- Flor
-
Partes de la flor
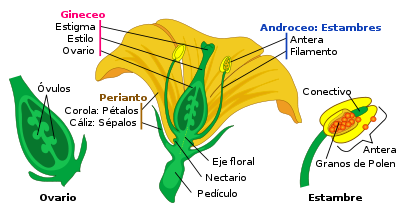 Partes de una flor madura.
Partes de una flor madura.La flor es la estructura reproductiva característica de las plantas llamadas espermatofitas o fanerógamas. La función de una flor es producir semillas a través de la reproducción sexual. Para las plantas, las semillas son la próxima generación, y sirven como el principal medio a través del cual las especies se perpetúan y se propagan.
Todas las espermatofitas poseen flores que producirán semillas, pero la organización interna de la flor es muy diferente en los dos principales grupos de espermatofitas: gimnospermas vivientes y angiospermas.
Las gimnospermas pueden poseer flores que se reúnen en estróbilos, o la misma flor puede ser un estróbilo de hojas fértiles. En cambio una flor típica de angiosperma está compuesta por cuatro tipos de hojas modificadas, tanto estructural como fisiológicamente, para producir y proteger los gametos: sépalos, pétalos, estambres y carpelos.[1]
En las angiospermas la flor da origen, tras la fertilización y por transformación de algunas de sus partes, a un fruto que contiene las semillas.[2]
El grupo de las angiospermas, con más de 250.000 especies, es un linaje evolutivamente exitoso que conforma la mayor parte de la flora terrestre existente. La flor de angiosperma es el carácter definitorio del grupo y es, probablemente, un factor clave en su éxito evolutivo. Es una estructura compleja, cuyo plan organizacional está conservado en casi todos los miembros del grupo, si bien presenta una tremenda diversidad en la morfología y fisiología de todas y cada una de las piezas que la componen. La base genética y adaptativa de tal diversidad está comenzando a comprenderse en profundidad,[3] así como también su origen, que data del Cretácico inferior, y su posterior evolución en estrecha interrelación con los animales que se encargan de transportar los gametos.
Con independencia de los aspectos señalados, la flor es un objeto importante para los seres humanos. A través de la historia y de las diferentes culturas, la flor siempre ha tenido un lugar en las sociedades humanas, ya sea por su belleza intrínseca o por su simbolismo. De hecho, cultivamos especies para que nos provean flores desde hace más de 5.000 años y, actualmente, ese arte se ha transformado en una industria en continua expansión: la floricultura.
Definición
 Diagrama esquemático que muestra las partes de la flor. 1: receptáculo, 2: sépalos, 3: pétalos, 4: estambres, 5: pistilo.
Diagrama esquemático que muestra las partes de la flor. 1: receptáculo, 2: sépalos, 3: pétalos, 4: estambres, 5: pistilo.
La flor es un corto tallo de crecimiento determinado que lleva hojas modificadas estructural y funcionalmente para realizar las funciones de producción de gametos y de protección de los mismos, denominadas antófilos.[a]La plantilla {{Note label}} está obsoleta, véase el nuevo sistema de referencias.[2]
El tallo se caracteriza por un crecimiento indeterminado. En contraste, la flor muestra un crecimiento determinado, ya que su meristema apical cesa de dividirse mitóticamente después de que ha producido todos los antófilos o piezas florales. Las flores más especializadas tienen un período de crecimiento más breve y producen un eje más corto y un número más definido de piezas florales que las flores más primitivas.
La disposición de los antófilos sobre el eje, la presencia o ausencia de una o más piezas florales, el tamaño, la pigmentación y la disposición relativa de las mismas son responsables de la existencia de una gran variedad de tipos de flores. Tal diversidad es particularmente importante en estudios filogenéticos y taxonómicos de las angiospermas. La interpretación evolutiva de los diferentes tipos de flores tiene en cuenta los aspectos de la adaptación de la estructura floral, particularmente aquellos relacionados con la polinización, dispersión del fruto y de la semilla y de la protección contra los predadores de las estructuras reproductivas.[4] [5] [6]
Morfología de las flores: diversidad y tendencias evolutivas
Con más de 250.000 especies, las angiospermas forman un grupo taxonómico evolutivamente exitoso que conforma la mayor parte de la flora terrestre existente. La flor es el carácter definitorio del grupo y es, probablemente, un factor clave en su éxito evolutivo.[3]
La flor está unida al tallo por un eje, denominado pedicelo, que se dilata en su parte superior para formar el receptáculo en el cual se insertan las diversas piezas florales, las cuales son hojas modificadas que están especializadas en las funciones de reproducción y de protección. Desde afuera hacia adentro de una flor típica de angiosperma se encuentran las denominadas piezas estériles, con función de protección, y compuestas por sépalos y pétalos. Por dentro de los pétalos se disponen las denominadas piezas fértiles, con función reproductiva, e integrado por estambres y carpelos. Los carpelos de las angiospermas son, con respecto a los carpelos de sus ancestros, una estructura innovativa y privativa, ya que por primera vez en el linaje, encierran completamente al óvulo, de forma que el polen no cae directamente en el óvulo (como en las gimnospermas) sino en una nueva estructura del óvulo llamada estigma, que recibe al polen y estimula la formación del tubo polínico que llegará al óvulo para producirse la fecundación.[7]
La flor de angiosperma es una estructura compleja cuyo plan organizacional está conservado en casi todas las angiospermas, con la notable excepción de Lacandonia schismatica (Triuridaceae) que presenta los estambres en posición central rodeados de los carpelos.[8] [9] Esta organización tan invariable no indica en modo alguno que la estructura floral es conservada a través de los diferentes linajes de angiospermas. Por el contrario, existe una tremenda diversidad en la morfología y fisiología de todas y cada una de las piezas que componen a la flor, cuya base genética y adaptativa está comenzando a comprenderse en profundidad.[3]
Se ha sugerido que existe una tendencia en la evolución de la arquitectura floral, desde un plan "abierto", en el que las variaciones están determinadas por el número y disposición de las piezas florales, hacia un plan "cerrado", en el cual el número y disposición de las piezas están fijados.[10] En tales estructuras fijas, las elaboraciones evolutivas ulteriores pueden tener lugar a través de la concrescencia, o sea, por medio de la fusión o estrecha conexión de las distintas partes.[11] El plan de organización "abierto" es común en las angiospermas basales y las primeras eudicotiledóneas, mientras que el plan de organización "cerrado" es la regla en el clado de las gunnéridas (o núcleo de las eudicotiledóneas) y en las monocotiledóneas.[12]
Disposición de las piezas florales
 Magnolia grandiflora, una flor espiralada.
