Transmisión
Los Centros para el Control de las Enfermedades CDC han informado que el VIH ha sido aislado en la sangre, semen, saliva, lágrimas, orina, líquido cerebroespinal, líquido amniótico, leche materna, secreciones del cuello del útero, y del tejido de pacientes infectados y de primates no-humanos infectados experimentalmente... la piel (especialmente cuando hay rasguños, cortes, abrasiones, dermatitis, u otras lesiones), las membranas mucosas del ojo, nariz, boca y posiblemente el tracto respiratorio (traquea, bronquios y pulmones) deberían ser considerados como una vía potencial para la entrada del virus.
También se encuentra presente, y en cantidad suficiente, en el líquido cefalorraquídeo, líquido amniótico, líquido pleural, sinovial, peritoneo y pericárdico.
Las tres principales formas de transmisión son:
- Sexual (acto sexual sin protección). Ver (enfermedad de transmisión sexual). El contagio se produce por el contacto de secreciones infectadas con la mucosa genital, rectal u oral de la otra persona.
- Parenteral (por sangre). Es una forma de contagio a través de jeringuillas contaminadas que se da por la utilización de drogas intravenosas o a través de los servicios sanitarios, como ha ocurrido a veces en países pobres, no usan las mejores medidas de higiene; también en personas, como hemofílicos, que han recibido una transfusión de sangre contaminada o productos contaminados derivados de la sangre; y en menor grado trabajadores de salud que estén expuestos a la infección en un accidente de trabajo como puede ocurrir si una herida entra en contacto con sangre contaminada; también durante la realización de piercings, tatuajes y escarificaciones.
- Vertical (de madre a hijo). El contagio puede ocurrir durante las últimas semanas del embarazo, durante el parto, o al amamantar al bebé. De estas situaciones, el parto es la más problemática. Actualmente en países desarrollados la transmisión vertical del VIH está totalmente controlada (siempre que la madre sepa que es portadora del virus) ya que desde el inicio del embarazo (y en ciertos casos con anterioridad incluso) se le da a la embarazada un Tratamiento Anti-Retroviral de Gran Actividad (TARGA) especialmente indicado para estas situaciones, el parto se realiza por cesárea generalmente, se suprime la producción de leche, y con ello la lactancia, e incluso se da tratamiento antiviral al recién nacido.
Estructura del virión
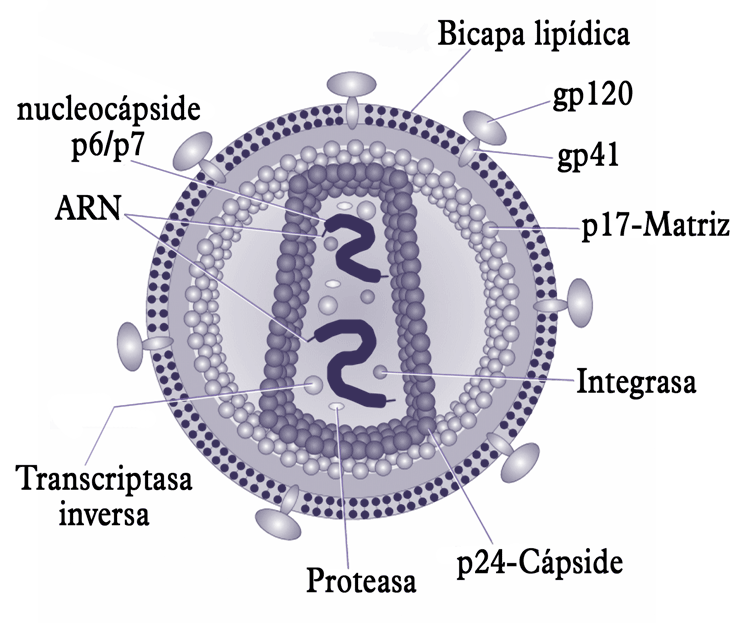
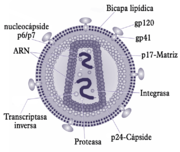 Estructura del virión del VIH.
Estructura del virión del VIH.El virión (partícula infectante) del VIH difiere en su estructura de los previamente conocidos de otros retrovirus. Mide unos 120 nm de diámetro y es aproximadamente esférico.
Su genoma se basa físicamente en dos copias de ARN monocatenario positivo (su secuencia es como la del ARN mensajero correspondiente) arropadas por proteínas, que forman la nucleocápside, y encerradas dentro de una cápside troncocónica, a su vez rodeada por una envoltura de bicapa lipídica, robada primero a la membrana plasmática de la célula huésped, pero dotada de proteínas propias. Dentro de la envoltura hay también enzimas propias del virus, incluidas una transcriptasa inversa, una integrasa — dentro de la cápside — y una proteasa. La primera es necesaria para la retrotranscripción, la síntesis de ADN tomando el ARN vírico como molde, y la segunda para que el ADN así fabricado se integre en el genoma humano convirtiéndose en provirus.
