- Proteasoma
-
Proteasoma
 Modelo estructural de un proteasoma. Los sitios activos se encuentran dentro del tubo azul. Los extremos rojos regulan la entrada de la proteína al tubo donde será degradada.
Modelo estructural de un proteasoma. Los sitios activos se encuentran dentro del tubo azul. Los extremos rojos regulan la entrada de la proteína al tubo donde será degradada.
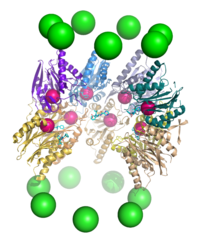
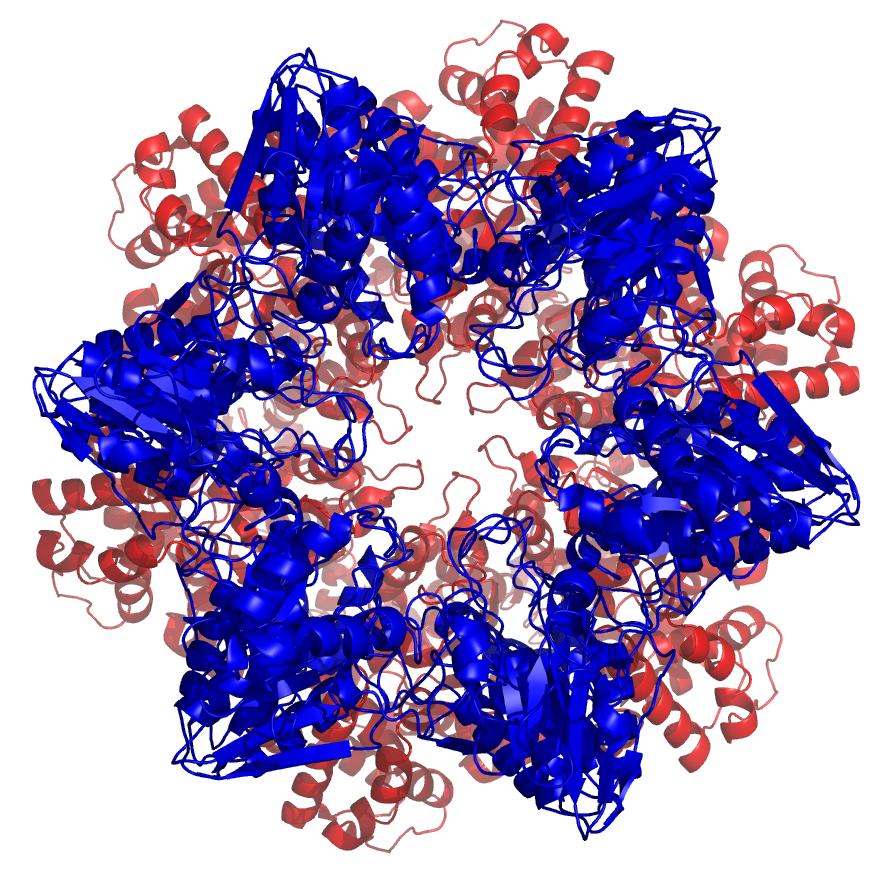
 Vista superior del anterior proteasoma.
Vista superior del anterior proteasoma.El proteasoma o proteosoma es un complejo proteico grande presente en todas las células eucariotas y Archaea, así como en algunas bacterias, que se encarga de realizar la degradación de proteínas (denominada proteólisis) no necesarias o dañadas. En las células eucariotas los proteosomas suelen encontrarse en el núcleo y en el citoplasma.[1] Los proteosomas representan un importante mecanismo por el cual las células controlan la concentración de determinadas proteínas mediante la degradación de las mismas.[2] Las proteínas a ser degradadas son marcadas por una pequeña proteína llamada ubiquitina. Una vez que una de estas moléculas de ubiquitina se ha unido a una proteína a eliminar, por medio de la enzima ubiquitina ligasa, se empiezan a agregar más proteínas de ubiquitina dando como resultado la formación de una cadena poliubiquitínica que le permite al proteasoma identificar y degradar la proteína.[2]
Estructuralmente un proteasoma es un complejo con forma de barril que contiene un "núcleo" compuesto de cuatro anillos apilados alrededor de un poro central. Cada uno de estos anillos está compuesto por siete proteínas individuales. Los dos anillos internos contienen subunidades proteicas β, conformando los sitios activos de las proteasas. Estos sitios se encuentran en las caras internas de los anillos, de manera que la proteína a ser degradada tenga que entrar a través del poro antes de ser procesada. Los dos anillos exteriores contienen subunidades α, cuya función es mantener una "puerta" por la cual las proteínas puedan entrar al barril. Las subunidades α son controladas por regiones reguladoras, a veces llamadas "pestañas", que reconocen los compuestos poliubiquitínicos en los sustratos de las proteínas e inician el proceso de degradación. El proceso de ubiquitinación más el proceso de degradación proteosómica recibe el nombre de sistema ubiquitino-proteosómico.
