- Cattleya lueddemanniana
-
Cattleya lueddemanniana
?Cattleya lueddemanniana
Cattleya lueddemanniaEstado de conservación 
IUCN 3.1Clasificación científica Reino: Plantae División: Magnoliophyta Clase: Liliopsida Orden: Asparagales Familia: Orchidaceae Subfamilia: Orchidoideae Tribu: Diuridiae Subtribu: Cattleyae Género: Cattleya
R.Br., 1810Especie: Cattleya lueddemanniana
Rchb. f. 1854Sinonimia - Cattleya dawsonii R. Warner (1862)
- Cattleya roezlii Rchb.f. (1882)
- Cattleya malouana Linden (1885)
- Cattleya bassetii H.J. Veitch (1887)
La Cattleya lueddemanniana es quizás uno de los miembros más representativos de la familia de las orquidáceas, familia perteneciente a las monocotiledóneas, de donde gran parte de las más conocidas especies de valor florístico-ornamental provienen. Para ser precisos la Cattleya lueddemanniana comparte parentesco con la subtribu Laeliinae (sin. Cattleyae), una de las divisiones de la familia de las orquídeas con más representación de especies con alto valor comercial, dentro de ellas comparte un parentesco especialmente cercano con el resto de las especies del subgénero Cattleya, división del género Cattleya donde se encuentran todas las especies de Cattleyas unifoliadas que son más importantes a nivel ornamental dentro de la ya citada subtribu.
En Venezuela, su país de origen, es quizás la orquídea con mayor representación en exposiciones y exhibiciones de orquídeas por igual, así como una de las orquídeas reproducidas artificialmente con más éxito a nivel de viveros comerciales del país.
Como todas las Cattleyas unifoliadas, la Cattleya lueddemanniana es de crecimiento simpodial. Los pseudobulbos más nuevos normalmente desarrollan una sola hoja y una espata simple, la cual contendrá las flores en formación. La floración normalmente ocurre entre septiembre a marzo con un pico en enero-febrero, sin embargo en cultivo no es inusual que la especie florezca una segunda o hasta una tercera vez al año en contadas ocasiones. La especie produce racimos normalmente a partir de espata verde en nuevos pseudobulbos. Las flores, que duran un mínimo de dos semanas, se producen en racimos de 2 a 4 flores, raramente 5, normalmente son de tamaño grande y muy bien formadas si tomamos en cuenta los criterios de exposición. La especie en cultivo no posee un reposo marcado y a veces ni siquiera muestra signos de reposo evidente, sin embargo en la naturaleza los periodos de reposo son más marcados y evidentes. Es una especie tolerante, que muy bien se adapta a condiciones mucho más temperadas a las que encontramos en su ambiente de tierra cálida original.
Sus flores están entre las flores de Cattleyas unifoliadas más fáciles de distinguir, tanto de entre otras especies emparentadas, como de posibles híbridos naturales o artificiales. Las flores tienden a poseer labios en forma de un disco aplanado con bordes ondulados evidentes, aunque no tan evidentemente ondulados como en otras especies de Cattleyas unifoliadas. Todas las Cattleya lueddemanniana puras poseen una columna que es completamente cubierta por el labelo de una manera particularmente distintiva que produce como resultado los dos lóbulos laterales bien formados en los cuales reposan las características manchas oculares amarillas de la especie, estas manchas están presentes en la gran mayoría de las flores, con muy pocas excepciones en flores cuya mancha púrpura-amatista del labelo las enmascare. Igualmente, todas las flores Cattleya lueddemanniana a excepción de las albinas poseen líneas púrpura-amatista que corren sin interrupción desde la base del labelo hasta la mancha frontal del mismo, en las muy pocas plantas donde las rayas se interrumpen (entre 2 a 3%), esta interrupción no supera el centímetro, máximo 2, interrupciones mayores a esto posiblemente indiquen características híbridas en la flor. La columna de la flor posee otro elemento muy distintivo, esta posee una antera en forma de doble asta completamente desarrollada, que cuando es comparada con la antera de apariencia de “uña” o “gancho” de las demás Cattleyas unifoliadas se torna evidente. Muy pocas Cattleyas lueddemanniana poseen astas parcialmente desarrolladas (no más de 2 o 3%), la mayoría de ellas de hecho normalmente son consideradas candidatos a posibles híbridos naturales (con Cattleya mossiae) o artificiales (normalmente con Cattleya warscewiczii).