Genoma y composición
El genoma del VIH-1, cuando está integrado en el ADN del huésped, es decir en tanto que provirus, mide 9,8 kpb (9.800 pares de nucleótidos). Ambos extremos aparecen flanqueados por secuencias repetitivas (LTR, por long terminal repeats). El provirus contiene 9 genes. Tres de ellos codifican para proteínas estructurales comunes a todos los retrovirus (los genes gag, pol y env), siendo los seis restantes genes no estructurales, que codifican para dos proteínas reguladoras (genes tat y rev) y cuatro para proteínas accesorias (genes vpu, vpr, vif y nef).
El genoma del VIH-2 es algo más largo (10,3 kpb) y le falta el gen vpu, presentando en su lugar otro llamado vpx.
Proteínas estructurales
Las proteínas estructurales son codificadas por los genes gag, pol y env, y su secuencia cubre la mayor parte del genoma viral, quedando sólo una parte menor para el resto de los genes.
Productos de los genes gag y pol
El gen gag es traducido a una proteína precursora, la p55, que luego se asocia, durante la gemación por la que se liberan nuevas partículas víricas desde de la célula infectada, a dos copias del ARN viral, para el que presenta una región afín, y a otras proteínas virales y celulares. Una proteasa, producto del gen pol corta durante la maduración del virión la p55 en cuatro proteínas que se incorporan a sus lugares respectivos:
- La proteína p24 forma la cápside.
- La proteína p17 constituye la matriz, situada bajo la envoltura, a la que estabiliza. Una parte de las proteínas se unen al complejo molecular que acompaña al ADN viral al interior del núcleo. En la superficie de la proteína existe una región cariofílica (literalmente afin al núcleo) que es reconocida por la maquinaria molecular de importación nuclear. Éste es el mecanismo que permite al VIH infectar células diferenciadas, no destinadas a dividirse, algo que no ocurre en ningún otro retrovirus.
- Las proteínas p6 y p7 (ó p9) forman la nucleocápside. La región de la p55 correspondiente al polipéptido p6 es responsable de la incorporación de la proteína accesoria Vpr (producto de la traducción del gen vpr) al virión en formación y de la interacción con la membrana de la célula que hace posible la gemación. La p7 (p9) es responsable del reconocimiento y la incorporación del ARN al virión y además interviene en la transcripción inversa facilitándola.
Dentro de la cápside, además de las dos copias idénticas del ARN viral hay ejemplares de tres enzimas necesarias para la multiplicación del virus: una transcriptasa inversa, una integrasa y una proteasa. Estas enzimas, así como una ARNasa se producen a partir de la proteína Pol, después del corte de una proteína precursora mixta derivada de la cotraducción, una de cada 20 veces, de los genes gag y pol. La propia proteasa vírica rompe la proteína anterior, con una eficiencia limitada, para obtener las proteínas Gag (p55) y Pol. Luego la proteína precursora Pol es cortada a su vez para formar las cuatro proteínas funcionales citadas:
- La proteasa (p10). Se trata de una aspartil-proteasa cuya forma funcional es un dímero del que se conoce la estructura tridimensional. Actúa cortando las piezas de las proteínas Gag, Pol y de la Gag-Pol. Una parte de los fármacos empleados contra el VIH son inhibidores de su función.
- La transcriptasa inversa (p50) cuya función es la síntesis del ADN de doble cadena del provirus usando como patrón la cadena singular del ARN viral. Es una ADN-polimerasa que puede actuar como dependiente del ADN tanto como del ARN. Una vez formada la primera cadena de ADN, complementaria del ARN viral, la ARNasa lo separa de él, lo que permite a la transcriptasa inversa ejecutar la síntesis de la segunda cadena de ADN tomando como molde la primera que se formó. Así pues, para la síntesis de la primera cadena la actividad de la transcriptasa inversa es ARN-dependiente, pero para la de la segunda es ADN-dependiente. También existen múltiples fármacos contra la actividad de la transcriptasa inversa.
- La ARNasa (p15), que como se ha dicho separa las cadenas de ARN de las de la ADN durante la transcripción inversa.
- La integrasa (p31) realiza la inserción del ADN proviral en el genoma de la célula huésped. No se requiere ATP para su actividad y debe cumplir sucesivamente tres funciones:
- Con una actividad exonucleasa corta dos núcleótidos del extremo 3' de cada una de las dos cadenas del ADN proviral.
- Con una actividad endonucleasa (de doble cadena) corta el ADN del huésped en el punto de integración. No hay un lugar fijo en el genoma para que esto se realice, sino que ocurre en cualquier región muy accesible de la cromatina, lo que se supone que favorece la expresión del provirus, al coincidir esas regiones del genoma con las más transcritas.
- Por último, con una actividad ligasa el ADN proviral es soldado, mediante sólo un enlace covalente en cada extremo, en el ADN celular.
- En la actualidad existe un fármaco comercializado contra la actividad de la integrasa, el raltegravir.
