- Recombinación V(D)J
-
La recombinación V(D)J es un mecanismo de recombinación genética que se da en vertebrados por el cual se selecciona y ensambla al azar segmentos de genes que codifican proteínas específicas con papeles importantes en el sistema inmunitario.[1] Esta recombinación específica de localización genera un repertorio diverso de receptores de linfocitos T (TCR) y moléculas de inmunoglobulina (Ig) que son necesarios para el reconocimiento de diversos antígenos bacterianos, víricos y de parásitos, así como de células disfuncionales, como son las tumorales.[2]
Contenido
Trasfondo
Las moléculas de anticuerpo humanas (y los receptores de linfocitos B) comprenden cadenas ligeras y pesadas que contienen regiones tanto constantes (C) como variables (V) que se codifican mediante tres tipos de genes.
- Gen de la cadena pesada – localizado en el cromosoma 14
- Gen kappa (κ)de la cadena ligera – localizado en el cromosoma 2
- Gen lambda (λ)de la cadena ligera – localizado en el cromosoma 22
Múltiples genes de las regiones variables están codificados en el genoma humano de forma que contienen tres tipos distintos de segmentos. Por ejemplo, la región de inmunoglobulina de la cadena pesada contiene 65 genes variables (V), además de 27 genes "Diversity" —diversidad— (D) y seis genes "functional joining" —unión funcional—(J).[3] Las cadenas ligeras también poseen numerosos genes V y J, pero no D. Por este mecanismo de reorganización del ADN de estos genes regionales es posible generar un repertorio de anticuerpos de más de 107 posibles combinaciones. (65 x 27 x 6 = 10530, que se ha de multiplicar por tres órdenes de magnitud si se tiene en cuenta las combinaciones de las cadenas ligeras)
La mayoría de los receptores de linfocitos T están compuestos por cadenas alfa y beta. Sus genes son similares a los de las inmunoglobulinas en el sentido de que comtienen mútiples genes V, D y J en sus cadenas beta (genes V y J en sus cadenas alfa) que se reorganizan durante el desarrollo del linfocito para dotar a la célula con un único receptor de antígeno.
Recombinación V(D)J de las inmunoglobulinas
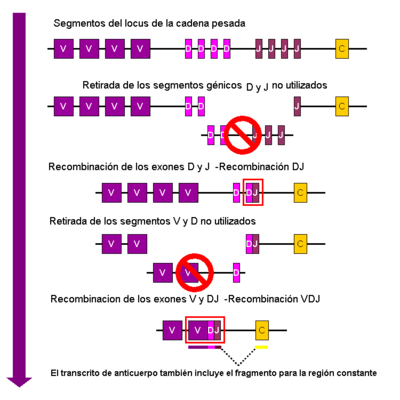 Esquema simplificado de la recombinación V(D)J de las cadenas de inmunoglobulina.
Esquema simplificado de la recombinación V(D)J de las cadenas de inmunoglobulina.
En los linfocitos B en desarrollo, el primer evento de recombinación tiene lugar entre un gen del segmento D y otro del segmento J del locus de la cadena pesada. Se elimina el fragmento de ADN entre los segmentos seleccionados. Tras esta recombinación D-J se produce la unión con el gen V, a partir de una región "corriente arriba" del recientemente formado complejo DJ, para dar lugar a un gen VDJ reorganizado. En ese momento también se elimina del genoma de la célula cualquier otro gen que estuviera comprendido entre los segmentos V y D. Se genera un transcrito primario (ADN sin splicing)que contiene la región VDJ de la cadena pesada, además de las cadenas constantes μ y δ (Cμ and Cδ). (es decir, este transcrito contiene los segmentos V-D-J-Cμ-Cδ). El ARN primario se procesa añadiendo una cola de poliadenilación (poly-A) después de la cadena Cμ y eliminando la secuencia entre el segmento VDJ y las zonas constantes. La traducción de este mARN produce la cadena pesada de la proteína Ig μ.
Las cadenas kappa (κ) y lambda (λ) de los loci de la cadena ligera de la inmunoglobulina se reorganizan de un modo muy similar, excepto por que las cadenas ligeras carecen de segmento D. En otras palabras, el primer paso de la recombinación en las cadenas ligeras implican la unión de las cadenas V y J para dar lugar un complejo VJ tras la adición de la cadena constante durante la transcripción primaria. La traducción del mARN procesado de las cadenas kappa o lambda da lugar a la formación de una proteína de cadena ligera Ig κ o Ig λ.
El ensamblaje de la cadena pesada Ig μ y una de las cadenas ligeras da lugar a la producción de la forma unida a membrana de la inmunoglobulina IgM que se expresa en la superficie de los linfocitos B inmaduros.
Recombinación V(D)J del receptor de linfocitos T
Durante el desarrollo de los linfocitos T, las cadenas de su receptor (TCR) sufren esencialmente la misma secuencia ordenada de sucesos que se ha descrito en el caso de las inmunoglobulinas. La recombinación de D-a-J tiene lugar primero en la cadena β del TCR. Este proceso implica la unión de un segmento de gen Dβ1 a uno de los seis Jβ2 segmentos. Tras esto se continúa con reordenaciones del tipo Vβ con DβJβ. Todos los genes entre Vβ-Dβ-Jβ dentro del complejo de nueva formación se suprimen y el transcrito primario se sintetiza incorporando la porción del dominio constante (Vβ-Dβ-Jβ-Cβ). La traducción genera la producción de la cadena del TCR Cβ completa.
Tras esto se produce el reordenamiento de la otra cadena (α), que comienza con una unión V-J como la descrita en las cadenas ligeras de las Ig. El ensamblaje de de las cadenas β y α produce el TCR- αβ que se expresa en la mayor parte de los linfocitos T.