La especie posee Pseudobulbos acanalados con dos a tres entrenudos, cada pseudobulbo maduro mide aproximadamente unos 25 centímetros, los pseudobulbos tienden a ser relativamente estrechos o aplanados, las hojas son algo más coráceas que en el resto de las Cattleyas unifoliadas, y las hojas nuevas tienden a presentar pequeñas manchas de color vino-tinto, así como también pigmentación rojiza en las plantas expuestas a mayor luminosidad.
Respecto a su reproducción, a diferencia de muchas orquídeas que usan diversos mecanismos de atracción sin recompenza de néctar, la Cattleya lueddemanniana si recompenza con néctar a los potenciales polinizadores. La polinización se cree es mayormente realizada por abejas del neotrópico pertenecientes a la tribu Euglossini, dentro de esa tribu, todos los géneros son potenciales polinizadores siempre que posean la anatomía correcta, pero posiblemente sea mayormente polinizada por abejas del genero Eulaema. Las flores atraen tanto a machos como hembras, y por lo que se ve en cultivo, son capaces de atraer y ser polinizadas por un mayor número de especies a las ya citadas, incluyendo especies foráneas naturalizadas en Venezuela como por ejemplo las abejas melíferas.
Contenido
Hábitat




Hábitat de la Cordillera de la Costa
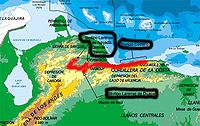
Crece en la vertiente norte de la cordillera de la Costa, en las laderas que dan al mar Caribe al norte de los estados de Carabobo, Aragua, Vargas y en el Distrito Federal. Normalmente crece en zonas calientes y con poca precipitación anual promedio (400-800mm con zonas circundantes entre 800-1200mm), casi exclusivamente como epifita con preferencia a bosques semi-xerófitos a xerófitos, pero también se encuentra en bosques de tierra caliente más húmedos y protegidos por vegetación, bosques de galería, tanto los que recorren pequeños ríos estaciónales, como aquellos bosques que crecen protegidos por las oquedades naturales que se forman en los pliegues de las colinas. Crece desde el nivel del mar, colindante a la línea costera, hasta aproximadamente los 900 msnm. La especie normalmente se consideraba como abundante hasta el litoral central del estado Vargas, en los alrededores de Cabo blanco, en Maiquetía (Ploch, 1996), estando presumiblemente ausente desde Cabo Blanco a la Región de Barlovento/Higüerote. Las zonas de recolección mayormente citadas ocurren comprendidas en la zona occidental del estado Vargas (alrededores de Chichiriviche/Pto. Cruz), Distrito Federal (quebrada de Tacagua) y el litoral del estado Aragua (zona de Choroní/El Limón).
Debido a la diversidad de estos bosques de tierra caliente, la especie forma colonias creciendo bien iluminada en árboles de portes de diverso tamaño (zonas más húmedas), hasta muy expuesta o directamente expuesta en árboles de porte medio, arbustos, matorral xerófito y hasta cardonales (zonas menos húmedas), muchas veces forma colonias en terrenos muy escarpados, especialmente cuando las paredes arboladas de estas zonas escarpadas ayudan a "acumular" mayor humedad. La temporada de sequía en esta zona dura unos 3-4 meses. Fuera de las áreas de recolección citadas en el norte de Aragua, este de Vargas y valle de tacagua en el Distrito federal, se presume que la especie continúa de forma ininterrumpida por el norte de Carabobo hasta alcanzar el norte de la depresión de Yaracuy (zona "costera" del estado Yaracuy), este último lugar se cita es donde las poblaciones de la costa se unen con las poblaciones occidentales presentes en la serranía de Aroa, sin embargo el autor no pudo confirmar la presencia de la especie en la depresión de Yaracuy, y desconoce hasta qué tan al oeste ha sido avistada la especie en en la costa de Carabobo.