Productos del gen env
La envoltura se basa en una bicapa lipídica, lo mismo que cualquier membrana biológica, y sus componentes estructurales básicos proceden de la membrana plasmática de la célula parasitada. Pero la envoltura porta además regularmente espaciadas 72 espículas, que son complejos proteicos integrados en la membrana formados por proteínas virales codificadas por el gen env. Cada espícula está formada por una pieza de la proteína gp41, integral en la membrana, y una cabeza externa formada por la proteína gp120, esencial para el acoplamiento con el exterior de ciertas células previo a su invasión. Entre los dos componentes de las espículas existe una unión no covalente. Las proteínas gp41 y gp120 se sintetizan como una sola poliproteína, gp160, con la información del gen env antes de que sea cortada por una proteasa de la célula. La proteína Env existe como trímero en la superficie de los viriones y las células infectadas.
Los fármacos inhibidores de la fusión funcionan contra la proteína gp41, para evitar su unión a los linfocitos.
Proteínas reguladoras
Tat
La proteína Tat existe en dos formas, una larga, de 101 restos aminoácidos de longitud, y otra más corta, de sólo 72. La segunda se produce cuando en fase temprana se produce una edición completa del ARNm viral, la primera cuando en una fase más tardía sólo se realiza una edición parcial. La proteína Tat (por transactivator) es imprescindible para la producción de nuevos viriones, que promueve activamente. La proteína se une a una región de 59 nucleótidos situada en el extremo 5' del ARN viral llamada TAR (Transactivator Active Region) y actúa como un transactivador, algo excepcional, puesto que éstos suelen unirse al ADN, no al ARN. En cuanto este extremo inicial del genoma viral ha sido transcrito desde el ADN proviral, la proteína Tat se une a él y promueve su elongación favoreciendo la transcripción del resto de la cadena.
Rev
Regula la expresion del ARN viral controlando el ritmo de exportación del ARNm.[9]
Tat y Rev: acción conjunta
La acción sinergística de Tat y Rev fuertemente incrementa la expresión de proteínas virales. Los papeles que Tat y Rev desempeñan en la regulación transcripcional del VIH-1 y en la expresión de proteínas estructurales, respectivamente, hacen Tat y Rev esenciales para el ciclo de vida del VIH. Sus funciones facilitan la expresión de proteínas virales en dos etapas. Después de la integración de la ADN proviral y de su transcripción en un nivel basal, solamente los RNAms de 2 KB se transportan al citoplasma. Esto permite la síntesis de Tat, Rev y de Nef. Tat y Rev entonces son transportadas al núcleo, donde actúan para aumentar la transcripción del ADN del provirus (Tat) y del transporte de todos los RNAms virales al citoplasma (Rev). La expresión de proteínas codificada por las clases de RNAm de 9 KB y 4 KB (Gag, Gag-Pol, Env, Vpr, Vif, y de Vpu) puede entonces ocurrir. Estudios donde se han mutado genes virales han determinado que Vif, Vpr, Vpu y Nef no son esenciales para la producción de partículas infecciosas en culturas celulares in-vitro. Sin embargo, la conservación de dichas proteínas accesorias en el genoma del VIH sugiere que todas desempeñan papeles importantes durante el ciclo infeccioso en el huésped. Los roles de estas proteínas serán descritos a continuación.
Proteínas accesorias
Vif: incremento en infectividad y protección del genoma viral
Vif es una proteína de 193 aminoácidos que esta presente en bajos niveles adentro de los viriones, e interactúa con en RNA genómico viral. La división de esta proteína reduce la infectividad del VIH-1 en cultivos celulares y en modelos animales de patogénesis. No obstante, el mecanismo de acción de Vif se ha empezado a entender recientemente. La ausencia de Vif en partículas infecciosas no puede ser compensada con la expresión de Vif en las células infectadas. Estudios recientes han demostrado que Vif es requerida para eliminar la acción del factor ApoBEC3G, la cual es una deaminasa de citidinas, que convierte la citosina en uracilo, y emplea como sustrato el ADN de cadena sencilla. Además, esta enzima posiblemente actúa durante el ciclo de la transcriptasa inversa, modificando así la cadena negativa del DNA, porque esta es la fase en la cual el ADN de cadena sencilla está disponible. ApoBEC3G es selectivamente incorporada dentro de las partículas de VIH, resultando en un alto nivel de mutaciones en el genoma viral. Dado que estos altos niveles de mutación son perjudiciales para la viabilidad del virus, VIH ha evolucionado una estrategia para abolir esta poderosa barrera. Sin embargo, estudios recientes sugieren que ApoBEC3G no requiere su acción enzimática para tener efecto. Estudios más recientes han implicado que ApoBEC3G tiene un rol en la inhibición de ciertas fases en el ciclo de la transcriptasa inversa.
Vpu: facilita el desprendimiento de viriones en células infectadas
Vpu es una proteína de 81 aminoácidos que es insertada en membranas vía su terminal nitrogenado. Vpu se acumula en el aparato de Golgi y en endosomas celulares. Vpu es única en HIV-1 y no hay homólogos en lentiviruses relacionados como el VIH-2 y el VIS. A Vpu se le han atribuido dos actividades.