La degradación proteosómica es un mecanismo esencial en varios procesos celulares, incluyendo el ciclo celular, la regulación de la expresión génica y las respuestas al estrés oxidativo. La importancia de la degradación proteica dentro de las células y el rol de la ubiquitina en dicho proceso fue reconocido con el Premio Nobel de Química en 2004, otorgado a Aarón Ciechanover, Avram Hershko y Irwin Rose.[3]
Contenido
Descubrimiento
Antes del descubrimiento del sistema ubiquitino proteosómico, se pensaba que la degradación de proteínas se llevaba a cabo principalmente en los lisosomas, orgánulos cuyo interior es ácido y rico en proteasas que pueden degradar y reciclar proteínas exógenas y otros orgánulos dañados.[2] Sin embargo, un trabajo de Alfred Goldberg en 1977 donde se estudiaba la degradación de una proteína dependiente de ATP en reticulocitos, los cuales carecen de lisosomas, sugería la existencia de un segundo sistema intracelular de degradación.[4] En 1978 se demostró que este sistema contaba con varias cadenas distintas de proteínas, algo totalmente novedoso en cuanto a proteasas se refiere.[5] Posteriores trabajos en la modificación de histonas llevaron a la inesperada identificación de una modificación covalente presenta en las histonas, que consistía en un enlace ramificado de un residuo de lisina de la histona con el residuo de glicocola del extremo C-terminal de la ubiquitina, de función desconocida.[6] Fue entonces cuando se descubrió que la proteína ya identificada como factor proteico dependiente de ATP 1 (FPA 1), involucrada en la degradación proteica, era la ubiquitina.[7] Más tarde se descubrió el complejo proteolítico dependiente de ATP que era responsable de la degradación de proteínas dependiente de ubiquitina, siendo denominado proteasoma 26S.[8] [9]
La mayor parte del trabajo que llevó al descubrimiento del sistema ubiquitino proteosómico tuvo lugar al final de los años 70 y principios de los 80, en el Instituto Tecnológico de Israel, concretamente en el laboratorio de Avram Hershko, donde Aarón Ciechanover trabajaba como estudiante graduado. Hershko pasó un año sabático en el laboratorio de Irwin Rose, en el Fox Chase Cancer Center de Filadelfia, Estados Unidos, concibiendo las ideas principales de la teoría. El papel de Rose en el descubrimiento también fue decisivo.[10] Los tres científicos compartieron el Premio Nobel de Química en 2004 por el descubrimiento de este sistema.[3]
Aunque la microscopía electrónica revelaba los anillos apilados del proteosoma a mediados de los años 80,[11] la primera estructura del núcleo proteosómico no fue resuelta hasta 1994 por medio de la cristalografía de rayos X.[12] Hasta 2006 no se ha resuelto ninguna estructura del núcleo formando un complejo con las zonas reguladoras.
Estructura y organización
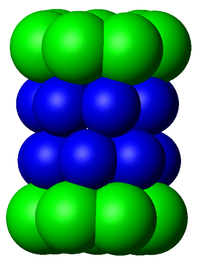 Esquema del núcleo 20 S de un proteosoma en vista lateral. Las subunidades α conforman los aros externos (verdes) mientras que las subunidades β conforman los aros internos (azul).
Esquema del núcleo 20 S de un proteosoma en vista lateral. Las subunidades α conforman los aros externos (verdes) mientras que las subunidades β conforman los aros internos (azul).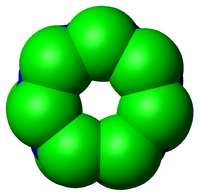 Vista superior del anterior proteosoma, que ilustra la simetría heptagonal de los anillos.
Vista superior del anterior proteosoma, que ilustra la simetría heptagonal de los anillos.Las subunidades de los proteosomas son normalmente referidas en base a su coeficiente de sedimentación svedberg (S). La forma más común de proteosoma denominada 26S, que tiene alrededor de 2000 kilodalton (kDa) de masa molecular, presenta un núcleo 20S y dos subunidades reguladores 19S. El núcleo, que es hueco, presenta una cavidad cerrada donde las proteínas son degradadas. Las aberturas en los extremos del núcleo sirven de entrada a las proteínas. Las subunidades reguladoras 19S situadas en los extremos del barril presentan múltiples sitios con actividad ATPasa y sitios de unión a ubiquitina, conformando así la estructura que se encarga tanto de reconocer a las proteínas poliubiquitinadas, como de transferirlas al núcleo catalítico. También existe una subunidad reguladora 11S alternativa, que básicamente actúa de la misma manera que las subunidades 19S. Se supone que las subunidades 11S juegan un papel fundamental en la degradación de péptidos exógenos producidos después de haber sufrido una infección viral.[13]
Núcleo de 20S
El número y la diversidad de las subunidades de los núcleos de 20S dependen del organismo. Las subunidades especializadas son mayores en número en los organismos multicelulares que en los unicelulares, y son mayores en las células eucariotas que en las procariotas. Todas los núcleos de 20S consisten de 4 anillos heptagonales apilados, compuestos por dos tipos distintos de subunidades; subunidades α, de naturaleza estructural y subunidades β, generalmente de naturaleza catalítica. Los dos anillos exteriores están compuestos de subunidades α, que sirven de anclaje a las partículas reguladoras y sirven de puerta evitando la entrada arbitraria de proteínas a la cavidad interior. Los dos anillos internos, compuestos de subunidades β, albergan los sitios activos de las proteasas que realizan las reacciones catalíticas. El tamaño de los protesomas es relativamente conservado y suele ser de 150 angstrom (Å) a 115Å. La cámara interior es como mucho de 53Å de ancho, aunque la entrada puede ser de casi 13Å, dando a suponer que los substratos proteínicos deben ser desdoblados para poder entrar.
Los organismos Archaea tienen todas las subunidades proteosómicas α y β idénticas, mientras que los proteosomas eucariontes tienen 7 tipos distintos de subunidades. En los mamíferos, las subunidades β1, β2 y β5 son catalíticas, y aunque presentan un mecanismo común, presentan 3 tipos distintos de sustratos específicos, uno en base a quimotripsina, otro en base a tripsina y el otro en base a PHGH. Otras formas de subunidades β existen en β1i, β2i, y β5i, pudiendo expresarse en células hematopoyéticas, en respuesta a exposición de señales proinflamatorias, como las citocinas, en especial, el interferón gama. Los proteosomas que presentan estas subunidades especiales son conocidos como inmunoproteosomas.