Hábitat de la Serranía de Aroa y tierras altas de Lara
La especie también habita más al occidente, en las faldas norteñas de la serranía de Aroa en donde se encuentra desde el área comprendida entre San Felipe y Aroa, hasta adentrarse siguiendo esta misma serranía hasta el Estado Lara, en los alrededores de Duaca. Entre los poblados de Duaca y Aroa, colindante a con ellos y al oeste de ellos, se encuentra otra área paralela de tierras altas donde también habita la especie en la frontera Yaracuy-Lara. Finalmente encuentra su punto más occidental de dispersión ya bien adentrada en el estado Lara, desde los Alrededores de la sierra de Bobare en las tierras altas al Norte de Barquisimeto que rodean al río tuy, hasta la región colindante de Aguada grande la cual es cortada en su lado occidental y norte por la depresión que forma el río Tocuyo. Al oeste y norte del río tocuyo también se encuentran serranías, sin embargo aunque citada para la zona, el autor no tiene conocimiento de que exista la especie en dicha área (a excepción de la serranía entre el Río Baragua y tocuyo por los alrededores del cerro Guacamuco frente a Siquisique). Fuera de estas áreas, al parecer los avistamientos de la especie en zonas de serranías colindantes son raros, todos presumiblemente acaecidos décadas atrás, y el autor además no pudo encontrar evidencia de ejemplares en cultivo que se identifiquen como recolectados en el área. Es probable que en estas últimas zonas la especie de estar realmente presente solamente esté representada por colonias dispersas, situación que amerita una mayor clarificación.
La ecología para Yaracuy-Lara es equivalente a la ecología de la costa exceptuando algunos detalles, en algunas zonas especialmente de Lara la diferencia de temperaturas día-noche puede ser extrema, alcanzando en el día los alrededores de los 40 ºC, lo que hace que la evaporación no sea compensada por la precipitación. Por otro lado, los bosques serranos de tierra caliente donde habita la especie en Yaracuy poseen índices pluviométricos más altos que en la zona costera (1200-1600mm), y los extremos de temperatura señalados con anterioridad no son completamente aplicables. La especie en esta zona habita a mayor altura que en la costa, aproximadamente entre los 400 y 900 msnm. En esta zona también se han reportado ocasionales colonias de introgresión con la Cattleya mossiae, que aunque presentes, deben ser consideradas como inusuales (Yaracuy).
Hábitat de la Serranía de San Luís
También se cita la posible presencia de la especie en la serranía de San Luís (Estado Falcón), un área de serranías tanto más al norte de las serranías ya descritas en Lara-Yaracuy. Dentro de esta área se cita específicamente a los alrededores de Curimagua aunque a menor altura. Esto ha sido documentado en alguna literatura y por posibles avistamientos que incluyen descripciones de las plantas de la zona que han llegado a ser del conocimiento del autor de este artículo.
Aunque el autor no descarta esta área para el presente análisis de esta especie, considera que lo más apropiado sería afirmar que la Serranía de San Luís deba considerarse una mera posibilidad. El autor no conoce evidencia concreta sobre la presencia en la zona, no ha podido encontrar ejemplares en cultivo provenientes de dicha zona, y según información anecdótica recabada de los supuestos avistamientos, de efectivamente la especie estar presente en la zona, ésta sería muy inusual, quizás hasta restringida a una sola colonia de modesto tamaño.
Es de hacer destacar que reputados orquideólogos Venezolanos como Carlos García Esquivel, persona que ha investigado a fondo la Serranía de San Luís en busca de nuevas especies de orquídeas para la ciencia, jamás se ha topado con la especie en esa área.
Extrapolación del hábitat a el cultivo
En cutivo la especie es sumamente adaptable a regímenes de cultivo diversos y no siempre equivalentes a los que la especie soporta en su ambiente natural (mayor humedad, ausencia total de período de reposo, regímenes de iluminación más sombreados, diferencias de temperatura día-noche menos marcados y temperaturas promedio anuales tanto más frías).