Magnolia grandiflora, una flor espiralada.Según la familia considerada, las piezas de la flor se pueden disponer sobre el receptáculo de dos modos diferentes. En el caso de la denominada disposición espiralada, las piezas se insertan consecutivamente y a diferentes niveles, describiendo una espiral sobre el eje del mismo modo en que las hojas se insertan en el tallo. Ejemplo de especies con flores espiraladas son Magnolia grandiflora (magnoliáceas), Victoria cruziana (ninfeáceas) y Opuntia ficus-indica (cactáceas). El segundo tipo de disposición de los antófilos es la denominada disposición verticilada o cíclica, en el que las piezas se insertan en varios nudos del eje, disponiéndose en verticilos o ciclos. Cada pieza floral de un verticilo alterna con las piezas del siguiente, por ejemplo, los pétalos alternan con los sépalos. En estas flores, denominadas cíclicas o verticiladas, el número de verticilos puede variar, dependiendo nuevamente de la familia considerada. Muy frecuentemente las flores llevan cuatro ciclos (llamadas tetracíclicas), como las de Solanum (solanáceas): un ciclo de sépalos, uno de pétalos, otro de estambres y el último de carpelos. También son usuales las flores pentacíclicas (llevan cinco ciclos) ya que, en este caso, presentan dos ciclos de estambres en vez de uno solo, como las flores de Lilium (liliáceas). Hay muchos otros casos, finalmente, en los que las flores presentan varios verticilos de estambres, como en Poncirus trifoliata (rutáceas), en cuyo caso las flores presentan más de cinco ciclos.[13]
En las angiospermas primitivas las flores son relativamente voluminosas y sobre el receptáculo cónico y alargado llevan, en disposición helicoidal o espiralada, numerosas piezas periánticas, estambres y carpelos. En los grupos derivados, más evolucionados, se observa un progresivo empequeñecimiento (reducción) de las flores y una disminución del número de sus piezas (oligomerización). Se ha postulado que la base adaptativa de esta tendencia evolutiva es el desarrollo más rápido y un menor riesgo de daños cuando hay numerosas flores pequeñas que cuando hay pocas flores y frutos grandes y, por otro lado, una mejor integración espacial y formal de los órganos en las flores oligómeras que en las polímeras. En relación con esta reducción y oligomerización se produjo un acortamiento del eje floral o receptáculo, de modo tal que, pasando por estados intermedios helicoide-verticilados, se llegó finalmente a la posición uniformemente verticilada o cíclica de las piezas florales.[14]
Simetría floral
Se dice que en un objeto existe simetría cuando por lo menos un plano puede dividirlo en dos partes, tal que cada parte sea la imagen especular de la otra. Ciertas flores no presentan ningún plano de simetría por lo que se dicen asimétricas o irregulares, como es el caso de las "achiras" (las especies del género Canna). En la mayor parte de las flores verticiladas, no obstante, debido a que existen repeticiones de piezas florales, pueden existir uno o más planos de simetría, por lo que las mismas pueden tener simetría bilateral (es decir, un solo plano de simetría) o simetría radial (o sea, varios planos pueden dividir a la flor en otras tantas imágenes especulares). Así, se distinguen por su simetría, dos tipos de flores. Las flores denominadas actinomorfas, radiadas o polisimétricas presentan simetría radial, como es el caso de Tulipa gesneriana (liliáceas) o Linum usitatissimum (lináceas). En cambio, las flores monosimétricas, dorsiventrales o cigomorfas tienen simetría bilateral y la evolución de su forma tiene relación con la necesidad de atraer y guiar a los polinizadores hasta ellas, como por ejemplo, las flores de las orquídeas y muchas leguminosas.[15] Las flores espiraladas se consideran asimétricas según algunos autores,[15] o actinomórficas según otros.[16] [17]
La simetría se define en general a través del perianto, no obstante, el androceo también puede estar implicado debido a la reducción de algunas piezas (estaminodios) o al aborto de uno o más estambres, como ocurre en las eudicotiledóneas y en las monocotiledóneas. Las flores cigomorfas aparecieron relativamente tarde en el registro fósil (Cretácico superior) en comparación con el período aceptado de diversificación temprana de las angiospermas (Cretácico temprano).[18] La polisimetría, entonces, se considera generalmente el estado ancestral de las angiospermas, y la monosimetría, el estado derivado que ha evolucionado de modo independiente en numerosas ocasiones. Como un carácter arquitectónico derivado, generalmente asociado con la diversificación de muchos linajes de plantas, constituye una innovación morfológica. De hecho, los cambios en la simetría floral están asociados con los cambios en el rango de polinizadores efectivos, lo que, por su lado, puede originar barreras a la hibridación entre las especies. Así, varios de los taxones con mayor riqueza de especies llevan flores monosimétricas, tanto en las eudicotiledóneas centrales (fabáceas, asteráceas) como en las monocotiledóneas (zingiberáceas, orquidáceas). Estos taxones tienen generalmente planes de organización "cerrados" con estructuras florales más o menos elaboradas. En contraste, la monosimetría está casi ausente en las angiospermas basales y es bastante infrecuente en las eudicotiledóneas basales (Ranunculales, Proteales) donde el plan de organización "abierto" es más común que en las eudicotiledóneas nucleares.[19] [18]
Perianto y perigonio
Del mismo modo que las Bennettitales extintas y que sus parientes actuales las Gnetales, las flores de las angiospermas poseen una envoltura floral, un perianto, el cual está constituido por los verticilos estériles de la flor: el cáliz y la corola. Es evidente que la presencia del perianto se halla en relación funcional con la polinización por animales, ya que en estado de pimpollo el perianto protege los órganos reproductores aún inmaduros de los animales herbívoros visitantes y, durante la floración, las partes del perianto vivamente coloreadas contribuyen de modo esencial a la atracción de los animales polinizadores. En las flores de las angiospermas primitivas se observa un perianto homoclamídeo (todas las piezas iguales) formado por numerosas hojas perigoniales dispuestas helicoidalmente y libres entre sí, las exteriores parecidas a brácteas y las internas progresivamente más coloreadas y corolinas, similares a pétalos (por ejemplo en Magnolia).[14]
En muchos casos, en particular en las especies que son anemófilas (aquellas que no utilizan animales para la polinización, sino la acción del viento), como por ejemplo el "sauce llorón" (Salix babylonica, salicáceas) o el "fresno europeo" (Fraxinus excelsior , oleáceas) el perianto puede faltar, es decir, las flores solo presentan los verticilos fértiles (androceo y gineceo) y se denominan aperiantadas, aclamídeas o, simplemente, flores desnudas. Las flores que presentan perianto, el caso más frecuente, se denominan periantadas, clamídeas o vestidas. Las flores desnudas se han formado en el curso de procesos evolutivos de progresivo empequeñecimiento y simplificación de la flor. Tales flores secundariamente simplificadas caracterizan grupos de plantas anemófilas en las que un perianto diferenciado no solo es innecesario, al no producirse la visita de las flores por los animales, sino que incluso llega a ser perjudicial para la dispersión o la captación del polen por los estambres y carpelos, respectivamente.[14]
En las flores periantadas puede darse el caso de que solamente presenten cáliz, por lo que las mismas se denominan monoclamídeas o, más usualmente, apétalas y el ejemplo más conspicuo es el de la familia de las urticáceas. La ausencia de pétalos en estas flores no debe conducir a la suposición de que las mismas no son vistosas, ya que hay casos en los que los sépalos (denominados sépalos petaloideos) adquieren la consistencia, forma y colorido propio de los pétalos, como por ejemplo en las especies del género Clematis (ranunculáceas). Cuando las flores periantadas presentan tanto cáliz como corola se denominan diclamídeas. Los miembros de ambos verticilos pueden ser diferentes entre sí en forma y color y las flores se dicen heteroclamídeas, como por ejemplo la "rosa" (Rosa sp., rosáceas). Cuando, en cambio, las piezas del cáliz y de la corola son indistinguibles entre sí en forma y color, la flor se llama homoclamídea. En este tipo de flores, típicas de muchas familias de monocotiledóneas como las iridáceas y las amarilidáceas, el perianto se denomina perigonio y las piezas que lo componen se llaman tépalos. Si los tépalos se parecen a un pétalo el perigonio se llama corolino (de corola), y si se parecen a sépalos se dice perigonio calicino (de cáliz).[20]
Asimismo, las piezas del perianto pueden ser independientes (cáliz dialisépalo y corola dialipétala) o estar más o menos soldados entre sí por sus bordes (cáliz gamosépalo y corola gamopétala, ver más adelante) o con otras piezas de la flor. Esta frecuente fusión de las piezas de un mismo ciclo se denomina concrescencia de las piezas del perianto y se produce en relación directa con la especialización en la zoogamia. Así, las flores con las piezas del perigonio libres son la condición más primitiva y todos los diferentes tipos de unión o concrescencia son derivados. La soldadura de las piezas del perianto determina muchas veces una mejor protección de los órganos reproductores respecto a las inclemencias atmosféricas o a los animales visitantes y una mejor coordinación espacial y fijación de los órganos florales frente a los animales polinizadores. A veces permite ofrecer a éstos mejores superficies para posarse, mejor acceso al néctar, mayor facilidad para entrar en contacto con los estambres y los estigmas. La concrescencia de las piezas del perianto también permite el desarrollo evolutivo de estructuras especializadas directamente involucradas en la polinización, por ejemplo, la formación de espolones nectaríferos en la base de los pétalos (Aquilegia, Corydalis, Viola, Orchis), o paracorolas (Narcissus).[14]
Hay que tener en cuenta, asimismo, la importancia que presentan en Sistemática la posición y superposición lateral de las piezas del perianto, a la que se le da el nombre de prefloración. Esta disposición se debe observar en el pimpollo debido a que en la flor completamente abierta a menudo se han separado tanto las piezas florales unas de otras que la prefloración no puede determinarse. Hay seis tipos principales de prefloración. En la prefloración valvar las piezas florales pueden llegar a tocarse por los bordes, pero sin que ninguna de ellas se coloque por encima o por debajo de las inmediatas, en la contorta cada una monta sobre la que le sigue y es solapada por la que le precede; en la quincuncial, existen dos piezas totalmente externas, dos totalmente internas y la quinta pieza, es externa por uno de los bordes e interna por el otro. En la prefloración imbricada, hay una pieza externa por ambas orillas, otra, contigua a la anterior, totalmente interna, y las tres restantes son externas por un borde e internas por el otro. La prefloración vexilar es una variante de la precedente, con la pieza totalmente interna contigua a la externa, la cual ocupa una posición superior. Finalmente, en la prefloración carinal la pieza externa de la prefloración vexilar se hace interna y una de las piezas inferiores pasa a ser exterior. La prefloración es importante en la descripción e identificación de plantas ya que, a menudo, caracteriza a las diferentes familias. Así, las malváceas presentan prefloración valvar, las gencianáceas, contorta; y en las leguminosas, la prefloración vexilar y la carinal son características.[21]
Cáliz
El cáliz es el verticilo más externo de la flor. Tiene función protectora y está constituido por los sépalos, generalmente de color verde. Si los sépalos están libres entre sí, el cáliz se denomina dialisépalo, mientras que si están unidos se llama gamosépalo como en el "clavel" (Dianthus caryophyllus, cariofiláceas) o el "seibo" (Erythrina crista-galli, leguminosas).[2]
 Detalle del espolón nectarífero de la flor de Tropaeolum majus.