Mecanismo de recombinación
Secuencias señal de recombinación
Los genes regionales (V, D, J) se encuentran flanqueados por secuencias señal de recombinación (RSSs, por sus siglas en inglés) que son reconocidas por un grupo de enzimas conocidas colectivamente como VDJ recombinasa. Estas RSSs están compuestas por siete nucleótidos conservados (un heptámero) que reside cerca del gen que codifica la secuencia seguida por un espaciador (que contiene entre 12 y 23 nucleótidos no conservados) seguidos por un nonámero conservado (9 pares de bases). Las RSSs están presentes en el lado 3' (secuencia abajo) de una región V y en el lado 5' (secuencia arriba) de la región J. Estos son los lados que están implicados en el empalme. Solo se recombinan eficientemente un par de RSSs espaciadoras que no sean idénticas (p.ej. una con un espaciador de 12 nucleótidos se recombinará con otra con un espaciador de 23 nucleótidos). Esto se conoce como la regla del 12/23 de la recombinación.
Recombinasa VDJ
La recombinasa VDJ es una colección de enzimas, algunas de las cuales son específicas de linfocitos, mientras que otras se expresan en muchos tipos celulares. Los pasos iniciales de la recombinación VDJ se lleva a cabo por enzimas críticas específicas de linfocitos, llamados recombination activating gene-1 y -2 (RAG1 y RAG2). Estas enzimas se asocian unas con otras para reconocer las secuencias RSS e inducir una escisión del ADN en ellas. Esto solo tiene lugar en una de las cadenas de ADN, lo cual conduce a un ataque por nucleótido y la creación de un motivo estructural en bucle.
Otras enzimas de recombinasa VDJ se expresan en muchos tipos celulares y están implicadas en la reparación del ADN que sigue a la actividad de RAG1 y RAG2. una de estas enzimas se llama complejo proteín quinasa activada por ADN (DNA-PK) que repara el ADN de doble cadena. Este complejo se une a cada uno de los extremos del ADN escindido, y recluta varias proteínas más, incluida la nucleasa Artemis, XRCC4 (X-ray repair cross-complementing factor 4), ADN ligasa IV, Cernunnos (también llamado XLF o factor semejante a XRCC4), y cualquiera de las varias ADN polimerasas. Los complejos DNA-PK de cada extremo del aDN se fosforilan mutuamente, lo que resulta en una activación de Artemis. Artemis entonces rompe el bucle que se formó por las proteínas RAG.[4] XRCC4 y Cernunnos actúan concertadamente con DNA-PK para alinear los dos extremos de ADN entre si, y también contribuyen a reclutar una transferasa terminal, que añade nucleótidos al azar a sus extremos. Las ADN polimerasas λ y μ insertan nucleótidos adicionales según se necesiten para crear dos extremos compatibles para la unión. Finalmente la ligasa IV une las cadenas, concluyendo el proceso.[5]
Dada la variabilidad de la posición exacta de la escisión del bucle por Artemis, así como la adición al azar de nucleótidos por la transferasa terminal, la secuencia final de ADN y con ello el anticuerpo resultante es altamente variable, incluso si entre dos anticuerpos se diera la casualidad de que se unieran al azar los mismos segmentos V, D y J. Esta gran diversidad permite a la recombinación VDJ generar anticuerpos incluso para microbios que ni un organismo ni sus antepasados se hayan encontrado nunca.
Enfermedades asociadas con defectos en recombinación V(D)J
El Síndrome de Omenn es asociado con mutaciones en los genes que son necesarios para la síntesis de RAG1 y RAG2. La condición resulta en una inmunodeficiencia combinada severa.[6]
Deficiencias de Artemis, Cernunnos, y ADN ligasa IV también han sido documentado. Estas dolencias fueron asociadas con inmunodeficiencias combinadas severas, y también estas deficiencias de unas enzimas importantes en la reparación de ADN pueden causar una sensibilidad a la radiación ionizante.[6]
Referencias
- ↑ Janeway CA, Jr. et al (2005). Immunobiology. (6th ed. edición). Garland Science. ISBN 0-443-07310-4.
- ↑ Abbas AK and Lichtman AH (2003). Cellular and Molecular Immunology (5th ed. edición). Saunders, Philadelphia. ISBN 0-7216-0008-5.
- ↑ Li A, Rue M, Zhou J, et al (June 2004). «Utilization of Ig heavy chain variable, diversity, and joining gene segments in children with B-lineage acute lymphoblastic leukemia: implications for the mechanisms of VDJ recombination and for pathogenesis». Blood 103 (12): pp. 4602–9. doi:. PMID 15010366. http://www.bloodjournal.org/cgi/pmidlookup?view=long&pmid=15010366.
- ↑ Y. Ma, U. Pannicke, K. Schwarz and M.R. Lieber (2004). «Hairpin opening and overhang processing by an Artemis/DNA-dependent protein kinase complex in nonhomologous end joining and V(D)J recombination». Cell 108: pp. 781-794. doi:. PMID 11955432.
- ↑ D.C. van Gent and M. van der Burg (2007). «Non-homologous end-joining, a sticky affair». Oncogene 26: pp. 7731-7740. doi:. PMID 18066085.
- ↑ a b Raif Geha and Fred Rosen (2008). Case Studies in Immunobiology (5th ed. edición). Garland Science. ISBN 0-8153-4145-8.
Bibliografía
- Hartwell LH, Hood L, Goldberg ML, Reynolds AE, Silver LM, Veres RC (2000). Chapter 24, Evolution at the molecular level. In: Genetics. New York: McGraw-Hill. pp. 805-807. ISBN 0072995874.
Categoría:- Anticuerpos

Wikimedia foundation. 2010.