Variantes regionales
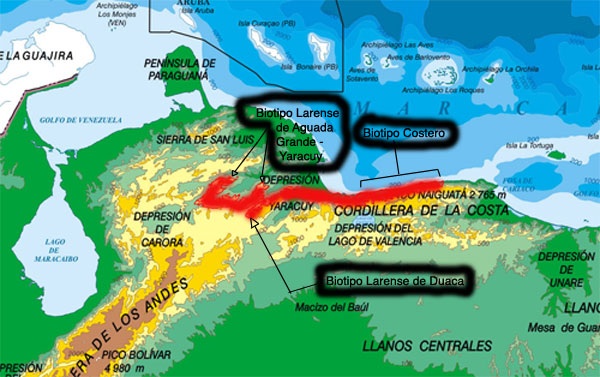
La especie muestra un grado relativamente notorio de regionalismo entre poblaciones, lo que se podría adscribir a ecotipos, biotipos o demes, lo que depende de si estas variaciones son vistas como adaptaciones a la ecología, adaptaciones a la ecología que también responden a un cline geográfico (variación gradual), o que simplemente responden a que las frecuencias genéticas ligadas al fenotipo, que a pesar del intercambio genético, pueden variar en sus frecuencias relativas entre poblaciones o colonias distintas, incluso si ellas son geográficamente colindantes.Para realizar esta descripción y por mera comodidad, utilizaremos la palabra “biotipo”, la cual es la más usada entre cultivadores de esta especie, haciendo la acotación que a pesar de las distintas tendencias entre "biotipos" diferentes, el solape entre fenotipos es usual, los fenotipos descritos son básicamente tendencias y así serán reflejadas en las descripciones aquí realizadas, por lo que postular "subespecies" dentro de la Cattleya lueddemanniana es según el entender del autor incorrecto.

Biotipo Costero:''Las poblaciones de la cordillera de la costa tienden a tener flores grandes, con pétalos normalmente anchos y labelo generalmente amplio y ancho. La forma general es usualmente buena, pero carece de substancia. Una característica quizás la más notoria de este biotipo, es que no se conocen ejemplares de tépalos realmente oscuros, siendo una característica de las costeras el que sea muy común que al abrir den la apariencia de semi-albas solamente para ecentuar el color al par de días de abiertas, lo que resulta en flores de fenotipo promedio suave. Las manchas oculares tienden normalmente a ser de color claro, aunque no se descartan manchas oculares amarillas mucho más oscuras. La mancha del labelo es variable en tamaño y tiende a presentarse desde moteados no muy densos, hasta glutinarse en patrones muy compactos (casi sólidos) que comprometen áreas variables del labelo. Labios uniformemente sólidos sin trazas de moteados son más bien poco usuales.

Biotipo Larense (área de Aguada Grande a Aroa)'': entre el municipio urdaneta hasta la serranía entre Aroa y San Felipe, la especie tiende a presentar flores normalmente de tamaño mediano, usualmente de segmentos relativamente estrechos, en colores con tendencia a los tonos oscuros (roseas hasta algunas rubras) y con un disco del labelo que a veces tiende a ser estrecho en relación al largo del tubo. La mancha del labelo tiende a ser profusamente moteada aunque no tan compacta como en muchas costeras, algunas veces con una base de color más oscuro que "cubre" parcialmente al moteado resaltando el color general del labelo, igualmente no se descartan labelos sólidos, lo que es lo menos usual. Las manchas oculares tienden a ser oscuras, aunque no se descartan manchas oculares más claras. En términos reales este "biotipo" es muy variable, en tamaño, forma como color general de las flores.

Biotipo Larense (área de Duaca)'': En las serranías colindantes al área de Duaca la especie posee flores en tonos claros a medios (Suaves a Roseas), en líneas generales parecidas a las costeras. Labelo generalmente moteado a profusamente moteado aunque usualmente no tan denso y compacto como en muchas costeras, raramente poseen labelos sólidos. Buena forma general. Manchas oculares de intensidad variable. Es probable que otras poblaciones geográficamente representativas dentro del área "larense" posean un fenotipo más acorde al aquí descrito.