Detalle del espolón nectarífero de la flor de Tropaeolum majus.Cuando el cáliz es gamosépalo se pueden distinguir tres partes bien definidas: el tubo, que es la porción en la cual los sépalos están unidos; la garganta, que es el sitio en que los sépalos se separan unos de otros; y el limbo, que es la porción libre, formada por los extremos apicales de cada sépalo o lóbulos.
Los sépalos pueden tener consistencia y forma variadas. En la familia de las compuestas, por ejemplo, los sépalos están reducidos a pelos o cerdas que constituyen el denominado papus o vilano.
Según su duración con respecto a las otras piezas florales, el cáliz puede ser efímero o fugaz, cuando los sépalos caen al abrirse la flor, como en la "amapola" (Papaver rhoeas, papaveráceas); deciduo, cuando los sépalos se desprenden después de que ha ocurrido la fecundación; o persistente cuando permanece después de la fecundación y acompaña al fruto, como en el caso del "manzano" (Malus domestica, rosáceas).[22] [23] [14] Los sépalos pueden formar espolones que llevan néctar (se dicen espolones nectaríferos) para atraer a los polinizadores como ocurre en Viola y Tropaeolum,. En otras especies como por ejemplo en la "alegría" (Impatiens balsamina), algunas de las piezas del cáliz adquieren la forma y coloración de los pétalos y, por esa razón, se las adjetiva como "petaloides".[23] [24]
Corola
La corola es el verticilo interno del perianto y el que rodea a los verticilos fértiles de la flor. Esta compuesto por antófilos denominados pétalos, los que son generalmente mayores que los sépalos y son coloreados.
 Flor de Mimulus. Flor bajo luz natural (izquierda) y bajo luz ultravioleta (derecha) mostrando las Guías de néctar que no son visibles para el ojo humano.
Flor de Mimulus. Flor bajo luz natural (izquierda) y bajo luz ultravioleta (derecha) mostrando las Guías de néctar que no son visibles para el ojo humano.Cada pétalo consta de una uña que lo fija al receptáculo y una lámina o limbo que es la parte más ancha y vistosa. La uña puede ser muy corta, como en Rosa o muy larga como en el "clavel" (Dianthus). La lámina puede ser de forma, color y margen muy variado.[2] Si los pétalos son libres entre sí la corola se denomina dialipétala; si, en cambio, se hallan unidos entre sí a través de sus márgenes, la corola se dice gamopétala y, como en el caso del cáliz, presenta tubo, garganta y limbo.[24]
La forma de la corola gamopétala puede ser muy variada: tubulosa (con forma de cilindro, como en las flores centrales de los capítulos de la familia de las compuestas), infundibuliforme (con forma de embudo, como en el caso de la "batata", Ipomoea batatas, convolvuláceas); campanulada (como un tubo inflado, parecido a una campana, como en el "muguet", Convallaria majalis, ruscáceas), hipocrateriforme (con forma de tubo largo y delgado, como en Jasminum), labiada (con el limbo formado por dos segmentos desiguales, como en el caso de Salvia splendens, una lamiácea), ligulada (con el limbo con forma de lengüeta, la cual se observa en las flores periféricas de los capítulos de muchas compuestas) y espolonada (cuando presenta uno o varios espolones nectaríferos, como en el caso de Aquilegia).[25] [23]
La anatomía de los tépalos y pétalos es similar a la de los sépalos. Las paredes de las células epidérmicas frecuentemente son convexas o papilosas, especialmente en la cara adaxial. En muchos pétalos, como los de Brassica napus, las papilas son cónicas, con un engrosamiento cuticular marcado en el ápice, y estrías radiales hacia la base. Se ha sugerido que estos engrosamientos permiten una difusión pareja de la luz emergente, de manera que el brillo de los pétalos es uniforme en cualquier ángulo de iluminación. Algunas células epidérmicas de los pétalos son osmóforos, contienen aceites esenciales que imparten la fragancia característica a las flores. El mesófilo generalmente no presenta parénquima clorofiliano, sino parénquima fundamental.[24] [26]
El color de los pétalos resulta de la presencia de pigmentos. En muchas flores las células presentan cromoplastos con pigmentos carotenoides (rojos, anaranjados, amarillos). Los pigmentos más importantes son los flavonoides, principalmente antocianinas, que se encuentran disueltos en el citoplasma de la célula; los pigmentos básicos son pelargonidina (de color rojo), cianidina (violeta), y delfinidina (azul), flavonoles (de color amarillo a marfil). El color de los pigmentos antociánicos depende de la acidez (del pH) del jugo celular: en la solanácea Brunfelsia pauciflora ("azucena del monte") las flores son violáceas, pero al envejecer se vuelven blancas debido a un cambio en el pH. El color blanco de muchas flores, como por ejemplo el de Magnolia grandiflora, se debe al fenómeno de reflexión total de la luz. Los pétalos pueden presentar espacios de aire en posición subepidérmica o una capa de células con abundantes granos de almidón, y en ambos casos la luz se refleja. Los colores oscuros, se deben a una absorción total de la luz operada por pigmentos complementarios. En los cultivares de color negro de "tulipán" (Tulipa gesneriana), por ejemplo, hay antocianina azul en las células epidérmicas y caroteno amarillo en las subepidérmicas.[27]
Ha sido establecido que las piezas florales exhiben un mayor grado de absorbancia de luz ultravioleta (UV) que las hojas. F.K. Richtmeyer en 1923 y F.E. Lutz en 1924 fueron los primeros en advertir este fenómeno pero no hallaron una explicación al mismo.[28] [29] En 1933, Lotmar postuló que el fenómeno debía tener un significado biológico. Karl Daumer en 1958 observó que, como regla, las bases de los pétalos y los verticilos reproductivos absorben más luz ultravioleta que las regiones periféricas de la corola.[30] En 1972 se hallaron las bases de estos patrones de reflectancia de la luz UV. Utilizando las flores liguladas de una especie de Rudbeckia (compuestas), se demostró claramente que la fuerte absorbancia en la base de las lígulas se debía a la existencia de flavonolglucósidos, los cuales tienen su máximo espectro de absorción en la región del UV (340 a 380 nm de longitud de onda).[31] Esta demostración brindó la base química para la existencia de las guías de néctar de las plantas, las que son invisibles al ojo humano pero a las cuales responden los insectos polinizadores. En un estudio en el que se observaron los patrones de reflectividad de luz UV de 54 especies pertenecientes a 22 familias, se observó que la fuerte absorción de luz UV puede darse en la flor entera o bien en zonas limitadas, tales como las bases de las corolas, las anteras o puntos precisos de los pétalos, en la zona de la flor donde los insectos suelen aterrizar. Estas zonas se denominan "guías de néctar" y sirven como medio de reclamo para los insectos polinizadores. El color particular, visible sólo para los insectos, se denomina "púrpura de abejas".[32] [33] [34]
Androceo
El androceo es uno de los ciclos fértiles de las flores. Las piezas que integran el androceo se denominan estambres, los cuales tienen como función la generación de los gametofitos masculinos o granos de polen.