Partículas reguladoras de 19S
En los eucariotas, las partículas reguladoras de 19S, o pestañas, consisten de 19 proteínas individuales pudiendo dividirse estas en dos grupos; una "base" de 10 proteínas, que se une directamente al anillo α exterior del núcleo de 20S y una "tapa" de 9 proteínas donde se une el complejo poliubiquitínico. De las 10 proteínas de la base, 6 presentan sitios activos de ATPasa. La asociación entre el núcleo de 20S y las partículas reguladoras de 19S entre sí, requiere de la unión de ATP a los sitios activos de unión de ATP en las partículas de 19S. La hidrólisis de ATP es necesaria para que el complejo entero pueda degradar una proteína doblada y ubiquitinizada. Aun así, no esta claro si la energía resultante de la hidrólisis es utilizada para el desdoblamiento del sustrato, o para permitir la entrada hacia el núcleo o ambos. Hasta el 2006 todavía no se ha resuelto la estructura de los proteosomas de 26S, pero se supone que tanto las partículas reguladoras de 19S y como las de 11S se unen de manera similar al anillo α del núcleo de 20S.
Algunos componentes individuales de las partículas de 19S, también tienen su participación en otros sistemas reguladores. Por ejemplo, Gankyrin es una recién descubierta proteína oncogénica, que forma parte de la partícula reguladora de 19S, que une a la ciclina dependiente de kinasa CDK4, y también juega un rol importante en el reconocimiento de la proteína P53 ubiquitinizada, gracias a su afinidad a la ligasa de ubiquitina MDM2. Esta proteína es antiapoptósica y se ha demostrado su exceso en algunas células tumorales como las del carcinoma hepatocelular.
Partículas reguladoras de 11S
Los núcleos de 20S también se pueden asociar a un segundo tipo de partículas reguladoras, denominadas de 11S, con forma de estructura heptagonal que no contienen sitios activos de ATPasa, siendo capaces de promover solo la degradación de pequeños péptidos y no de proteínas enteras. Se supone que esta incapacidad esta derivada de la imposibilidad de desdoblar sustratos mayores. La estructura recién mencionada es conocida con el nombre de PA28 o REG. Los mecanismos por los cuales se une al núcleo central a través de las terminales-C de sus subunidades proteicas, que inducen cambios en la conformación de los anillos α para abrir las partículas de 20S, parecerían ser similares a los de las partículas de 19S. La expresión de las partículas de 11S es inducida por el interferón gama y es responsable, junto con las subunidades β del inmunoproteosoma, de la generación de péptidos que se unen al complejo mayor de histocompatibilidad.
Armado
El armado del proteosoma es un proceso complejo debido a la cantidad de subunidades que se deben unir para formar el complejo activo. Las subunidades β son sintetizadas con una terminal-N "pro-peptídica" que es modificada post-traduccionalmente durante el armado de las partículas de 20S para no exponer el sitio activo. Las partículas de 20S son armadas con dos mitades, cada una de ellas consiste en 7 partes pro β adheridas a un anillo de 7 partes α. La asociación de los anillos β de las 2 mitades proteosómicas dispara la autolisis dependiente de treonina de los propétidos para exponer el sitio activo. estas interacciones entre partículas β son mediadas por uniones iónicas e hidrofóbicas donde se conservan las hélices alfa, que de ser corrompidas por mutaciones dañarían la capacidad de armado del proteosoma. La unión de las dos mitades, se inicia con la formación de las subunidades α en forma de anillo heptagonal, formando el marco para la asociación de los correspondientes anillos β. Poco se sabe de la formación de los anillos α.
En general, es poco lo que se sabe del armado y la maduración de las partículas reguladoras de 19S. Se cree que son armadas en dos distintos subcomponentes, la base contenedora de ATPasas y la tapa identificadora de proteínas ubiquitinizadas. Las 6 ATPasas en la base deben armarse en pares mediante interacciones Coiled coil. El orden en que los 19 componentes de la partícula reguladora son ensamblados, probablemente sea un mecanismo que evita la exposición de los sitios activos hasta que el armado sea completo.
Degradación de proteínas
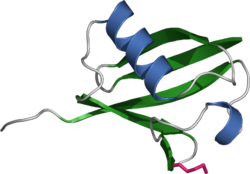 Diagrama de ubiquitina, proteína altamente consevada que sirve de marca molecular para señalar las proteínas a ser degradadas por el proteosoma.