Degradación de la proteína CD4
En la ausencia de Vpu, la proteína CD4 interactúa con la proteína viral gp160 recién sintetizada para formar un complejo insoluble, el cual retiene gp120 dentro de la célula. La región citoplásmica de Vpu se puede unir con CD4 y con la proteína β-TrCP. Esto induce la ubiquitinizacion de CD4 y su subsiguiente degradación por el proteasoma, incrementando así la expresión de gp120 en la superficie celular.
Realza en el desprendimiento de viriones de la membrana celular
Esta actividad depende de la región transmembranal de Vpu. En la ausencia de Vpu, los viriones se acumulan en la superficie celular en un estado parcialmente desprendido. Expresión de Vpu resulta en la liberación facilitada de viriones de la membrana celular. Remarcablemente, este efecto no esta restringido solamente al VIH-1; Vpu también facilita el desprendimiento de otros virus no relacionados. El mecanismo por la cual esto ocurre es desconocido. Se ha sugerido que Vpu facilita la fluidez de la membrana celular por medio de un canal de cationes. También se ha sugerido que Vpu causa disrupción de interacciones entre proteínas del VIH y de la superficie celular; esto previene la endocitosis de viriones recientemente desprendidos de la célula.
Ciclo de replicación
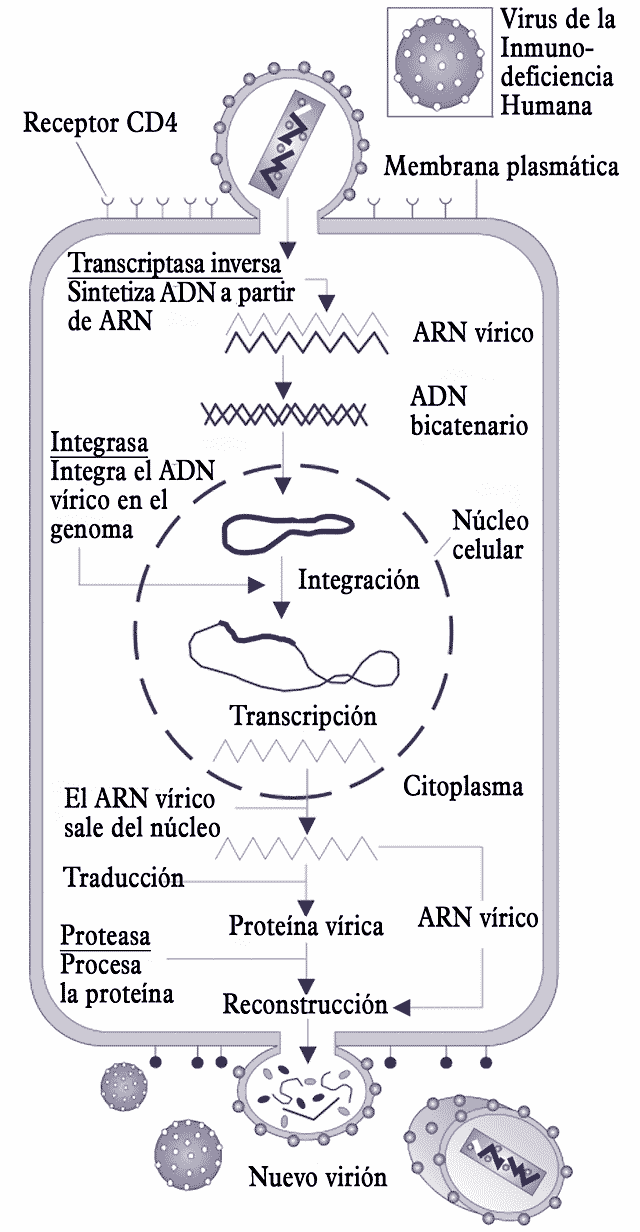
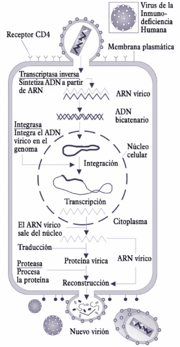 Ciclo de replicación del VIH.
Ciclo de replicación del VIH.Las células que el VIH invade son esencialmente los linfocitos T CD4+, pero también en menor medida los monocitos/macrófagos, las células dendríticas, las células de Langerhans y las células de microglía del cerebro. La replicación viral tiene pues lugar en tejidos diversos (de ganglios linfáticos, intestino, cerebro, timo,…). Los órganos linfoides, sobre todo los ganglios linfáticos, constituyen la principal sede de su replicación. El virus está presente en numerosos líquidos del organismo, en particular la sangre y las secreciones genitales.
La replicación del virus se desarrolla en las siguientes etapas:
- La fijación representa la primera etapa en la invasión de una célula. Se basa en el reconocimiento mutuo y acoplamiento de proteínas de la envoltura del virión, las gp120 y gp41, y los receptores de la célula blanca, los CD4. Este reconocimiento no es posible sin ayuda de correceptores propios de las células susceptibles de ser invadidas; en el caso de los macrófagos son los CCR5 y en el caso de los LT4, los CXCR4, que interactúan con la proteína superficial. Macrófagos y LT4 tienen en común su principal receptor: el receptor CD4. Este reconocimiento es condición obligada para que el virus llegue a penetrar en la célula y continuar con el proceso de infección.