Los estambres son hojas muy modificadas formadas por un pie que se inserta en el receptáculo de la flor, llamado filamento, y una porción distal llamada antera. El filamento es la parte estéril del estambre, puede ser muy largo, corto o faltar, en ese caso las anteras se denominan sésiles. Generalmente es filiforme, pero puede ser grueso, incluso petaloide, y puede estar provisto de apéndices. La antera es la parte fértil del estambre y suele constar de dos partes distinguibles, contiguas, llamadas tecas, unidas por una zona llamada conectivo, que es también por donde la antera se une al filamento. Generalmente está formada por dos tecas, a veces puede estar constituida por una sola teca como en las malváceas y cannáceas o por tres en el caso de Megatritheca (esterculiáceas). Si se corta la antera perpendicularmente a su eje, se observa que cada antera contiene uno o dos sacos polínicos que se extienden en toda su longitud.[35] [24]
Después de la maduración de los granos de polen se produce la dehiscencia o apertura de la antera para dejar salir el polen. El tejido responsable se llama endotecio. Si la apertura se produce a todo lo largo del tabique que separa los sacos polínicos, la dehiscencia es longitudinal, que es el caso más frecuente. En otros casos el endotecio se localiza en zonas limitadas que luego se levantan como valvas o ventanillas: en la dehiscencia poricida (como en las solanáceas) no hay endotecio, se produce la destrucción del tejido en el ápice de la antera y se forman poros por donde saldrá el polen.[35]
El número de estambres en cada flor es un carácter muy variable. Algunas especies de la familia de las euforbiáceas tienen flores con un solo estambre (se dicen monandras), las oleáceas presentan dos estambres (flores diandras) y en las mirtáceas hay numerosos (son poliandras). El número de estambres puede ser igual o no al número de pétalos. Así, se dice que la flor es isostémona si presenta el mismo número de estambres que de pétalos (como puede observarse en las liliáceas y amarilidáceas); anisostémona si es diferente la cantidad de pétalos que de estambres (por ejemplo, en el género Brassica, hay cuatro pétalos y 6 estambres); diplostémona, cuando el número de estambres duplica a la cantidad de pétalos (el género Kalanchoe, por ejemplo, presenta cuatro pétalos y ocho estambres) y es polistémona cuando el número de estambres es más que el doble de pétalos (como en Poncirus, con cinco pétalos y numerosos estambres).[36]
La disposición helicoidal de numerosos estambres es la condición primitiva dentro de las angiospermas, a la que se denomina poliandria primaria. La reducción del número de estambres (oligomerización) y el paso de la condición verticilada a cíclica fue una tendencia evolutiva dentro de las angiospermas; inicialmente a través de la formación de varios ciclos de estambres, luego dos (diplostemonía) y, finalmente, uno solo (haplostemonía). No obstante, en algunos linajes de angiospermas no es infrecuente que el número de estambres aumente (poliandria secundaria), fenómeno que se observa en especies que ofrecen a sus polinizadores una cantidad de polen especialmente grande.[24] Muchas veces las flores presentan un perianto reducido y los estambres son largos y vistosos. En estos casos la función de atracción de los polinizadores la cumple el androceo. Este tipo de flores suele estar dispuesto en inflorescencias que, por su forma, parecen cepillos o limpiatubos, como por ejemplo en algunas leguminosas (Inga uruguensis y Acacia caven) y en las mirtáceas (Callistemon rigidus).[35]
Los estaminodios son los estambres estériles que de forma normal aparecen en ciertas flores. Su función es variada y puede tener que ver con la producción de néctar o con la función de atracción que suelen cumplir los pétalos.[24]
Gineceo
En las angiospermas el gineceo, también llamado pistilo, consta de uno o más carpelos u hojas carpelares que forman una cavidad, el ovario, dentro de la cual quedan protegidos los óvulos o primordios seminales, tanto de la desecación como del ataque de insectos fitófagos.
El gineceo consta de tres partes: el ovario, parte inferior abultada que forma una cavidad o lóculo en cuyo interior se encuentran los óvulos; el estilo que es una columna más o menos alargada que soporta al tercer componente del pistilo: el estigma. Éste está constituido por un tejido glandular especializado para la recepción de los granos de polen. En ocasiones el estilo puede faltar, y en esos casos el estigma se dice que es sésil.[24]
Si los carpelos están separados o libres entre sí, el gineceo se denomina dialicarpelar o apocárpico (como ocurre en las crasuláceas como Sedum y Kalanchoe); si, por el contrario, los carpelos están soldados entre sí se llama gamocarpelar o sincárpico, que es lo más frecuente.[24]
En la flor dialicapelar cada carpelo constituye un pistilo, mientras en la sincárpica hay un solo pistilo. Por ejemplo, Kalanchoe, con cuatro carpelos libres, presenta cuatro pistilos. En el gineceo gamocarpelar o sincárpico la unión de los carpelos puede afectar sólo a la porción del ovario, por lo que quedan libres tanto los estilos como los estigmas (ejemplo, el género Turnera); puede involucrar los ovarios y los estilos, quedando libres los estigmas (como ocurre en las compuestas y en Hibiscus), por lo que se puede determinar el número de carpelos que conforman el pistilo por observación de la cantidad de estigmas. Finalmente, en muchas ocasiones la unión o soldadura de los carpelos es total. En estos casos el número de carpelos puede determinarse a través del número de los lóbulos estigmáticos (por ejemplo, en las bignoniáceas).
 Esquema de las posibles posiciones del ovario I ovario súpero, II ovario semiínfero, III ovario ínfero. a: androceo, g: gineceo, p: pétalos, s: sépalos, r: receptáculo.
Esquema de las posibles posiciones del ovario I ovario súpero, II ovario semiínfero, III ovario ínfero. a: androceo, g: gineceo, p: pétalos, s: sépalos, r: receptáculo. Microfotografía de un óvulo de una monocotiledónea.
Microfotografía de un óvulo de una monocotiledónea.El estilo es de longitud variable, desde menos de 0,5 mm (estigma subsésil) hasta más de 30 cm en ciertas variedades de maíz, que es lo que se conoce como barba de choclo. Generalmente nace en el ápice del ovario, pero puede ser lateral o nacer aparentemente en la base (estilo ginobásico).[37] Desde el punto de vista anatómico, el estilo puede ser sólido o hueco. En las plantas con estilo hueco el tejido de transmisión (por donde crecen los tubos polínicos para efectuar la fecundación) está constituido por una capa de células epidérmicas bastante diferenciadas que rodean un canal hueco (el denominado canal estilar). Los tubos polínicos crecen desde el estigma hacia el ovario a lo largo de la superficie de ese canal, normalmente a través de una fina capa de mucílago. En las plantas que poseen estilos sólidos, en cambio, las células epidérmicas se hallan íntimamente fusionadas y no dejan ningún espacio entre sí. Los tubos polínicos, en este caso, crecen entre las células del tejido de transmisión (como en el caso de Petunia,[38] ) o a través de las paredes celulares (como en Gossypium,[39] ). El tejido de transmisión en los estilos sólidos incluye una sustancia intercelular que contiene pectina, comparable al mucílago que se encuentra en el canal estilar de los estilos huecos.[40] Desde el punto de vista de la distribución de ambos tipos de estilos entre las diferentes familias de angiospermas, los estilos sólidos se consideran típicos de las eudicotiledóneas y son raros en las monocotiledóneas.[41] [42]
El estigma tiene forma variable, plumoso en el caso de las gramíneas, en cabezuela en Citrus, lobulado en Cucurbita, petaloide en Canna y hasta con forma de sombrilla invertida en el caso de Sarracenia. Tiene particularidades estructurales que permiten la germinación del polen y el desarrollo del tubo polínico que llegará hasta los óvulos. Se ha comprobado que el estigma esta cubierto por proteínas hidrofílicas en la pared externa; son probablemente las que actúan en el reconocimiento del polen adecuado y en las reacciones de autoincompatibilidad, en cuyo caso a veces de deposita calosa para detener la germinación del polen incompatible.[37]
Los estigmas se dividen en dos grandes grupos: estigmas húmedos y secos.[43] Los estigmas húmedos liberan un exudado durante el período receptivo y se presentan en familias como las orquidáceas, escrofulariáceas y solanáceas. Los estigmas húmedos pueden presentar papilas (estigmas papilosos, como en Annona, Mandevilla, Bignonia y Punica) o no presentarlas (estigmas no papilosos, en Citrus, Impatiens, Opuntia y Tamarix ). Los estigmas secos no liberan secreciones líquidas, sino que producen proteínas o ceras. Pueden ser plumosos (gramíneas) o no plumosos y, en este caso, papilosos (Cordyline, Yucca Pelargonium o no papilosos (Asclepias, Capparis, Cyperus).[37]
El ovario es la parte inferior del gineceo que contiene a los óvulos a fecundar. Está formado de una o más hojas modificadas que reciben el nombre de carpelos. Dentro del ovario hay una o más cavidades o "loculos" que contienen a los óvulos en espera de ser fecundados. Los óvulos es insertan dentro del ovario en una zona denominada placenta.