Diagrama de ubiquitina, proteína altamente consevada que sirve de marca molecular para señalar las proteínas a ser degradadas por el proteosoma.Ubiquitinización y Objetivación (targeting)
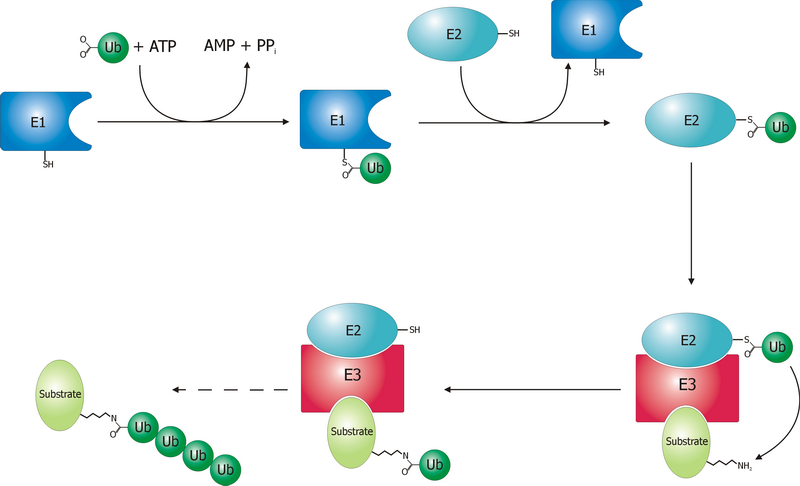
Las proteínas son marcadas para ser degradadas por el proteosoma mediante modificaciones covalentes de un residuo de lisina que requiere la reacción coordinada de tres enzimas. Como primer paso una enzima activadora de ubiquitina ¡'¡' (conocida como E1) hidroliza el ATP y adenila una molécula de ubiquitina. Esto es luego transferido al sitio activo de la cisteína de E1¡'¡', coincidiendo con la adenilación de una segunda ubiquitina. Esta ubiqutiina adenilada es entonces transferida a la cisteína ¡'¡'de una segunda enzima, "enzima conjugadora de ubiquitina" (E2). Por último, un integrante de una clase de enzimas de alta diversidad, conocida como ligasa de ubiquitina (E3), reconoce la proteína específica a ser ubiquitinizada y cataliza la transferencia de ubiquitina de E2 a la proteína fijada. Una proteína a ser degradada debe ser marcada mínimamente con 4 monómeros de ubiquitina en forma de cadena para que sea reconocida por el sistema ubiquitino proteosómico. Es entonces la E3 la que confiere la especificidad del sustrato al sistema¡'¡'. El número de E1, E2 y E3 expresados dependen del organismo y del tipo de célula. Existen muchas enzimas E3 diferentes presentes en humanos, indicando la gran cantidad de proteínas posibles de degradar por el sistema ubiquitino proteosómico.
El mecanismo por el cual una proteína poliubiquitinizada es objetivada por el proteosoma no se entiende completamente. Las proteínas ubiquitino-receptoras tienen una zona terminal-N de ubiquitina (UBL) y una o más zonas ubiquitino-asociadas (UBA). La zona UBL es reconocida por las zonas reguladoras de 19S y las zonas UBA unen la ubiquitina mediante enlaces de triple hélice. Estas proteínas receptoras podría escoltar las proteínas poliubiquitinizadas al proteosoma, aunque las especificaciones de estas interacciones y de su regulación no estén en claro.La proteína ubiquitina tiene 76 aminoácidos y fue llamada así debido a su naturaleza ubicua; tiene una secuencia altamente conservada y se encuentra en todos los organismos eucariotas. En eucariontes, los genes que codifican la ubiquitina se encuentran en tandas repetidas, posiblemente debido a la alta demanda de transcripción de estos genes para cumplir con los requerimientos de ubiquitina de la célula. Se ha sugerido que la ubiquitina es la proteína que evoluciona más lentamente identificada hasta la fecha.
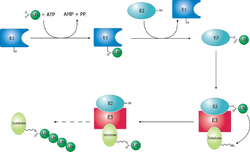 El proceso de ubiquitinización.
El proceso de ubiquitinización.Desdoblamiento y Translocación
Después de que una proteína es ubiquitinizada, es reconocida por las partículas reguladoras de 19S en un proceso de unión dependiente de ATP. El sustrato de la proteína debe entrar en el interior de la partícula de 20S para tomar contacto con los sitios activos. Debido a que el canal central es estrecho y esta limitada la entrada por los terminales-N de las colas de las subunidades del anillo α, los sustratos deben ser por lo menos parcialmente desdoblados para poder entrar a la cavidad catalítica. El pasaje del sustrato desdoblado al núcleo se llama translocation y ocurre después de la desubiquitinación. Aun así el orden en que los sustratos son desubiquitinizados y desdoblados no esta del todo claro. Cual de estos dos reactivos es el limitante en la reacción de proteólisis, depende del sustrato. En algunas proteínas el limitante es el desdoblamiento, mientras que la desubiquitinización es la reacción más lenta en otras. El punto hasta el que los sustratos deben ser desdoblados antes de la translocation no se conoce, pero una estructura terciaria, y en particular, la no existencia de interacciones no locales como enlaces de disulfuro es suficiente para inhibir la degradación.
Las puertas formadas por las subunidades α evitan que los péptidos mayores que 4 residuos entren en la partícula de 20S. Las moléculas de ATP ¡'¡' antes del reconocimiento inicial son hidrolizadas antes de la translocation, aunque existe un desacuerdo si esta energía es necesitada para el desdoblamiento del sustrato o la apertura de la puerta. El proteosoma de 26S armado puede degradar proteínas desdobladas en presencia de analog atp ¡'¡' no hidrolizable, pero no puede degradar proteínas desdobladas, indicando que la energía de la hidrólisis de ATP es usado para el desdoblamiento del sustrato. El pasaje del sustrato desdoblado a través de la puerta ocurre por difusión facilitada si la zona reguladora de 19S esta en estado de ¡'¡' ATP.
El mecanismo para desdoblar proteínas globulares es general, pero de alguna forma depende de la secuencia de los aminoácidos a desdoblar. Largas secuencias alternantes entre glicina y alanina han demostrado que inhiben el desdoblamiento del sustrato, reduciendo la eficiencia de la degradación proteosómica. Esto resulta en la liberación de bioproductos parcialmente degradados, posiblemente debido al desemparejamiento entre la hidrólisis de ATP y los procesos desdoblantes. Esa repetición de alanina/glicina también es encontrada en la naturaleza, por ejemplo en la fibroina de la seda o en ciertos genes del virus de Epstein-Barr que llevan esta secuencia pueden estallar el proteosoma, ayudando al virus a propagarse, impidiendo la presentación de antígenos en el complejo mayor de histocompatibilidad.