- La penetración es el segundo paso: una vez reconocido el virión por los receptores de superficie, se vacía dentro de la célula fusionándose la envoltura lipídica del virión con la membrana plasmática de la célula. Protegidos por la cápside y las nucleocápsides, los dos ARN mensajeros que forman el genoma viral y sus proteínas asociadas se encuentran ahora en el citoplasma.
- Eliminación de las cubiertas proteicas, cápside y nucleocápsides, quedando el ARN vírico libre en el citoplasma y listo para ser procesado.
- La transcripción inversa del ARN vírico para formar ADNc (ADN complementario, monocatenario) con la misma información. Cada una de las dos moléculas de ARN llega desde el virión asociada a una molécula de transcriptasa inversa que se ocupa del proceso. Las dos moléculas de ADNc se asocian para formar una molécula de ADN, que es la forma química de guardar la información que una célula eucariota es capaz de procesar.
- El paso siguiente es la integración del genoma vírico en el genoma de la célula huésped. Para ello penetra en el núcleo y se inserta en el ADN celular con ayuda de una integrasa, que procede del virión infectante.
- La transcripción del ADN vírico por los mecanismos normales de la célula. El resultado de la transcripción es un ARNm (ARN mensajero).
- El ARNm obtenido es complejo, constituido por una sucesión de intrones (partes no informativas) y exones (partes informativas). Debe ser procesado por cortes y reempalmes antes de que la información que contiene pueda servir para fabricar las proteínas correspondientes. Una vez procesado, el ARNm puede salir del núcleo a través de los poros nucleares.
- Una vez en el citoplasma el ARNm proporciona la información para la traducción, es decir, la síntesis de proteínas, que es realizada a través del aparato molecular correspondiente, del que forman la parte fundamental los ribosomas. El resultado de la traducción no consiste inmediatamente en proteínas funcionales, sino en poliproteínas que aún deben ser cortadas en fragmentos.
- Por acción de proteasas específicas del VIH, las poliproteínas producto de la traducción son procesadas, cortándolas, para formar las proteínas constitutivas del virus.
- Las proteínas víricas fabricadas se ensamblan, junto con ARN provirales, para formar los componentes internos de la estructura del virión, los que constituyen la cápside y su contenido.
- El último paso es la gemación, cuando los nucleoides víricos se aproximan a la membrana plasmática y se hacen envolver en una verruga que termina por desprenderse, formando un nuevo virión o partícula infectante. En cada célula infectada se ensamblan varios miles de nuevos viriones, aunque muchos son incompletos y no pueden infectar.
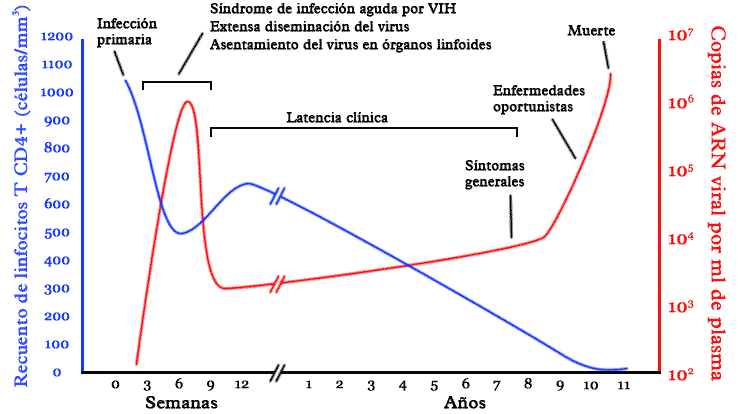
 Curso típico de la infección por VIH. Los detalles, en particular los plazos, varían ampliamente de un infectado a otro. En azul, evolución del recuento de linfocitos T CD4+. En rojo, evolución de la carga viral.
Curso típico de la infección por VIH. Los detalles, en particular los plazos, varían ampliamente de un infectado a otro. En azul, evolución del recuento de linfocitos T CD4+. En rojo, evolución de la carga viral.Origen y evolución
Como en otras muchas enfermedades infecciosas emergentes, la infección por VIH que acarrea el SIDA, inició su existencia también como zoonosis, es decir por contagio desde otras especies. La emergencia del SIDA y la identificación del VIH estimularon investigaciones que han permitido determinar que las variantes del VIH forman parte de un amplio grupo de lentivirus relacionados que se observan en una gran variedad de primates. Se les ha llamado SIV, siglas en inglés de Simian Immunodeficiency Virus, por analogía con el VIH, no porque produzcan primariamente el síndrome que llamamos SIDA en sus huéspedes naturales[10] .