Una de las terminologías de descripción del ovario se refiere al punto de inserción sobre el receptáculo (donde las otras partes florales (periantio y androceo) se unen y se fijan a la superficie del ovario. Si el ovario se sitúa arriba del punto de inserción, será súpero; si es por debajo, ínfero.
El óvulo o rudimento seminal, nace sobre la placenta, situada en la cara interna del carpelo. Son de tamaño reducido, de pocos milímetros, y generalmente de forma ovoide, de allí su nombre. Cada óvulo consta de un cuerpo de tejido compacto, la nucela y un pie, el funículo, que lo une a la placenta. La región basal, donde se unen el funículo y la nucela, es la cálaza o chalaza. La nucela está rodeada por el o los tegumentos, los cuales son envolturas que parten de la cálaza y dejan un orificio llamado micrópilo. Cada óvulo está inervado por un haz vascular que atraviesa el funículo y llega hasta la cálaza. Este hacecillo puede ramificarse e inervar los tegumentos. El número de tegumentos es constante en cada familia o grupo de familias. Las gimnospermas, por ejemplo, tienen óvulos unitégmicos, a excepción de las podocarpáceas que presentan un segundo tegumento llamado epimacio. La mayoría de las dicotiledóneas y monocotiledóneas tienen óvulos bitégmicos, y el tegumento externo se llama primina y el interno secundina. Los óvulos unitégmicos se presentan en muchas dicotiledóneas gamopétalas y en las familias de monocotiledóneas orquídeas, gramíneas y amarilidáceas. El micrópilo puede estar delimitado por uno de los tegumentos (ejemplo, Lilium) o por ambos tegumentos (como el caso de Gossypium). La nucela tiene espesor variable. Si es pequeña, el óvulo se dice tenuinucelado; si tiene varias capas de células por fuera del saco embrionario, se denomina crasinucelado.[44] [45]
La oósfera procede de una espora, denominada megáspora o macrospora, a través de un proceso denominado megagametogénesis que consiste básicamente en divisiones mitóticas. Su núcleo en general es haploide, o sea, posee la mitad de los cromosomas que la planta que le dio origen. La oósfera se fusiona con uno de los núcleos generativos del grano de polen durante la doble fecundación para originar al embrión. El otro núcleo generativo se fusionará con los núcleos polares de la célula media y originará al endosperma.[24]
La disposición que adoptan los óvulos dentro de la cavidad del ovario se denomina placentación. El número de placentas es, en general, igual al número de carpelos que forman el ovario. En ciertos casos, sin embargo, puede atrofiarse alguna de las placentas y un ovario pluricarpelar llega a contener un sólo óvulo, como por ejemplo en las gramíneas y en las compuestas. Existen distintos tipos de placentación. La placentación marginal, propia del gineceo unicarpelar (leguminosas) o dialicarpelar (magnoliáceas, ranunculáceas), donde cada carpelo tiene una sola placenta que corresponde a la zona de soldadura de la hoja carpelar. La placentación parietal, presente en las orquídeas, violáceas, pasifloráceas y cucurbitáceas, ocurre en el gineceo formado por dos o más carpelos soldados por sus bordes formando una sola cavidad en el ovario, de manera que cada placenta corresponde a los bordes de dos hojas carpelares contiguas. La placentación axilar, propia de solanáceas, rutáceas, liliáceas e iridáceas, entre muchos otros ejemplos, ocurre en el gineceo formado por dos o más carpelos soldados en que cada uno lleva la placenta en el ángulo central, de manera que las suturas placentarias forman una columna adentro del ovario. Los óvulos de cada lóculo quedan así aislados de los vecinos por medio de los tabiques carpelares. La placentación central, propia de las primuláceas y cariofiláceas, ocurre en gineceos uniloculares formados por dos o más carpelos unidos y donde los óvulos están fijos sobre una columna central y sin tabiques con la pared del ovario. La placentación basal, finalmente, ocurre en especies con gineceo pluricarpelar y unilocular. El óvulo se dispone en el centro basal de la cavidad del ovario y es típica de las familias de las poligonáceas, quenopodiáceas y compuestas.[24] [45]
Sexualidad
La sexualidad floral está relacionada con la presencia o ausencia de los verticilos reproductivos: androceo y gineceo. Las flores que presentan ambos verticilos (o sea, que producirán tanto gametos femeninos como masculinos) se dicen perfectas, bisexuales, monoclinas o, más frecuentemente, hermafroditas, como es el caso de las flores de la "papa" o "patata" (Solanum tuberosum, solanáceas). En muchas otras especies, por otro lado, las flores tienen un solo verticilo reproductivo y se dicen diclinas, imperfectas o unisexuales, como ocurre en la "morera" (Morus nigra, moráceas) y en la "calabaza" (Cucurbita maxima, cucurbitáceas). En el caso que tal verticilo sea el androceo, las flores se denominan masculinas o estaminadas; mientras que si el verticilo presente es el gineceo, las flores se denominan pistiladas o femeninas. Existen algunos casos en que la flor no presenta ninguno de los verticilos reproductivos y solo muestra sépalos y pétalos. La función de estas flores es el de especializarse, dentro de un grupo de flores que sí son perfectas, en la atracción hacia la inflorescencia de los insectos polinizadores. Tales flores, denominadas neutras o asexuales, se disponen usualmente en la periferia de la inflorescencia y se pueden observar, por ejemplo, en muchas especies de la familia de las compuestas, como la "margarita" (Bellis perennis) o el "girasol" (Helianthus annuus).[15]
Entre los botánicos prevalece actualmente la opinión de que las flores hermafroditas son menos evolucionadas que las flores unisexuales, al menos por las razones que se exponen a continuación. En primer lugar, en casi todos los grupos de angiospermas con flores unisexuales, se encuentran entre las flores femeninas y masculinas rudimentos de estambres o carpelos (por ejemplo, en Castanea y Urtica). En segundo lugar, los grupos de angiospermas más primitivos en cuanto a otros caracteres tienen principalmente flores hermafroditas (por ejemplo, las magnoliáceas) ya que sólo este tipo de flores hacen posible, en caso de polinización zoógama, la captación y cesión simultánea del polen. Finalmente, el paso de la condición hermafrodita a la unisexual ha sido determinada en numerosas ocasiones por vía selectiva a consecuencia de una transformación secundaria de flores entomógamas en anemógamas, como por ejemplo, en las aceráceas (Acer) y en algunas oleáceas (Fraxinus).[14]
Fórmula y diagrama floral
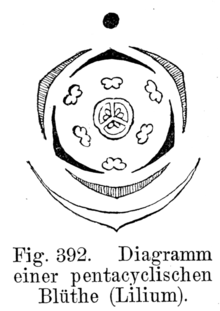
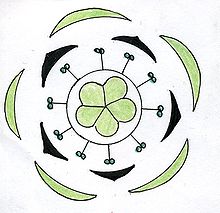
El diagrama floral es una representación gráfica de la disposición de las piezas florales y de la ordenación de los distintos verticilos, en un corte transversal de la flor. Cada verticilo se representa, por convención, con una circunferencia concéntrica alrededor del gineceo, indicado por un corte a la altura del ovario. Los sépalos se dibujan como lúnulas blancas, los pétalos como lúnulas negras o, a veces, coloreadas. Los estambres se simbolizan con cortes transversales de antera, y el gineceo queda representado en el centro del diagrama por un corte transversal del ovario. Generalmente las piezas de un verticilo alternan con las piezas del verticilo anterior. Los estambres pueden estar opuestos o alternos con respecto a los pétalos. La soldadura entre las piezas de cada verticilo o de verticilos opuestos, se indica con líneas de puntos. Mediante estos diagramas queda bien manifiesta la estructura floral, se deduce de él su simetría, el número de miembros de cada verticilo, y tratándose de los sépalos y pétalos, su prefloración, su soldadura o independencia; del androceo la posición relativa de los estambres con respecto al perianto, la concrescencia o separación de los mismos, su unión o autonomía con respecto a la corola y la posición introrsa o extrorsa de las anteras. Del gineceo, se puede observar el número de carpelos y de cavidades en el ovario y la placentación de los óvulos.[21] En el esquema de la derecha se observa el diagrama floral de Lilium, típico de la familia de las liliáceas. El diagrama muestra que las flores son hermafroditas (tienen estambres y pistilo), actinomorfas (presentan varios planos de simetría). El perigonio está formado por 6 tépalos dispuestos en dos verticilos trímeros (o sea, cada uno de ellos está formado por tres piezas), se hallan separados entre sí y libres de las demás piezas florales (se dice, entonces, dialitépalo). El androceo presenta 6 estambres dispuestos en 2 verticilos también trímeros, los filamentos se hallan separados entre sí y libres de las demás piezas florales. El androceo es diplostémono (es decir que el verticilo externo de estambres es opuesto a los tépalos externos y el ciclo interno es opuesto a los tépalos internos). El gineceo es de ovario súpero y está formado por 3 carpelos connados, es trilocular. Los óvulos presentan placentación axilar. En el esquema de abajo se puede observar el diagrama floral de una oxalidácea: Oxalis. Nuevamente, se aprecia que la flor es hermafrodita y actinomorfa, pentacíclica (o sea, presenta 5 verticilos florales) y pentámera (cada ciclo está compuesto por cinco piezas).