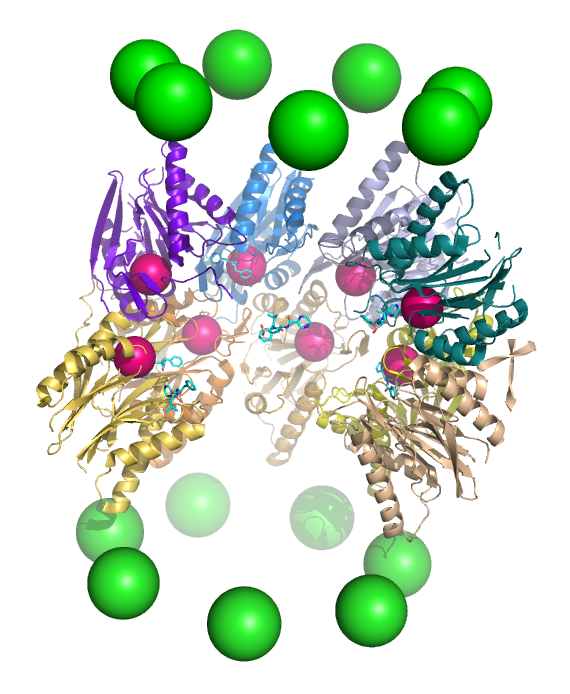
 A cutaway view of the proteasome 20S core particle illustrating the locations of the active sites. The α subunits are represented as green spheres and the β subunits as protein backbones colored by individual polypeptide chain. The small pink spheres represent the location of the active-site threonine residue in each subunit. Light blue chemical structures are the inhibitor bortezomib bound to the active sites.
A cutaway view of the proteasome 20S core particle illustrating the locations of the active sites. The α subunits are represented as green spheres and the β subunits as protein backbones colored by individual polypeptide chain. The small pink spheres represent the location of the active-site threonine residue in each subunit. Light blue chemical structures are the inhibitor bortezomib bound to the active sites.Proteólisis
La proteólisis llevada a cabo por las subunidades β del núcleo de 20S es a través de un ataque nucleófilo dependiente de treonina. este mecanismo depende de la asociación con una molécula de agua para la deprotonización¡'¡' del reactivo de hidroxilo de treonina. La degradación ocurre en la cámara central formada por la asociación de los dos anillos β y normalmente no libera productos parcialmente degradados, sino que reduce el sustrato a polipéptidos cortos, generalmente de 7 a 9 aminoácidos de largo, aunque puede variar de 4 a 25, dependiendo del organismo y del sustrato. El biomecanismo que determina la largo del producto no ha sido descrito totalmente. Aunque las tres subunidades β catalíticas comparten un mecanismo común, tienen pequeñas diferenciaciones en los sustratos que son consideraradas como en base a quimotripsina, en base a tripsina y en base a PHGH. Estas variaciones en la especifidad son el resultado de contactos interatómicos con residuos locales cerca de los sitios activos de cada subunidad. Cada subunidad β catalítica también posee un residuo de lisina requerido para la proteólisis.
Aunque los proteosomas producen fragmentos peptídicos muy cortos, en algunos casos estos productos son biológicamente activos y molecularmente funcionales. Ciertos factores de transcripción, incluyendo un componente del complejo presente en mamíferos NF-kB, son sintetizados como precursores inactivos que al ubiquitinizarse y degradarse proteosómicamente, se convierten en activos. Esta actividad requiere que el proteosoma ¡'¡' el sustrato proteico internamente en vez¡'¡'. Se ha sugerido que grandes ciclos ¡'¡' en la superficie de estas proteínas sirven de sustrato proteosómico y entran en la cavidad central mientras que la mayor parte de la proteína se mantienen afuera. Efectos similares han sido observados en proteínas de levaduras. Este mecanismo de degradación selectiva es conocido como "proceso de regulación dependiente de ubiquitina/proteosomas" (RUP).
Degradación independiente de ubiquitina
Aunque la mayoría de los sustratos proteosómicos deben ser ubiquitinizados para poder ser degradados, hay algunas excepciones a la regla. Especialmente si el proteosoma juega un rol en la normal procesamiento de una proteína posttranslational. La activación proteosómica de NF-k3 mediante el proceso de p105 en P50 mediante proteólisis interna es un ejemplo. Algunas proteínas hipotéticamente inestables debido a la región no estructurada intrínseca, son degradadas de manera distinta, sin dependencia de ubiquitina. El ejemplo mejor conocido es un sustrato proteósomico independiente de ubiquitina es la enzima ornithine decarboxylase. Mecanismos claves en la regulación del ciclo celular, como la P53, también se han reportado de usar mecanismo independientes de ubiquitina. Por último, proteínas estructuralmente anormales, mal dobladas o altamente oxificadas¡'¡' son sujetos de procesos de degradación independientes de ubiquitina y de 19S bajo condiciones de estrés celular.¡'¡'
Evolución
 El complejo armado de hs1V (azul) y hs1U (rojo) de una ¡'¡'. Este complejo ¡'¡' se supone como el ancestro de la proteosoma moderna.
