- Plasmodium
-
Plasmodium
?Plasmodium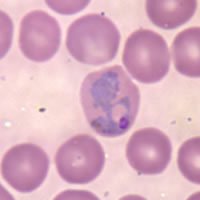
Trofozoito de un Plasmodium.Clasificación científica Reino: Protista (sin clasif.) Alveolata Filo: Apicomplexa Clase: Aconoidasida Orden: Haemosporida Género: Plasmodium Especies Para más especies, véase textoPlasmodium es un género de protistas del filo Apicomplexa, clase Aconoidasida, orden Haemosporida y familia Plasmodiidae del que se conocen más de 175 especies.[1] El parásito siempre tiene dos huéspedes en su ciclo vital: un mosquito que actúa como vector y un huésped vertebrado. Al menos diez especies infectan al hombre. Para humanos hay cuatro especies de Plasmodium que provocan la malaria o paludismo: P. falciparum, P. malariae, P. ovale y P. vivax, de las cuales la primera es la más virulenta y la que produce la mayor mortalidad. Otras especies infectan a otros animales, incluyendo aves, reptiles y roedores.
Contenido
Ciclo del parásito
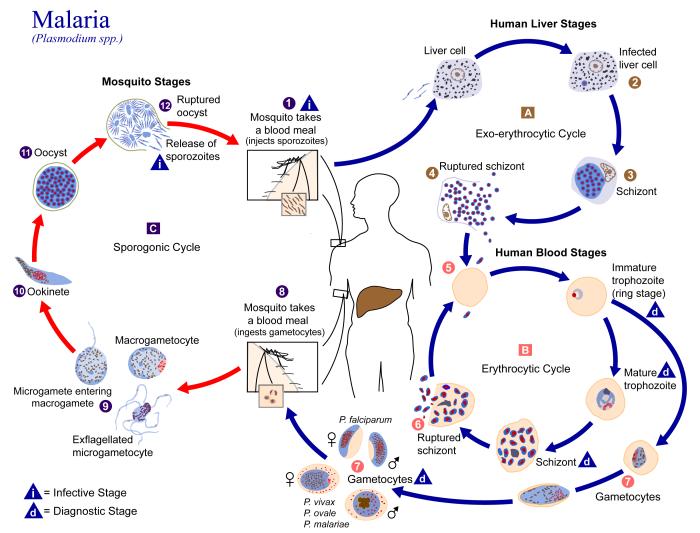
En el ciclo del Plasmodium existe un agente vector (la hembra de un mosquito), donde el Plasmodium se reproduce sexualmente y un hospedador vertebrado intermediario (el ser humano u otro animal). La siguiente explicación puede seguirse en la figura de abajo.
- Etapas exo-eritrocíticas o hepáticas (A en la figura). Tras la picadura del mosquito, éste inocula el parásito existente en su saliva en la sangre o en el sistema linfático[2] del huésped (1). En ese momento, el Plasmodium se encuentra en la fase de su ciclo conocida como esporozoito. Los esporozoitos pasan al torrente sanguíneo hasta que llegan a los hepatocitos del hígado (2). Allí se multiplican por esquizogénesis (disgregación) formando el esquizonte hepático (3), tras lo cual se rompe el hepatocito, apareciendo un nuevo estadio del Plasmodium, el merozoito (4). Aquí hay un primer ciclo asexual, en el que los merozoitos pueden o bien reinfectar hepatocitos o bien volver de nuevo al torrente sanguíneo, donde penetran en los eritrocitos.
- Etapas eritrocíticas o sanguíneas (B). En los eritrocitos, los merozoitos comienzan a alimentarse de la parte proteíca de la hemoglobina contenida en éstos, apareciendo entonces el trofozoito (5). Nuevamente por esquizogénesis se multiplica en el interior de dichas células, formándose el esquizonte hemático. También se rompe la célula, en este caso el eritrocito, liberando nuevos merozoitos (6). La mayoría de los merozoitos continúan con este ciclo replicativo infectando nuevos eritrocitos, pero algunos se convierten en gametocitos, masculinos y femeninos (7).
 Ciclo de vida del Plasmodium.
Ciclo de vida del Plasmodium.
- Etapas en el mosquito (C). Si el individuo infectado es nuevamente picado por un mosquito, los gametocitos masculinos y femeninos pasan al mosquito (8). En el interior de éste se diferencian en gametos (4-8 microgametos por cada gametocito masculino y una macrogameta por cada gametocito femenino) y al fusionarse ambos gametos, se producen los zigotos (9). Los zigotos, a su vez, se convierten en oocinetos móviles y alargados (10), que invaden la pared intestinal del mosquito, donde se desarrollan en ooquistes (11). Los ooquistes crecen, se rompen y liberan una nueva generación de esporozoitos (12), que hacen su camino a la glándulas salivares del mosquito. Es en esta fase en la que el Plasmodium puede volver a ser inyectado en el huéped.