La fórmula floral es un modo de representar simbólicamente la estructura de una flor mediante el uso de letras, números y otros signos. Típicamente, se utiliza la fórmula floral para representar las características morfológicas de las flores de una determinada familia de plantas, más que de una especie en particular. Las siguientes son los símbolos más utilizados:[46]
K = cáliz; por ejemplo, «K5» indica que la flor tiene 5 sépalos.
C = corola; por ejemplo, «C3» significa que la flor presenta 3 pétalos.
Z = se agrega si la corola es cigomorfa; ejemplo, «CZ»6 = indica una corola cigomorfa con 6 pétalos
A = androceo; por ejemplo, «A∞» significa que presenta muchos estambres
G = gineceo; así, «G(3)1∞» indica un gineceo de ovario súpero (por eso la "G" está subrayada), compuesto por 3 carpelos unidos entre sí (por eso el número tres se pone entre paréntesis), con un solo lóculo y numerosos óvulos por lóculo. La cantidad de lóculos del ovario se indica como subíndice del número de carpelos; el número de óvulos por lóculo se indica como exponente.Una fórmula floral, entonces, adquiere una forma como la siguiente, típica de la familia de las liliáceas:
-
- K3 C3 A6 G(3)3∞
La cual indica que las flores de esta familia presentan 3 sépalos, 3 pétalos, un androceo con 6 estambres y un gineceo de ovario súpero formado por 3 carpelos unidos entre sí, con tres lóculos y numerosos óvulos por lóculo. La fórmula floral que describe simbólicamente la estructura floral de Oxalis, previamente descripta, es:
-
- K5 C5 A10 G(5)5∞
Analogías y homologías
 Las brácteas de color rojo que rodean a las inflorescencias de Euphorbia pulcherrima adoptaron la función de atracción que usualmente presentan los pétalos.
Las brácteas de color rojo que rodean a las inflorescencias de Euphorbia pulcherrima adoptaron la función de atracción que usualmente presentan los pétalos.Durante la diversificación de las angiospermas, diferentes linajes evolucionaron de modo tal de utilizar el mismo patrón morfológico básico de modos diferentes. Por ejemplo, en la "estrella federal" (Euphorbia pulcherrima) las brácteas que rodean a las flores lograron un color rojo brillante y comenzaron a funcionar como órganos de atracción para los polinizadores. Los "pétalos" de la "estrella federal", entonces, no son en realidad pétalos sino órganos homólogos a las brácteas de las restantes fanerógamas, lo que significa que ambos evolucionaron a partir de la misma estructura en un ancestro común. Desde el punto de vista funcional, las brácteas de la estrella federal parecen y funcionan como los pétalos de cualquier otra planta, lo que indica que ambas estructuras son análogas o similares debido a una evolución convergente. Los pétalos reales de la "estrella federal" se perdieron durante su evolución ya que, de hecho, sus flores, diminutas y casi imperceptibles, no presentan corola.
Las costáceas ofrecen otro ejemplo. Dos de los estambres de las flores de esta familia son bastante inusuales, ya que durante la evolución, los mismos han perdido su función original (son estériles), se han fusionado y formaron una estructura muy llamativa denominada labelo. Este labelo es homólogo a los estambres y análogo a los pétalos de los restantes linajes de angiospermas.[47]
Inflorescencias
 No es una flor, sino una inflorescencia, formada en este caso por un círculo de flores blancas rodeando a un contingente de flores amarillas.
No es una flor, sino una inflorescencia, formada en este caso por un círculo de flores blancas rodeando a un contingente de flores amarillas.Las flores pueden aparecer solitarias o en parejas, pero lo más común es que crezcan juntas en grupos organizados que se llaman inflorescencias. En ocasiones las flores aparecen tan apretadas y ordenadas que toda la inflorescencia puede confundirse con una flor, en cuyo caso podemos referirnos a ella como un pseudanto (falsa flor). El caso más notable lo ofrecen las plantas de la familia de la margarita o el girasol (compuestas), cuya inflorescencia, denominada capítulo está integrada por decenas o cientos de flores.[2] [48] [14]
Los órganos constitutivos de las inflorescencias plurifloras son las flores provistas o no del pedicelo, el eje o receptáculo común, el pedúnculo y las brácteas. El pedicelo es la parte del tallo que sostiene la flor; a veces es muy corto, y otras veces es nulo, en cuyo caso la flor se dice sentada o sésil. El eje o raquis es la parte alargada del tallo que lleva las ramas floríferas; si es corto y está ensanchado en forma de plato se llama receptáculo común. El pedúnculo, es la parte del tallo que soporta el raquis o el receptáculo común. El eje que sale de la base arrosetada de la planta o de un órgano subterráneo se llama escapo (como por ejemplo en Amaryllis, Agapanthus y Taraxacum).[48] [2]
Las brácteas o hipsófilos son las hojas modificadas, generalmente de menor tamaño que las hojas normales, coloreadas o verdosas, que nacen sobre el raquis o acompañan a las flores. Algunas veces faltan, como en el caso de las crucíferas, otras veces reciben nombres especiales, tales como glumas y glumelas en las gramíneas y ciperáceas, o espata en las aráceas y palmeras. En otros casos las brácteas forman órganos protectores de las flores (involucros), como la cúpula de los robles (Quercus) y el erizo del castaño. El prófilo o bracteola es la primera bráctea de una rama axilar, está dispuesta del lado opuesto a la hoja normal. En las monocotiledóneas es bicarenada y por el dorso, cóncavo, se adosa el eje que lleva la rama. En las espiguillas de las poáceas el prófilo recibe el nombre de pálea o glumela superior.[48] [2]
Polinización
 Las flores de muchas especies de angiospermas están evolutivamente diseñadas para atraer y recompensar a una o pocas especies polinizadoras (insectos, aves, mamíferos). Su gran diversidad en cuanto a forma, color, fragancia y presencia de néctar es, en muchos casos, el resultado de la coevolución con cada especie de polinizador.