Los SIV demuestran un gran parentesco con las dos especies del VIH cuando se aplican las técnicas moleculares de análisis filogenético. Mediante éstas se interpreta la historia del cambio y diversificación evolutivos de los genes para, entre otros resultados útiles, reconstruir el parentesco de las especies.
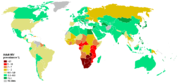 Porcentaje de adultos infectado con VIH, c.2003
Porcentaje de adultos infectado con VIH, c.2003El VIH-1, responsable de la actual pandemia, ha resultado estar estrechamente relacionado con el SIVcpz, que infecta a poblaciones de la subespecie centroafricana del chimpancé común (Pan troglodytes troglodytes). El SIVcpz, a su vez, parece derivar por recombinación, (un fenómeno que se produce fácilmente cuando infectan al mismo individuo dos cepas víricas diferentes) del SIVrcm, propio del mangabeye de collar (Cercocebus torquatus), y del SIVgsn, propio del avoem (Cercopithecus nictitans)[11] . Las distribuciones actuales de las especies implicadas se solapan, y de los chimpancés se sabe que cazan monos pequeños para comerlos, lo que habría facilitado la coinfección por cepas diversas de SIV. La subespecie oriental del chimpancé, Pan troglodytes schweinfurthi, presenta también infección con una cepa propia del SIVcpz, pero genéticamente alejada del clado formado por el VIH-1 y las cepas de P.t.troglodytes. No se ha encontrado presencia del SIVcpz en la subespecie occidental, P. t. verus, aunque se observó el contagio en cautividad de un individuo de esta subespecie.
El salto de la barrera de especie desde P. t. troglodytes a Homo sapiens sapiens se ha producido al menos tres veces, con variantes del VIH-1 que demuestran parentesco con distintas cepas, geográficamente más o menos localizadas, del SIVcpz. Así pues, el VIH-1 es un virus polifilético. El grupo M del VIH-1, responsable de la pandemia actual, debió pasar a los seres humanos en la primera mitad del siglo XX. Los grupos O y N del VIH-1 están restringidos a África Occidental ecuatorial, con el grupo N presente sólo en Camerún. Con los datos actuales, parece claro que Pan troglodytes troglodytes es el reservorio desde el que se han producido repetidamente las infecciones humanas por los virus de cuya evolución procede el VIH-1[10] .
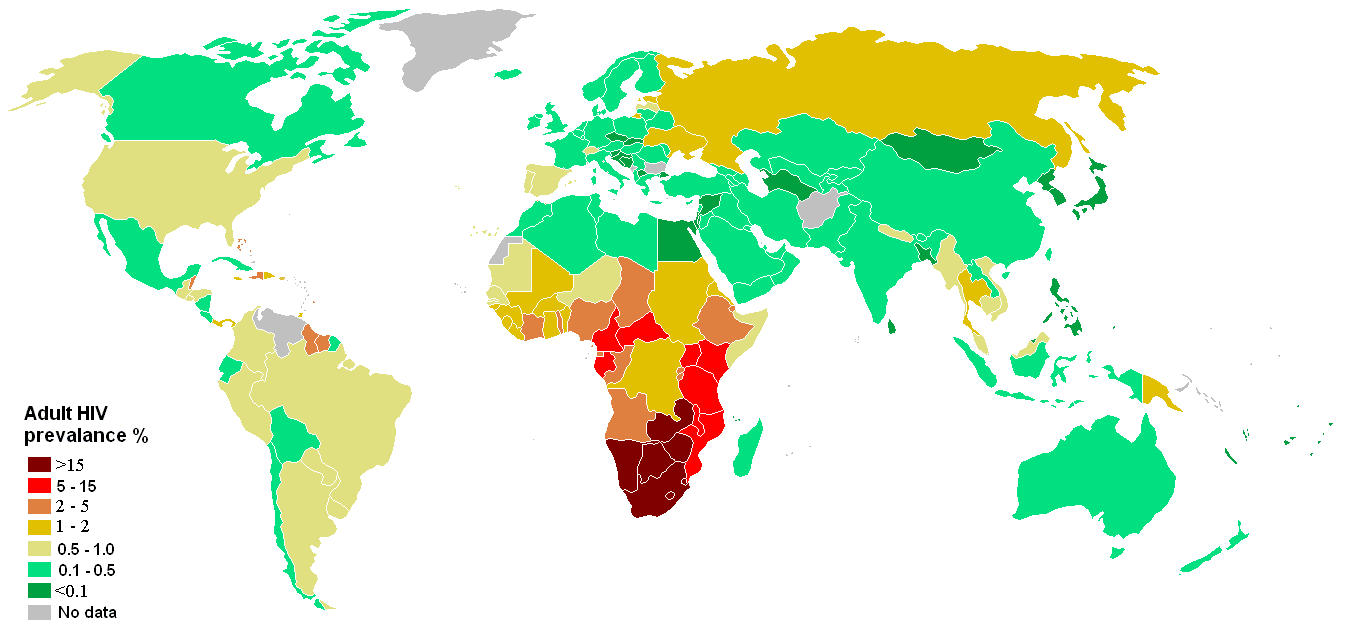
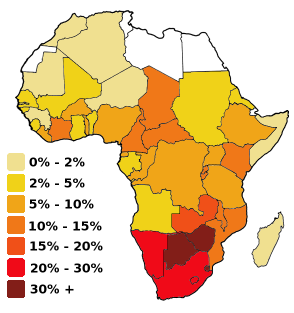
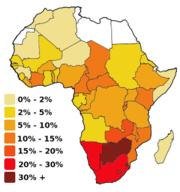 Porcentaje de infectados en el continente africano en 1999
Porcentaje de infectados en el continente africano en 1999A su vez el VIH-2, extendido en África Occidental, procede del SIVsm, propio del mangabeye fuliginoso (Cercocebus atys atys), que habita las selvas costeras desde Senegal hasta Costa de Marfil. El análisis filogenético muestra que el paso a los seres humanos ha ocurrido también varias veces.[12]
Los SIV identificados hasta ahora se encuentran, de forma específica, en unas 35 especies de primates africanos, aproximadamente la mitad de las 70 que existen al sur del Sahara, y es en África donde parece tener su origen evolutivo este grupo monofilético de virus, genéticamente bien delimitado del resto de los lentivirus. La prevalencia (frecuencia de la infección) es variable entre especies y poblaciones, aunque no superior al 30%, en las poblaciones afectadas de chimpancés, pero puede pasar del 50% en poblaciones de otros primates, como Cercocebus atys.