Biotipo Larense (área de Curimagua, Falcón)'': El autor no ha podido confirmar la presencia de la especie en esa zona, sin embargo por información recolectada, las plantas de esta área podrían ser similares a lo que aquí descrito para la zona alrededor de Duaca.
Es de hacer notar que en cultivo solamente se reconocen 2 “biotipos”, no haciéndose distinción alguna entre las dos áreas “larenses” que poseen cada una particularidades específicas, lo que ha generado una amplia confusión en lo que respecta al biotipo costero, ya que este último ha sido muchas veces confundido con lo que aquí llamamos “Biotipo Larense de Duaca”, que aunque similares, no pertenecen a la misma población. Respecto a esto debemos destacar que en términos de su contribución genética en plantas cultivadas, los genes de Lara-Yaracuy representan la totalidad en plantas albas, es la norma en las semialbas, y constituye la mayor parte del bagaje genético de las plantas color. El único caso en donde los genes costeros predominan, y en este caso en la totalidad de sus plantas, es en las plantas coeruleas. Por esta misma razón de relativa predominancia de plantas “Larenses” normalmente se dice que el pico de floración de la Cattleya lueddemanniana se encuentra entre enero y febrero, sin embargo ambas zonas geográficas poseen un comportamiento de floración diferenciado, siendo las costeras normalmente las primeras en florecer (de septiembre a febrero con un pico en noviembre-diciembre), seguidas por las “Larenses” (de diciembre a marzo con un pico en enero-febrero), sin embargo esto no es tan evidente excepto si se manejan numerosas plantas en cultivo, la especie como ya se dijo es capaz de florecer en cultivo más de una vez al año, y incluso un cultivar específico por condiciones de cultivo puede variar considerablemente de un año a otro su "época de floración" usual.
Estatus de Conservación de la Especie
Según Foldats (2003), la especie debería estar clasificada como en peligro de extinción con una recomendación de que la especie pase del apéndice II a el apéndice I de CITES para garantizar el comercio internacional de plantas propagadas en viveros comerciales, sin embargo, a pesar de los esfuerzos de los cultivadores comerciales por suplir la demanda exitosamente de manera artificial, aún es común ver los remanentes poblacionales siendo comúnmente atacados por aficionados inescrupulosos en búsqueda de plantas gratuitas para sus colecciones, lo que aunado a la extirpación de dichos remanentes por parte de campesinos de la zona que buscan vender las plantas al borde de las carreteras, y la práctica barbárica de la agricultura de tala y quema, hacen que esta especie muy bien pueda ser considerada en peligro crítico debido a su tasa de extirpación. Los dos biotipos, así como su área de transición, actualmente sobreviven bajo poblaciones remanentes que deberían considerarse como altamente fragmentadas entre ellas y dentro de ellas, lo que seguramente afecta un intercambio genético que garantice la salud de la especie.
Variedades de valor botánico en cultivo




La especie es ampliamente variable, tan variable como variable es considerada otras especies de Cattleyas unifoliadas, sin embargo, y como igualmente pasa con otras especies de Cattleya, la variación conocida aunque refleja poblaciones en cultivo, muchas de ellas son raras a extremadamente raras en poblaciones naturales, siendo solamente las clasificadas como “color” (especialmente en sus tonos claros a medios con labelos moteados), las que realmente reflejan el denominador común a esperar en poblaciones naturales, independientemente del biotipo al que pertenezcan.
Clasificación a partir del color base de la flor
En la especie lo normal es que los segmentos de la flor posean una coloración uniforme o bastante uniforme, sin embargo se conocen algunos casos donde los pétalos son evidentemente más oscuros que los sépalos, lo que normalmente no se clasifica de manera alguna y de hecho para juzgamientos es una característica más bien penalizada. En el caso de las flores uniformes o sin diferencia muy notoria entre segmentos, lo que como ya se dijo constituye la virtual mayoría de las flores, estas podrían definirse como rosa-lila uniforme, y en el caso de la C. lueddemanniana usualmente no se somete a clasificación alguna excepto definiéndolas a todas como “Flores Color” o “flores tipo”. Sin embargo la variación del color base puede ser grande, desde flores casi blancas, hasta llegar a flores lilas muy oscuras con infusiones rojizas, el espectro completo de gradación en intensidad del color podría definirse como suavissima, suave, rosea y rubra, pero de esta sub-clasificación solamente se usan los extremos, a saber las suavissimas para distinguirlas de las semialbas verdaderas, y las rubras, para resaltar esta característica llamativa y inusual de la flor.