Las flores de muchas especies de angiospermas están evolutivamente diseñadas para atraer y recompensar a una o pocas especies polinizadoras (insectos, aves, mamíferos). Su gran diversidad en cuanto a forma, color, fragancia y presencia de néctar es, en muchos casos, el resultado de la coevolución con cada especie de polinizador.Atracción de los polinizadores
La zoofilia que caracteriza a muchas angiospermas presupone que los animales polinizadores visiten las flores de manera regular y se detengan en ellas el tiempo suficiente; que las anteras y el estigma sean rozados o tocados con cierta frecuencia y que el polen quede adherido a los visitantes de modo tan perfecto que pueda llegar con la debida seguridad a los estigmas de otras flores. El resultado de la zoofilia depende esencialmente de que los animales puedan reconocer las flores desde una cierta distancia y de que se vean compelidos a visitar durante un cierto tiempo las flores de la misma especie. Las flores zoófilas, entonces, deben poseer productos atractivos (cebos, como el polen y el néctar), medios de reclamo (tales como olores y colores) y, además, polen viscoso o adherente.[14] En el curso de la evolución de las angiospermas se ha producido una diferenciación muy intensa de los medios de atracción y reclamo, así como de la forma de la flor; gracias a ello un número cada vez más grande de animales ha podido colaborar en la polinización. De la visita casual de las flores por animales variados se ha pasado de modo progresivo, en el transcurso de la evolución, al establecimiento de estrechas relaciones entre los «animales antófilos» y las «flores zoófilas», con evidentes ventajas para ambos grupos. Para las plantas implicó una precisión creciente en la atracción de solo determinados visitantes y una transferencia del polen a los estigmas de otras plantas cada vez más segura, lo que resultó en un ahorro progresivo en la producción de polen. De hecho, la relación entre el número de granos de polen y el número de óvulos producidos por una flor es del orden de un millón para las plantas anemófilas, mientras que en las orquídeas es de uno. Para los animales polinizadores especializados, la competencia con otros animales antófilos resultó disminuida y la polinización orientada o especializada en una sola especie pasó a ser, en última instancia, ventajosa para ellos.[14] El desarrollo evolutivo de las angiospermas zoófilas y de los grupos de animales que se han ido adaptando a ellas sólo puede comprenderse como una coevolución condicionada por relaciones recíprocas. La adaptación entre sí de muchas plantas y sus polinizadores ha llegado a veces tan lejos que no pueden existir el uno sin el otro.[14]
Productos atractivos
Son los cebos que utilizan las flores como recompensa al agente polinizador. Básicamente se trata de alimentos, como polen y néctar; o sustancias químicas que los insectos utilizarán en sus rituales de apareamiento.[49] El polen es una recompensa generalmente en aquellas flores polinizadas por coleópteros e himenópteros, atraídas por el fuerte aroma que despide. El polen contiene proteínas, almidón, aceites y otros nutrientes. El néctar es un líquido azucarado producido por estructuras llamadas nectarios. Se ubican estratégicamente en la flor para que los insectos al tomarlo toquen los estambres y se lleven el polen de una flor a otra.[50]
Medios de reclamo
Los medios de reclamo indican la presencia de la recompensa y pueden ser visuales u olfativas. Generalmente esta función la ejercen el cáliz y la corola. Las claves visuales están determinadas básicamente por el color de los pétalos, los de color rojo en general atraen aves, los amarillos a las abejas y los blancos a las mariposas nocturnas. Las claves olfativas son utilizadas para atraer a los polinizadores desde una distancia mayor. Se trata de las fragancias que despiden las flores gracias a compuestos volátiles excretados por estructuras llamadas osmóforos. Estas fragancias pueden ser agradables o desagradables para los seres humanos, como por ejemplo las que atraen a las moscas.[51] La forma de la flor también debe estar adaptada a cada tipo de polinizador, evitando al mismo tiempo a los ladrones de néctar. Aquellas flores polinizadas por escarabajos, por ejemplo, deben tener una constitución que soporte el peso del insecto y proteja los óvulos de la voracidad de los mismos. Para los colibríes e insectos que polinizan durante el vuelo, las flores deben tener corolas tubulosas, adecuadas a los picos de los pájaros y a las espiritrompas de las mariposas. Para las abejas, las flores son amplias y planas, brindando "pistas de aterrizaje" para el insecto.[52] [53]

 Colibrí polinizando una flor.
Colibrí polinizando una flor.El conjunto de los medios de reclamo que presenta una planta para atraer a un determinado agente polinizador se denomina síndrome floral. Los diferentes síndromes florales en relación a la polinización por animales son:[53] [54] [55]
- Melitofilia: es un sistema por medio del cual ciertas especies de plantas atraen a insectos himenópteros, en particular abejas y avispas, para que estos realicen la polinización. Los himenópteros tienen un aparato bucal suctor y buscan para alimentarse néctar que esté accesible, ya que sus órganos bucales son cortos. Las flores de las especies melitófilas atraen a las abejas por medio de una combinación de formas, fragancias y colores. Así, las corolas son amariposadas (por ejemplo, las Leguminosas Papilionóideas como los tréboles y la alfalfa), labiadas (o sea, con los pétalos unidos en dos grupos de modo que forman dos labios, tal como en las labiadas y algunas compuestas) o con forma de fauce. Las corolas presentan superficies para que el insecto se pose y guías de néctar que señalan la ubicación de los nectarios. Asimismo, este tipo de flores produce sustancias aromáticas en osmóforos que se encuentran en la corola (como por ejemplo en Citrus), en la corona (Narcissus), o en otros órganos florales. A veces esta coevolución ha llegado al punto en que la flor, para atraer a los machos de una especie de abeja, imita a la hembra de la misma especie, como ha sido descrito en los estudios relativos a la polinización en orquídeas.[56] [57]
- Psicofilia: las flores que han coevolucionado con mariposas presentan generalmente el néctar en el fondo del tubo de la corola, donde solo el largo aparato bucal suctor de estos insectos puede llegar. Las flores que atraen a mariposas nocturnas en general son pálidas y de olores intensos.
- Miofilia: las flores emiten aromas que semejan el alimento natural o los substratos que visitan las moscas, los cuales comúnmente son materiales orgánicos en descomposición, excrementos o carroña, en búsqueda de alimento o para depositar sus huevecillos.[58] Diferentes partes florales producen olores putrescentes o dulces, comúnmente forman trampas que retienen al polinizador, además de poseer varios apéndices que los atraen y colores ya sea brillantes o que semejan carne o algún otro tinte de materia putrefacta.[59] [60] [61] [62]
- Ornitofilia: las flores de este tipo de síndrome no tienen aroma, ya que los pájaros no tienen sentido del olfato, si presentan colores intensos, rojos y amarillos y abundante néctar.
- Quiropterofilia: es la polinización por murciélagos pequeños. Las flores tienen colores pálidos, fuertes aromas y abundante néctar.
Cada especie tiene un determinado síndrome floral, no obstante, diferentes especies dentro de un mismo género pueden presentar variados síndromes. Así, por ejemplo, el género de solanáceas Schizanthus presenta básicamente especies entomófilas, es decir que requieren que el polen sea transportado de planta a planta por medio de insectos. La mayoría de las especies del género son polinizadas por himenópteros (abejas, abejorros y avispas de los géneros Alloscirtetica, Bombus, y Megachile, entre otros). No obstante, las especies que presentan flores de color blanco (tales como S. candidus, S. integrifolius y S. lacteus) son polinizadas por mariposas nocturnas. Finalmente, Schizanthus grahamii es polinizada por colibríes o picaflores (como, por ejemplo, Oreotrochilus leucopleurus).[63] La gran diversificación en los síndromes florales en Schizanthus ha sido el producto de la adaptación de estas especies a los diferentes grupos de polinizadores existentes en los ecosistemas mediterráneo, alpino de altura y desértico de Chile y áreas adyacentes de Argentina donde el género es originario.[63]
Estas estrategias favorecen la fecundación cruzada y el consiguiente intercambio de genes. Otra manera de lograr ese mismo objetivo es la presencia de barreras a la autofecundación: ya sea genéticas por autoincompatibilidad, o físicas como la separación de sexos (plantas dioicas como el "mamón" (Carica), o monoicas como el "maíz" (Zea mays)), o de los diferentes tiempos de liberación de polen y receptividad del estigma en la misma flor.
Formación de los gametofitos y fecundación
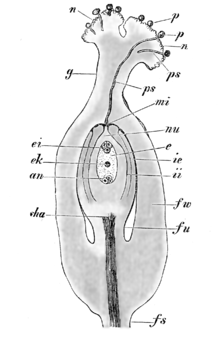 Representación esquemática de los eventos que tienen lugar durante la fertilización de una angiosperma. Leyendas: fu: funículo, cha: chalaza, nu: nucela, mi: micrópila, ii: tegumento interno, ie: tegumento externo, e: saco embrionario, ek: núcleos polares, ei: óosfera y sinérgidas, an: antípodas, g: estilo, n: estigma, p: granos de polen, ps: tubos polínicos. Véase la explicación del proceso en el texto.
Representación esquemática de los eventos que tienen lugar durante la fertilización de una angiosperma. Leyendas: fu: funículo, cha: chalaza, nu: nucela, mi: micrópila, ii: tegumento interno, ie: tegumento externo, e: saco embrionario, ek: núcleos polares, ei: óosfera y sinérgidas, an: antípodas, g: estilo, n: estigma, p: granos de polen, ps: tubos polínicos. Véase la explicación del proceso en el texto.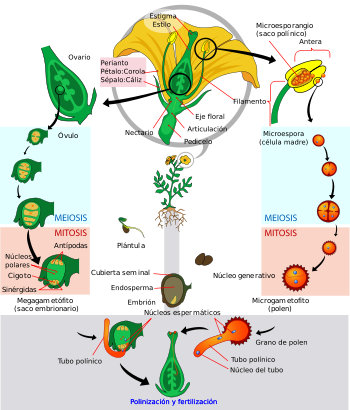 Diagrama del ciclo de vida de una angiosperma.