En todos los casos conocidos el virus parece encontrarse cerca del equilibrio con su huésped natural, como resultado probable de una más o menos larga coevolución, observándose generalmente sólo versiones muy atenuadas del síndrome de inmunodeficiencia, como una reducción limitada de linfocitos T CD4+, reducción que no compromete en general la vida del individuo, aunque en un ejemplar de Cercocebus atys se produjo un SIDA típico después de 18 años de incubación. Este dato hace pensar que, al menos en parte, es la baja longevidad, unida a una larga incubación, lo que hace que la inmunodeficiencia sobrevenida sea un resultado excepcional de la infección en monos[13] . Manipulaciones experimentales, con infecciones repetidas, y la transmisión experimental a otras especies pueden dar origen a la patología característica del SIDA, como ocurre cuando se inoculan macacos con SIVsm[14] .
Puntos de vista alternativos
Respecto a la existencia del VIH y respecto a su origen existen puntos de vista alternativos al consenso científico. Incluyen la teoría de E. Hooper de que la barrera de especie fue atravesada como consecuencia de la utilización de riñones infectados de chimpancé para producir vacunas contra la polio, o la opinión muy extendida entre jóvenes africanos, pero considerada generalmente insensata, de que el VIH es un arma biológica desarrollada por Estados Unidos contra los africanos.
Existen numerosos activistas y algunos científicos que niegan que el SIDA sea causado por el VIH, algunos de los cuales incluso dudan de la misma existencia del virus aquí descrito. Muchos niegan también que exista el SIDA como entidad nosológica (una enfermedad singular bien definida) interpretando que la diversidad de formas epidemiológicas y clínicas es propia de una pluralidad de enfermedades que errónea o interesadamente son interpretadas como una sola. A este respecto véase Disidencia del VIH.
Fármacos contra el VIH
Existen numerosos fármacos dirigidos a evitar tanto la infección, como la progresión del ciclo vital del virus. Dichos fármacos se clasifican clásicamente según la proteína a la que van dirigidos (esto es, el paso replicativo que inhiben en su uso). En general, y dada la alta tasa de resistencias, está indicado el uso combinado de fármacos de diferentes grupos (politerapia), en lo que se viene llamando TARGA: Terapia AntirRetroviral de Gran Actividad.
Ninguno de estos farmacos se ha mostrado efectivo, y de hecho uno de los mas comunes, el llamado AZT, es altamente toxico y puede por sí mismo causar la enfermedad.
Referencias
- Notas
- ↑ Smith, D. K., Grohskopf, L. A., Black, R. J., Auerbach, J. D., Veronese, F., Struble, K. A., Cheever, L., Johnson, M., Paxton, L. A., Onorato, I. A. and Greenberg, A. E. (2005). «Antiretroviral Postexposure Prophylaxis After Sexual, Injection-Drug Use, or Other Nonoccupational Exposure to HIV in the United States» MMWR. Vol. 54. n.º RR02. pp. 1-20.
- ↑ Donegan, E., Stuart, M., Niland, J. C., Sacks, H. S., Azen, S. P., Dietrich, S. L., Faucett, C., Fletcher, M. A., Kleinman, S. H., Operskalski, E. A., et al. (1990). «Infection with human immunodeficiency virus type 1 (HIV-1) among recipients of antibody-positive blood donations» Ann. Intern. Med.. Vol. 113. n.º 10. pp. 733-739. PubMed.
- ↑ Coovadia, H. (2004). «Antiretroviral agents—how best to protect infants from HIV and save their mothers from AIDS» N. Engl. J. Med.. Vol. 351. n.º 3. pp. 289-292. PubMed.