El complejo armado de hs1V (azul) y hs1U (rojo) de una ¡'¡'. Este complejo ¡'¡' se supone como el ancestro de la proteosoma moderna.La proteosoma de 20S es tanto ubicua como esencial en los eucariontes. Algunos procariontes, incluyendo alguans bacterias del orden actinomycetales tienen homólogos del proteosomas de 20S, mientras que la mayoría de las bacterias posee los genes hsIV y hsIU, cuyos productos genéticos son una proteasa ¡'¡' conformada por ¡'¡' anillos y ATPasa. la proteína del hsIV es considerada como el ancestro del proteosoma de 20S. El hsIV, por lo general, no es esencial en las bacterias y no todas las bacterias lo poseen. Algunos protistas poseen tanto el sistema del proteosoma de 20 S y el hsIV.
Análisis de secuencia¡'¡' sugieren que las subunidades catalíticas β divergieron evolutivamente antes que las subunidades de predominancia estructural α. En bacterias con proteosomas de 20S, las subunidades β tienen mayor coincidencia secuencial con respecto a subunidades β de archaeas y eucariontes, mientras que las subunidades α la coincidencia es menor. La presencia de proteosomas de 20S en bacterias se podría adjudicar a transferencias horizontales genéticas¡'¡' mientras que la diversificación entre las subunidades eucariotas se adscriben a eventos múltiples de duplicación génica¡'¡'.
Control del ciclo celular
La progresión del ciclo celular es controlada por la acción ordenada de kinasas dependientes de ciclinas (CDK) activadas por ciclinas específicas que demarcan las fases del ciclo celular. Las ciclinas mitóticas que persisten en la célula por unos pocos minutos, tienen uno de los tiempos de vida más cortos de todas las proteínas intracelulares. Una vez que el complejo de CDK + ciclina ha realizado su función, la ciclina asociada es poliubiquitinizada y degradada por el proteosoma, dándole continuidad al ciclo celular. Particularmente, la salida de la mitosis requiere la disociación dependiente de proteosomas del componente regulador ciclina B del complejo factor de promoción de mitosis. En células de vertebrados ¡'¡'.
checkpoints tempranos como el ¡'¡' entra la fase G1 y la fase S similimarmente involucran la degradación proteosómica de la ciclina A, cuya ubiquitinización es promovida por el complejo promotor de anafase (APC), un E3 ligasa de ubiquitina. El APC y el ¡'¡' complejo proteínico (complejo SCF) son 2 reguladores ¡'¡' de la degradación ciclina y el checkpoints. El SCF es regulado por el APC mediante la ubiquitinización del componente ¡'¡', que previene la actividad SCF antes de la transición de la fase G1 a la fase S.
Regulación del crecimiento en plantas
== En las plantas, la utilización de auxinas o fitohormonas, que ordenan la dirección y el tropismo del crecimiento, induce el marcado para la degradación proteosómica una clase de factor de transcripción represivo conocido como proteínas Aux/IAA. Estas proteínas son ubiquitinizadas mediante SCFTIR1 o SCF en complejo con el receptor de auxinas TIR1. La degradación de las proteínas Aux/IAA libera los factores de transcripción en el auxin response factor (ARF) family¡'¡' e induce la expresión de genes relacionados con el ARF. Las consecuencias a nivel celular de la activación del ARF depende del tipo de planta y de la etapa de desarrollo de la misma, pero en general dirigen el crecimiento de raíces y venas de hojas ¡'¡'. La especifidad de la respuesta de la liberación del ARF se piensa que es mediada por la especificidad de la respuesta en el apareamiento de proteínas individuales de ARF y Aux/IAA. ==
Apoptosis
Tanto señales internas como externas pueden inducir la apoptosis, o muerte celular programada. La consecuente deconstrucción de los componentes celulares es llevada a cabo principalmente por proteasas específicas llamadas caspasas, pero el proteosoma también juega diversos e importantes roles en los procesos apoptósicos. Durante la apoptosis, se ha observado a los proteosomas cercanos al núcleo trasladando al exterior a los blebs¡'¡' de la membranas, características de la apoptósis.
La inhibición proteosómica tiene diferentes efectos en la inducción apoptósica de diferentes tipos celulares. El proteosoma, generalmente, no es requerido para la apoptosis, aunque su inhibición es pro- apoptósica en la mayoría de los tipos celulares que han sido estudiados. Sin embargo, algunas especies celulares- en especial tejidos primarios de células en fase estacionaria y diferenciadas como los linfocitos T y las neuronas, no inducen la apoptosis ante la exposición a inhibidores proteosómicos. El mecanismo de este efecto no esta del todo claro, pero se supone que es específico en células en estados estacionarios o del resultado de una actividad distinta de la quinasa proapoptósica JNK. La habilidad de los inhibidores proteosómicos de inducir la apoptósis en células de rápida división ha sido explotada últimamente en el desarrollo de agentes quimioterapéuticos.
Respuesta al estrés celular
En respuesta a situaciones de estrés célular, como pueden ser infecciones, cambios bruscos de temperaturas o daño oxidativo, se expresan proteínas de choque térmico, que identifican proteínas mal o no dobladas y las marca para la degradación proteosómica. Tanto Hsp27 como Hsp90 -proteínas chaperonas son implicadas en el incremento de la actividad del sistema ubiquitino proteosómico, aunque no sean participantes directos del proceso. Por el otro lado la proteína Hsp70 ¡'¡' en la superficie de las proteínas mal dobladas y utiliza ligasas de ubiquitina E3 como CHIP para marcar proteínas para la degradación proteosómica. La proteína CHIP (termino carboxilo de la proteína interactuante con Hsp70) es regulada mediante la inhibición de las interacciones entre la enzima E3 CHIP y su compañero de ¡'¡' E2.