En algunas especies de Plasmodium, el esquizonte hepático puede permacer en estado latente, en forma de hipnozoito. La reactivación de los hipnozoitos puede ocurrir hasta un máximo de 30 años después de la infección inicial en el ser humano. No se conoce todavía cuáles son los factores de reactivación. Entre las especies que forman hipnozoitos están P. malariae, P. ovale y P. vivax. La reactivación no se produce en las infecciones por P. falciparum. No se sabe si la reactivación del hipnozoito se produce en las restantes especies que infectan a los seres humanos, pero es posible que este sea el caso.
La transición desde la etapa hepática a la eritrocítica ha sido confusa hasta hace poco tiempo. En el año 2006[3] se demostró que el parásito sale de los hepatocitos en merosomas que contienen cientos o miles de merozoitos. Se ha demostrado posteriormente[4] que estos merosomas se dirigen a los capilares pulmonares y lentamente se desintegran en unas 48-72 horas liberando los merozoitos. La invasión de los eritrocitos es mayor cuando el flujo sanguíneo es lento y las células están perfectamente empaquetadas: estas dos condiciones se dan juntas en los capilares alveolares.
Evolución
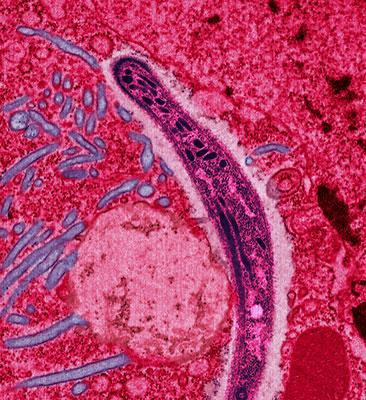
 Esporozoito de Plasmodium.
Esporozoito de Plasmodium.Se cree que los apicomplejos comenzaron infectando la pared intestinal del huésped con una ruta de propagación fecal-oral. En algún punto de la evolución, el parásito evolucionó a la capacidad de infectar el hígado de huésped. Este patrón se observa en el género Cryptosporidium. En algún otro punto de la evolución, un apicomplejo desarrolló la capacidad de infectar las células de la sangre y de sobrevivir e infectar a los mosquitos. Una vez que la transmisión del mosquito quedó firmemente establecida, se perdió la anterior vía de transmisión fecal-oral. La teoría actual (2007) sugiere que los géneros Plasmodium, Haemoproteus y Hepatocystis evolucionaron a partir de especies de Leukocytozoon. Los parásitos del género Leukocytozoon infectan a los leucocitos (glóbulos blancos) y a las células del hígado y del bazo y se transmiten por medio de moscas negras (Simulium), un extenso género de moscas relacionadas con los mosquitos.
Los leucocitos, la mayoría de los hepatocitos y las células del bazo realizan activamente la fagocitosis de partículas, haciendo la entrada en la célula más fácil para el parásito. El mecanismo de entrada en los eritrocitos de las especies del género Plasmodium está todavía poco clara, teniendo en cuenta que la entrada se realiza en menos de 30 segundos. Todavía no se sabe si este mecanismo evolucionó antes que los mosquitos se convirtieran en los principales vectores de transmisión Plasmodium.
Estudios realizados con microscopía electrónica en los 70s, y estudios epigenéticos recientes indican que el parásito se une a la membrana del eritrocito y promueve la formación de una invaginación, a través de la cual el merozoito (que es el estadio que invade a los eritrocitos) logra penetrar los hematíes. Las primeras uniones son débiles y reversibles y se dan entre proteínas del parásito y de membrana del eritrocito, estas uniones le ayudan al merozoito a orientarse y, de esta forma, facilitar la formación de uniones fuertes merozoito-eritrocito , gracias a las cuales la invaginación de la célula sanguínea se lleva a cabo.
La evolución de Plasmodium comenzó hace cerca de 130 millones de años. Este período es coincidente con la rápida propagación de las angiospermas (plantas con flores). Se cree que esta expansión de las angiospermas es debida a por lo menos un evento de duplicación genómica. Parece probable que el aumento en el número de flores haya llevado a un aumento en el número de mosquitos y a su contacto con los vertebrados. Los mosquitos se desarrollaron en lo que hoy es Suramérica hace unos 230 millones de años. Se conocen más de 3500 especies de mosquitos, pero hasta la fecha su evolución no se conoce en detalle, por lo que existen lagunas en nuestro conocimiento de la evolución de Plasmodium.