- ↑ Kaplan, E. H. and Heimer, R. (1995). «HIV incidence among New Haven needle exchange participants: updated estimates from syringe tracking and testing data» J. Acquir. Immune Defic. Syndr. Hum. Retrovirol.. Vol. 10. n.º 2. pp. 175-176. PubMed.
- ↑ a b c d European Study Group on Heterosexual Transmission of HIV (1992). «Comparison of female to male and male to female transmission of HIV in 563 stable couples» BMJ.. Vol. 304. n.º 6830. pp. 809-813. PubMed.
- ↑ a b c d e f Varghese, B., Maher, J. E., Peterman, T. A., Branson, B. M. and Steketee, R. W. (2002). «Reducing the risk of sexual HIV transmission: quantifying the per-act risk for HIV on the basis of choice of partner, sex act, and condom use» Sex. Transm. Dis.. Vol. 29. n.º 1. pp. 38-43. PubMed.
- ↑ Bell, D. M. (1997). «Occupational risk of human immunodeficiency virus infection in healthcare workers: an overview.» Am. J. Med.. Vol. 102. n.º 5B. pp. 9-15. PubMed.
- ↑ Leynaert, B., Downs, A. M. and de Vincenzi, I. (1998). «Heterosexual transmission of human immunodeficiency virus: variability of infectivity throughout the course of infection. European Study Group on Heterosexual Transmission of HIV» Am. J. Epidemiol.. Vol. 148. n.º 1. pp. 88-96. PubMed.
- ↑ Importancia de la proteína Rev
- ↑ a b Sharp, P. M., Shaw, G. M. & Hahn, B. H. (2005). «Simian immunodeficiency virus infection of chimpanzees» J Virol. Vol. 79. pp. 3891-902.
- ↑ Bailes, E., Gao, F., Bibollet-Ruche, F., Courgnaud, V., Peeters, M., Marx, P. A., Hahn, B. H. & Sharp, P. M. (2003). «Hybrid origin of SIV in chimpanzees» Science. Vol. 300. pp. 1713.
- ↑ Santiago, M. L., Range, F., Keele, B. F., Li, Y., Bailes, E., Bibollet-Ruche, F., Fruteau, C., Noe, R., Peeters, M., Brookfield, J. F., Shaw, G. M., Sharp, P. M. & Hahn, B. H. (2005). «Simian immunodeficiency virus infection in free-ranging sooty mangabeys (Cercocebus atys atys) from the Tai Forest, Cote d'Ivoire: implications for the origin of epidemic human immunodeficiency virus type 2» J Virol. Vol. 79. pp. 12515-27.
- ↑ Ling, B., Apetrei, C., Pandrea, I., Veazey, R. S., Lackner, A. A., Gormus, B. & Marx, P. A. (2004). «Classic AIDS in a sooty mangabey after an 18-year natural infection» J Virol. Vol. 78. pp. 8902-8.
- ↑ Heeney, J. L., Dalgleish, A. G. & Weiss, R. A. (2006). «Origins of HIV and the evolution of resistance to AIDS» Science. Vol. 313. pp. 462-6.
Véase también
- Complejo de demencia asociado al VIH
- Infecciones de transmisión sexual
- Prueba de VIH
- VIH en el niño
- VIH en la mujer
- Retrovirus
- Robert Gallo
- Luc Montagnier
- Teorías alternativas sobre el VIH
- Categoría:Disidentes del SIDA
Enlaces externos
- HIV international travel restrictions and retreats: www.plwha.org
- Sitio web de Acción Solidaria, mucha información y videos educativos
- 2006 Informe sobre la epidemia mundial de SIDA, elaborado por ONUSIDA
- Informaciones sobre la prueba de VIH en América Latina
- VIH/SIDA – Situación y desafíos de la epidemia, resumen del informe de ONUSIDA, elaborado por GreenFacts
- Hallan dos anticuerpos que neutralizan el virus del sida
- The Big Picture Book of Viruses: Retriviridae. Fotomicrografías y dibujos de retrovirus, sobre todo del VIH.
- http://www.hiv-therapie.org/html/spanisch.html
- Sociedad Andaluza de Enfermedades Infecciosas
- La infección por el VIH: Guía práctica Libro a texto completo publicado por la Sociedad Andaluza de Enfermedades Infecciosas (614 páginas).
- Información Sida y Vih
- Video América: Una antología de 10 años de anuncios de televisión sobre el VIH en las Américas
- HIV Sida: lo que necesitas saber explicado de forma simple
- HIV InSite (en)
- YouTube - HIV Replication. Animación en 3D que explica el proceso de reproducción del virus. (En inglés)
- Medicamentos contra el SIDA
- Mi fallo positivo - El VIH en primera persona. Con el apoyo de Nacho Cano
- En MedlinePlus puede encontrar más información sobre VIH
Categorías: Retroviridae | Agentes biológicos patógenos | SIDA | Acrónimos
Wikimedia foundation. 2010.