También existen flores en apariencia albinas, es decir, que no llegan a sintetizar los pigmentos de manera completa o al menos evidente para el ojo humano, este caso de albinismo aparente se conoce como albescens. Otras variaciones posibles son las flores realmente blancas, estas pueden ser completamente blancas, lo que es un caso de albinismo total clasificado en esta especie como “alba”, o solamente serlas en su color base, lo cual no compromete el color del labelo y representa un caso de albinismo parcial clasificado en esta especie como “semi-alba”.
También existen flores que en vez de ser rosa-lilas o blancas son lila azuladas, normalmente muy tenues y de color lila-azulado fuerte en el labelo, estas flores suavissimas por su coloración inusual azulada son clasificadas como “coerulea”, un tono que puede llegar a ser también poco intenso en el labelo, lo que más bien le da una apariencia grisácea a la flor (caerulescens).
También existen flores de un color muy sutil, al punto que solamente puede ser observado de manera más notoria en el color sobrepuesto y no en su color base, las así llamadas “delicatas”, las cuales tienen un color que se puede definir como rosa algo desvaído en el labelo y puede ocurrir tanto en plantas semialbas como suavissimas.
Clasificación a partir del patrón del color sobrepuesto de la flor:La inmensa mayoría de las lueddemannianas poseen un patrón estriado en la garganta del labelo que cuando llega a su parte frontal se convierte en un patrón visual parecido a estrías compuestas por motas, el cual puede ocurrir de manera muy localizada, o en todo el labelo (lo que es más usual), hasta comprometer en algunos casos parte de la zona exterior del tubo. Este moteado también puede variar mucho en densidad, desde flores con apenas unas cuantas “motas” esparcidas en el labelo, hasta flores con un moteado tan intenso y compacto que dan la apariencia de ser sólidas. No existe clasificación alguna para describir esta variabilidad en la especie, exceptuando en las más compactas, las cuales son muchas veces incluidas entre los “labios sólidos” o “solid lip” (ver siguiente párrafo).
La especie aunque de manera más inusual, también puede presentar labios uniformemente sólidos, los cuales pueden comprometer parte del labelo, cubrirlo por entero, hasta desparramarse parcial o totalmente por la parte externa del tubo. Para otras especies emparentadas esto sería clasificado por el orden de la parte del labelo comprometida como “maculata”, “orlata” y “atro”, sin embargo en la C. lueddemanniana este fenotipo simplemente se le caracteriza por los nombres “labio sólido” o su anglicismo “Solid Lip”.
A veces el color presente en el labelo puede estar presente en pétalos y hasta en sépalos, cosa que ocurre normalmente en plantas de labio sólido aunque también puede ocurrir en las moteadas, especialmente si son profusamente moteadas. Cuando esto ocurre en esta especie se lo clasifica como “aquinii”, y el fenotipo resultante de una aquinii de hecho puede variar un poco, este se puede presentar como una orla algo más oscura en el borde de los pétalos, presentarse como una mancha sólida en la punta de los mismos (lo más deseado en cultivo), o presentarse como un variegado (pincelado) en sépalos y pétalos. Flores aquinii pueden ocurrir tanto en flores color, como semialbas y coeruleas.
Otro caso particular ocurre ocasionalmente en flores que no tienen color en el labelo excepto por el color base, razón por la cual son clasificadas como concolor. En el caso de esta especie las estrías de la garganta están presentes a pesar de estar ausente patrón alguno de color en la parte frontal del labelo, y como muchas concolor en cultivo son descendientes de flores albinas, es posible que este patrón se explique como un caso de albinismo parcial análogo a el caso de las semialbas, solamente que en este caso compromete únicamente el color sobrepuesto en vez de el color base de la flor.