Mecanismos similares existen para promover la degradación de proteínas altamente dañadas por oxidación¡'¡' mediante el sistema proteosómico. En particular, los proteosomas localizados cerca del núcleo son regulados por PARP y activados suelen degradar histonas oxidificadas¡'¡' inapropiadas. Las proteínas oxidificadas ¡'¡' suelen formar gigantes agregados amorfos en la célula, pueden ser degradados directamente por el núcleo de 20S sin la regulación de la partícula de 19S y no requieren hidrolización de ATP o ubiquitinización, como sea, altos niveles de daño oxidativo, incrementan el grado de cruzamiento ¡'¡' entre fragmentos de proteínas, ¡'¡' resistentes a la proteólisis. Gran cantidad y tamaño de esos agregados altamente oxidificados están relacionados con el envejecimiento.
la deteriorada actividad proteosómica ha sido sugerida como una explicación para algunas de las enfermedades neurodegenerativas de avanzada edad que comparten la presencia característica de proteínas mal dobladas, como la Enfermedad de Parkinson o el Mal de Alzheimer. En estas enfermedades, grandes agregados insolubles de proteínas mal dobladas pueden formarse y causar neurotoxicidad, a través de mecanismos que todavía no son bien entendidos. El descenso de la actividad proteosómica ha sido sugerido como una de las causas de la agregación y formación de cuerpos de lewi¡'¡' en la Enfermedad de Parkinson. Esta hipótesis es avalada por la observación que modelos de levaduras de Parkinson son más susceptibles a la toxicidad de α-sinuclein, la principal proteína componente de los cuerpos de lewy, bajo condiciones de baja actividad proteosómica.
Rol en el sistema inmunológico
Los proteosomas juegan un rol secundario pero a la vez crítico en el funcionamiento del sistema inmune adaptativo. Los péptidos antígenicos son presentados por las proteínas del sistema mayor de histocompatibilidad clase I (MHC I) en la superficie de la células presentadoras de antígenos (CPA). Estos péptidos son productos de la degradación proteaosómica de proteínas originadas por el patógeno invasor. Aunque los proteosomas comunes pueden participar de este proceso, un complejo especializado compuesto de proteínas cuya expresión está inducida por el interferón gamma, se encarga de producir péptidos con la composición y tamaños óptimos para la unión del MHC I. De entre las proteínas que aumentan su expresión ante la respuesta inmunológica, se encuantran la partícula reguladora de 11S, cuya principal función biológica conocida es la regulación de la producción de ligandos de MHC y las subunidades β especializadas llamadas β1i, β2i y β5i con afindades de sustrato específicas. El complejo formado con las subunidades β especializadas es llamado inmunoproteosoma.
La fuerza de la unión MHC clase I y su ligando depende de la composición del extremo C-Terminal del ligando, los péptidos se unen mediante uniones puente de hidrógeno y mediante el contacto cercano con la región llamada "B pocket" de la superficie del MHC I. Varios alelos de MHC clase I presentan mayor afinidad por los residuos C-terminales hidrofóbicos de los péptidos; el inmunoproteosoma tiende a generar péptidos con extremos C-terminales hidrofóbicos.
Debido a su rol en la generación de la forma activada de NF-kB, un regulador antiapoptótico y pro-inflamatorio de la expresión de citoquinas, la actividad proteaosómica ha sido relacionada con las enfermedades autoinmunes e inflamatorias. Niveles altos de actividad proteosómica están correlacionados con la presencia de enfermedades y han sido implicados en enfermedades autoinmunes como el lupus eritematoso y la artritis reumatoide.
Inhibidores proteosómicos
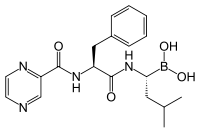 Estructura química del Bortezomib, inhibidor proteosómico usado en la quimioterapia que es especialmente efectivo con el mieloma múltiple.
Estructura química del Bortezomib, inhibidor proteosómico usado en la quimioterapia que es especialmente efectivo con el mieloma múltiple.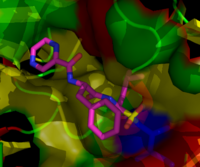 Bortezomib enlazado a la partícula nuclear en un proteosama de levadura. La molécula de bortezomib está en el centro coloreada según el tipo de átomo (Carbono = rosa, nitrógeno = azul, oxígeno = rojo y boro = amarillo), rodeada por la superficie de la proteína local¡'¡'. El remiendo azul es el reisudo catalítico de treonina cuya actividad esta bloqueada por la presencia del bortezomib.
Bortezomib enlazado a la partícula nuclear en un proteosama de levadura. La molécula de bortezomib está en el centro coloreada según el tipo de átomo (Carbono = rosa, nitrógeno = azul, oxígeno = rojo y boro = amarillo), rodeada por la superficie de la proteína local¡'¡'. El remiendo azul es el reisudo catalítico de treonina cuya actividad esta bloqueada por la presencia del bortezomib.Los inhibidores proteosómicos tienen una efectiva actividad antitumoral en cultivos celulares, induciendo la apoptosis por la disrupción de la degradación reguladora de proteínas claves para ciclo celular. Este acercamiento a la selección inducida de la apoptosis en células tumorales ha probado efectividad en modelos animales y pruebas en humanos. Bortezomib, una moléluca desarrollada por millenium pharmaceuticals y conocida comercialmente como velcade, es el primer inhibidor proteosómico en alcanzar el uso clínico como agente quimioterapéutico. Bortezomib es usado en el tratamiento del mieloma múltiple. El mieloma múltiple ha sido observado incrementando el nivel de proteosomas en el plasma sanguíneo, que disminuye al realizarse una quimioterapia efectiva. Estudios en animales indicaría que bortezomib podría tener un efecto clínico significativo en el tratamiento de cáncer pancreático. Estudios preclínicos y clínicos tempranos han empezado a investigar la efectividad del uso de bortezomib en otros cánceres relacionados con células B, en especial algunos tipos de Linfomas no-Hodgkins.