Pueden actuar como vectores de la malaria los mosquitos de los géneros Culex, Anopheles, Culiceta, Mansonia y Aedes. Los mosquitos que se conoce que actúan como vectores de la malaria humana (más de 100 especies), pertenecen todos al género Anopheles. La malaria en las aves es comúnmente propagada por especies pertenecientes al género Culex. Sólo las hembras de los mosquitos pican. Los mosquitos de ambos sexos se alimentan de néctar, pero las hembras precisan de una o más ingestiones de sangre para realizar la puesta de huevos, pues el contenido en proteínas del néctar es muy bajo.
Actualmente parece probable que las aves fueran el primer grupo infectado por Plasmodium seguidas por los reptiles, probablemente los lagartos. En algún punto de la evolución fueron infectados primates y roedores. El resto de las especies fuera de estos grupos parece probable que fueran infectadas en tiempos relativamente recientes.
Especies de Plasmodium
P. achiotense, P. achromaticum, P. aegyptensis, P. aeuminatum, P. agamae, P. anasum, P. atheruri, P. azurophilum, P. balli, P. bambusicolai, P. basilisci, P. berghei, P. bigueti P. brasilianum, P. brygooi, P. booliati, P. bubalis, P. bucki, P. coatneyi, P. cathemerium, P. cephalophi, P. chabaudi, P. chiricahuae, P. circularis, P. cnemidophori, P. coatneyi, P. coggeshalli, P. colombiense, P. corradettii, P. coturnix, P. coulangesi, P. cuculus, P. popo, P. cyclopsi, P. cynomolgi, P. diminutivum, P. diploglossi, P. dissanaikei, P. dominicana, P. durae, P. egerniae, P. elongatum, P. eylesi, P. fabesia, P. fairchildi, P. falciparum, P. fallax, P. fieldi, P. foleyi, P. forresteri, P. floridense, P. fragile, P. garnhami, P. gallinaceum, P. giganteum, P. giovannolai, P. girardi, P. gonatodi, P. gonderi, P. georgesi, P. gracilis, P. griffithsi, P. guanggong, P. gundersi, P. guyannense, P. heischi, P. hegneri, P. hermani, P. heteronucleare, P. hexamerium, P. holaspi, P. huffi, P. hylobati, P. icipeensis, P. inopinatum, P. inui, P. jefferi, P. josephinae, P. juxtanucleare, P. kempi, P. knowlesi, P. kentropyxi, P. leanucteus, P. lemuris, P. lophurae, P. lepidoptiformis, P. lygosomae, P. mabuiae, P. mackerrasae, P. maculilabre, P. maior, P. malariae, P. marginatum, P. matutinum, P. mexicanum, P. minasense, P. morulum, P. nucleophilium, P. octamerium, P. odocoilei, P. ovale, P. papernai, P. paranucleophilum, P. parvulum, P. pedioecetii, P. pelaezi, P. percygarnhami, P. petersi, P. pifanoi, P. pinotti, P. pinorrii, P. pitheci, P. pitmani, P. polare, P. praecox, P. reichenowi, P. relictum, P. rhadinurum, P. rhodaini, P. robinsoni, P. rouxi, P. sandoshami, P. sasai, P. schweitzi, P. silvaticum, P. simium, P. semiovale, P. shortii, P. smirnovi, P. subpraecox, P. tenue, P. tejerai, P. tomodoni, P. torrealbai, P. traguli, P. tribolonoti, P. tropiduri, P. uilenbergi, P. watteni, P. wenyoni, P. vacuolatum, P. vastator, P. vaughani, P. vinckei, P. vivax, P. volans, P. yoelii, P. youngi, etc.
Enlaces externos
 Wikimedia Commons alberga contenido multimedia sobre Plasmodium.
Wikimedia Commons alberga contenido multimedia sobre Plasmodium.
Referencias
- ↑ Chavatte JM, Chiron F, Chabaud A, Landau I. (2007) Probable speciations by "host-vector 'fidelisation'": 14 species of Plasmodium from Magpies. Parasite 14(1):21-37
- ↑ [http://www.hhmi.org/news/menard20060122.html HHMI Staff (22 January 2006) "Malaria Parasites Develop in Lymph Nodes" HHMI News Howard Hughes Medical Institute
- ↑ Sturm A, Amino R, van de Sand C, Regen T, Retzlaff S, Rennenberg A, Krueger A, Pollok JM, Menard R, Heussler VT. (2006) Manipulation of host hepatocytes by the malaria parasite for delivery into liver sinusoids. Science 313(5791):1287-1290
- ↑ Baer K, Klotz C, Kappe SH, Schnieder T, Frevert U. (2007) Release of Hepatic Plasmodium yoelii Merozoites into the Pulmonary Microvasculature. PLoS Pathog. 3(11):e171
Categorías: Protozoos | Agentes biológicos patógenos | Parásitos | Aconoidasida | Eritrocito
Wikimedia foundation. 2010.