Variación en la coloración del labelo:La gran mayoría de las flores poseen una mancha del labelo del mismo color rosa-lila de pétalos y sépalos pero mucho más saturado y intenso, pudiendo variar de planta en planta entre un color más rojizo, y en otras hacia un morado más oscuro. Esta variación no se refleja en la clasificación como si se refleja en otras Cattleyas en donde sus extremos en gradación son clasificados como “vinicolor” y “purpúreo”.
Otro factor de variación en el color del labelo se tiene en el color base del labelo, este normalmente es igual en gradación al de los pétalos y sépalos, sin embargo en algunas flores puede ser más oscuro, hasta llegar a un rojizo intenso, último caso el cual el moteado queda camuflado entre el color base hasta el punto en que solamente una inspección cercana a la flor denotará la presencia del moteado. Esta característica de que el labelo “ayude” en algunos casos a resaltar el patrón de color sobrepuesto en el caso de la Cattleya lueddemanniana no es sujeto a clasificación varietal alguna.
Variación en las manchas oculares:En cultivo no se usa una clasificación para las variaciones en las manchas oculares, exceptuando por dos casos, el término “cegata”, término bastante inusual que a veces es usado en plantas con manchas oculares muy poco evidentes, tenues a parcialmente cubiertas por el color oscuro del labelo, y el término “alba plena”, que a veces es usado en flores albinas con manchas oculares de similares características.
Las manchas oculares en la Cattleya lueddemanniana normalmente están bien definidas como dos óvalos ubicados en los lóbulos laterales del labelo. Su tamaño va desde óvalos apenas sugeridos, pasando por lo más usual, óvalos de tamaño medio, hasta en contadas ocasiones desparramarse afuera de los lóbulos laterales hacia los lados del labelo hasta llegar próximo a su base.
Respecto a el color de las manchas oculares, ellas normalmente son de un amarillo evidente bordeado por una pequeña orla blanca, sin embargo el “amarrillo” puede variar en intensidad, desde un color crema tenue, pasando por crema amarillento, hasta alcanzar un amarillo encendido. El amarillo de la garganta puede variar también en intensidad, lo cual no necesariamente depende el tono del amarillo presente en las manchas oculares, sin embargo esto no es evidente en muchos casos por ser este amarillo enmascarado por las líneas púrpura de la garganta del labelo, por lo que este detalle es más fácil de visualizar en las albas, en donde en contados casos virtualmente ambas manchas oculares se fusionan a través de un amarillo igualmente intenso en la garganta del labelo.
Lasmanchas oculares en las pocas circunstancias donde se encuentra con el color oscuro del labelo, este último usualmente termina circundándolas sin enmascararlas, sin embargo en algunos casos lejos de hacer esto, las solapa parcialmente, lo que normalmente ocurre sobreponiéndose al amarillo y cubriéndolo, y raramente ocurre mezclándose parcialmente con el amarillo en leves tonos rojizos.
Bibliografía
- Aulisi C. (1997). Cattleya lueddemanniana y sus Variedades. Editorial Torino.
- Aulisi, C. & Foldats, E. (1989). Monography of the Venezuelan Cattleyas and its Verieties. Editorial Torino.
- Foldats, E. Et Col (2003). Libro Rojo de la Flora Venezolana. Litografía Imagen Color C.A.
- Mantellini, A. (2008). Selective Reproduction of Cattleya lueddemanniana. Documento de apoyo a la conferencia dictada en la W.O.C. de Miami (U.S.A).
- Pahl,J; Castiglione, G & Vaamonde, R (2007).Some considerations about Venezuelan unifoliate Cattleya. Orchidnews
- Ploch, U. (1996). Cattleya lueddemanniana Rchb. F. Orquideophilo. Vol 4 (Nº2).
Enlaces externos
 Wikimedia Commons alberga contenido multimedia sobre Cattleya lueddemanniana.Commons
Wikimedia Commons alberga contenido multimedia sobre Cattleya lueddemanniana.Commons
Categorías: Especies en peligro de extinción | Cattleya | Flores












Wikimedia foundation. 2010.