La molécula ritonavir, llamada comercialmente Norvir, fue desarrollada como un inhibidor proteásico para atacar la infección del VIH. Sin embargo, se ha demostrado que inhibe proteosomas así como proteasas libres; específicamente, la actividad proteosómica de la quimotripsina, mientras la actividad de la tripsina de alguna forma se ve aumentada. Estudios en animales sugieren que el ritonavir quizá tenga efectos inhibitorios en el crecimiento de las células de gliomas¡'¡'.
Los inhibidores proteosómicos también han mostrado promesas en el tratamiento de enfermedades autoinmunes en estudios en animales. Por ejemplo, estudios en ratones con trasplantes de piel humana, descubrieron una reducción en el tamaño de las lesiones de psoriasis, después del tratamiento con un inhibidor proteosómico. Los inhibidores proteosómicos también han mostrado efectos positivos en roedores con asma.
El marcado y la inhibición del proteosoma es de interés en el laboratorio, tanto para estudios in vivo o in vitro de la actividad proteosómica celular, el más común inhibidor proteosómico usado en laboratorio es la Lactacystin, un producto natural sintetizado por la bacteria Streptomyces. Inhibidores fluorescentes también han sido desarrollados para poder marcar específicamente los sitios activos del proteosoma armado.
Referencias
- ↑ Peters JM, Franke WW, Kleinschmidt JA (March de 1994). «Distinct 19 S and 20 S subcomplexes of the 26 S proteasome and their distribution in the nucleus and the cytoplasm» J Biol Chem. Vol. 269. n.º 10. pp. 7709–18. PMID 8125997.
- ↑ a b c Lodish H, Berk A, Matsudaira P, Kaiser CA, Krieger M, Scott MP, Zipursky SL, Darnell J (2004). «3», Molecular cell biology, 5th ed. edición, New York: W.H. Freeman and CO, pp. 66–72. ISBN 0-7167-4366-3.
- ↑ a b Nobel Prize Committee (2004). «Nobel Prize Awardees in Chemistry, 2004». Consultado el 2006-12-11.
- ↑ Etlinger JD, Goldberg AL (January de 1977). «A soluble ATP-dependent proteolytic system responsible for the degradation of abnormal proteins in reticulocytes» Proc. Natl. Acad. Sci. U.S.A.. Vol. 74. n.º 1. pp. 54–8. PMC 393195. DOI 10.1073/pnas.74.1.54. PMID 264694.
- ↑ Ciehanover A, Hod Y, Hershko A (1978). «A heat-stable polypeptide component of an ATP-dependent proteolytic system from reticulocytes» Biochem Biophys Res Commun.. Vol. 81. n.º 4. pp. 1100–1105. DOI 10.1016/0006-291X(78)91249-4. PMID 666810.
- ↑ Goldknopf IL, Busch H (1977). «Isopeptide linkage between nonhistone and histone 2A polypeptides of chromosomal conjugate-protein A24» Proc Natl Acad Sci USA. Vol. 74. n.º 3. pp. 864–8. DOI 10.1073/pnas.74.3.864. PMID 265581.
- ↑ Ciechanover A (2000). «Early work on the ubiquitin proteasome system, an interview with Aaron Ciechanover» Cell Death Differ. Vol. 12. n.º 9. pp. 1167–77. DOI 10.1038/sj.cdd.4401691. PMID 16094393.
- ↑ Tanaka K, Waxman L, Goldberg AL (June de 1983). «ATP serves two distinct roles in protein degradation in reticulocytes, one requiring and one independent of ubiquitin» J. Cell Biol.. Vol. 96. n.º 6. pp. 1580–5. PMC 2112434. DOI 10.1083/jcb.96.6.1580. PMID 6304111.
- ↑ Hough R, Pratt G, Rechsteiner M (June de 1987). «Purification of two high molecular weight proteases from rabbit reticulocyte lysate» J. Biol. Chem.. Vol. 262. n.º 17. pp. 8303–13. PMID 3298229.
- ↑ Hershko A (2005). «Early work on the ubiquitin proteasome system, an interview with Avram Hershko» Cell Death Differ. Vol. 12. pp. 1158–1161. DOI 10.1038/sj.cdd.4401709. PMID 16094391.
- ↑ Kopp F et al. (1986). «Size and shape of the multicatalytic proteinase from rat skeletal muscle» Biochim Biophys Acta. Vol. 872. n.º 3. pp. 253–60. PMID 3524688.
- ↑ Löwe J et al. (1995). «Crystal structure of the 20S proteasome from the archaeon T. acidophilum at 3.4 Å resolution» Science. Vol. 268. pp. 533–539. DOI 10.1126/science.7725097. PMID 7725097.
- ↑ Wang J, Maldonado MA (2006). «The Ubiquitin-Proteasome System and Its Role in Inflammatory and Autoimmune Diseases» Cell Mol Immunol. Vol. 3. n.º 4. pp. 255. PMID 16978533.
Véase también
Enlaces externos
- Partes de la célula.
- Mecanismo de proteólisis intracelular.
- Funcionamiento del proteosoma animado.
- Noticia sobre la aprobación del bortezomib.
Categorías: Biología molecular | Biología celular | Complejos proteicos






Wikimedia foundation. 2010.