Diagrama del ciclo de vida de una angiosperma.El gametofito masculino de las angiospermas es el grano de polen que se origina a partir de las microsporas, como se describe a continuación. En el interior de los sacos polínicos de las anteras se encuentran localizadas las células madres de las microsporas, las cuales son las células diploides (tienen el mismo número de cromosomas que cualquier otra célula de la planta) que sufren el proceso de división celular llamado meiosis, luego del cual forman una cuatro microsporas que presentan la mitad del número de cromosomas que la planta original (se dice, entonces, que son haploides). Cada microspora haploide atraviesa dos procesos concomitantes para originar un grano de polen. Por un lado, su pared celular se desarrolla, formando la exina, compuesta de un polisacárido complejo, la esporopolenina y la intina constituida por celulosa. Dicha pared del grano de polen presenta una o varias aperturas y un diseño de la exina que es característico de cada especie vegetal. Por otro lado, el núcleo haploide de cada microspora se divide por mitosis originando dos núcleos, también haploides, uno es el denominado núcleo vegetativo y el otro es el núcleo generativo. Este último vuelve a dividirse, antes o después de la polinización, dando dos núcleos gaméticos o espermáticos (ver figura a la izquierda).[64] [2] [23]
El gametofito femenino de las angiospermas se desarrolla dentro de la nucela del óvulo. Una célula diploide de la nucela sufre meiosis y origina cuatro células haploides denominadas megásporas. Tres de ellas degeneran y la restante sufre tres mitosis sucesivas hasta formar el gametofito femenino, el cual se denomina saco embrionario, con 7 células y 8 núcleos haploides. En el saco embrionario típico (denominado "tipo Polygonum", por ser éste el primer género en el que se describió) se encuentran, desde el extremo dónde se halla la micrópila del óvulo, la oósfera u ovocélula, cuyo núcleo es el gameto femenino; dos células acompañantes o sinérgidas que flanquean a la oósfera y que presentan estructuras especiales en sus paredes responsables de la atracción del tubo polínico; la célula media que se dispone en el centro de saco embrionario y lleva dos núcleos haploides denominados núcleos polares y, finalmente, las tres antípodas que se ubican en el extremo chalazal del saco embrionario, opuesto al sitio donde se halla la oósfera (ver figura a la izquierda).[65] [2] [23]
La fertilización en las angiospermas es un proceso complejo que involucra varias etapas sucesivas: la germinación de los granos de polen en el estigma, el crecimiento del tubo polínico en el estilo orientado hacia el saco embrionario del óvulo y la fusión de los núcleos generativos del gametofito masculino con el núcleo de la oósfera y con los dos núcleos polares. En el esquema de la derecha se muestra como los granos de polen (p) aterrizan en el estigma de la flor (n) y germinan, emitiendo un tubo polínico (ps) gracias al crecimiento de la célula vegetativa del grano de polen. El tubo polínico se elonga dentro del estilo (g) y atraviesa la micrópila (mi). Ya dentro del óvulo, se dirigen hacia el saco embrionario (e) para descargar los núcleos generativos, uno de los cuales se fusiona con el núcleo de la oósfera (la célula central de las 3 señaladas como (ei)) y, el otro, con los dos núcleos polares (ek), produciéndose de ese modo la doble fecundación característica de las angiospermas. La unión del núcleo de la oósfera con uno de los núcleos generativos, ambos haploides, determina la formación de un núcleo diploide denominado cigoto, el que por sucesivas divisiones mitóticas originará el embrión. La fusión del otro núcleo generativo del grano de polen con los dos núcleos polares de la célula media origina un núcleo triploide (tiene dos juegos cromosómicos de la madre y uno del padre) que es el que por sucesivas mitosis originará el endosperma, tejido que brindará nutrientes al embrión durante sus primeras etapas de desarrollo. Ambos, el embrión y el endosperma, constituyen la semilla, la estructura de dispersión típica de las espermatófitas.[66] [2] [23] [67] [24]
Origen y evolución
En 1998 se descubrió en los estratos provenientes del Jurásico Superior (de más de 125 millones de años de antigüedad), en China, un fósil de un eje con frutos, que se ha denominado Archaefructus. La infrutescencia constaba de frutos de tipo folículo, provenientes de carpelos conduplicados (doblados sobre sí mismos a lo largo de su nervadura media), dispuestos helicoidalmente alrededor del eje.[68] Semejante descubrimiento, que parecía datar la edad de las angiospermas más antiguas, hizo mundialmente famosa a la Formación Yixian, donde fue descubierto este fósil. Algunos años más tarde, el fósil de otra angiosperma, Sinocarpus, fue también descubierto en esta formación.[69] No obstante, siempre hubo controversias al respecto de si ambos fósiles estaban anidados en el linaje de las angiospermas primitivas, ya que de Archaefructus sólo se ha encontrado el fruto por lo que no se conoce la estructura de su flor,[70] [71] y la validez del género Sinocarpus ha sido puesta en duda.[72] Una interpretación alternativa del fósil Archaefructus sugiere que el tallo alargado es una inflorescencia en vez de una flor, con flores estaminadas (masculinas) y flores pistiladas (femeninas) por encima.[73] El descubrimiento de Archaefructus eoflora[74] apoya esta interpretación, debido a que flores bisexuales están presentes entre las flores estaminadas y las pistiladas. Si esta interpretación es correcta, Archaefructus podría no ser un miembro extinguido de las angiospermas basales y más bien podría estar relacionada a Nymphaeales o ser un grupo basal de eudicotiledóneas.
No obstante, en 2007 se informó del hallazgo de una flor perfecta en la formación Yixian. Esta flor tiene la organización típica de las angiospermas, incluyendo tépalos, 10 estambres y gineceo. Los tépalos son espatulados. Las anteras son globosas y con granos de polen triangular-redondeados. Los carpelos están rodeados por un receptáculo excepto por los estigmas plumosos que sobresalen a través de un poro. Esta especie ha sido bautizada como Euanthus (del griego, "flor verdadera") por sus descubridores, e indica que en el Cretácico inferior ya existían flores como las de las angiospermas actuales.[75]
La mayor parte de los análisis morfológicos y moleculares disponen a Amborella, las nymphaeales y Austrobaileyaceae en un clado basal denominado "ANA". Este clado parece haber divergido en el Cretácico inferior (alrededor de 130 Ma), una fecha aproximadamente igual a la de los fósiles de la formación Yixian.[76] [77] Las magnólidas difirieron poco después y una rápida radiación produjo las eudicotas y monocotiledóneas hace 125 Ma.[77] Hacia el final del Cretácico ya habían surgido más del 50% de los órdenes actuales de angiospermas, clado que constituye el 70% de las especies del planeta.[78] Las características de los grupos basales "ANA" sugieren que las angiospermas se originaron en áreas oscuras, cenagosas y frecuentemente disturbadas.[79] Aparentemente, permanecieron recluidas en ese tipo de hábitats durante todo el Cretácico.[78] Esto debe haber restringido su importancia inicial, pero sus innovaciones adaptativas, la flor por ejemplo, les otorgaron la notoria flexibilidad que explica la rapidez de su diversificación posterior hacia otros hábitats.[79]
El origen de las flores y de la polinización por insectos
La forma primaria de polinización de las espermatófitas primitivas es indudablemente la anemofilia, o sea, el transporte del polen por el viento. Esta es también la forma principal de diseminación de las esporas en las pteridófitas. La falta de dirección en el transporte de los gametos masculinos hacia los estigmas se supera gracias a la producción de grandes cantidades de polen[b]La plantilla {{Note label}} está obsoleta, véase el nuevo sistema de referencias. y a la gran capacidad de flotación de los mismos por su tamaño diminuto y ligereza. Todas estas características, no obstante, quedan aseguradas solamente si las plantas productoras de polen crecen en lugares abiertos y expuestas al viento y, además, constituyen poblaciones densas con escasa distancia entre individuos.[14] Estos argumentos a favor de la anemofilia de las angiospermas primitivas se han visto confirmados por los análisis de plantas fósiles.[80] [81] [82] La competencia por el alimento y la selección natural en los insectos llevó a ciertos grupos hematófagos a convertirse en fitófagos.[83] Este proceso comenzó antes del Triásico, donde tanto insectos como plantas eran muy abundantes en toda la Tierra. De hecho, los hallazgos de insectos fitófagos conservados en ámbar, combinados con los análisis de fósiles, evidenciaron que la fitofagia en los insectos comenzó mucho antes de la aparición de las primeras flores.[84] Ciertos grupos de insectos se adaptaron a una nueva dieta basada en las partes vegetativas de las plantas y, posteriormente, en las partes reproductivas, en particular el polen y las semillas, puesto que este proceso coincidió en parte con la transición de las pteridofitas con esporas a los helechos con semilla y las primeras gimnospermas y angiospermas.[83] [84] [82]
El cambio de dieta de ciertos grupos de insectos tuvo un impacto muy fuerte sobre las plantas, ya que la presión de selección que ejercía la herbivoría sobre ellas era muy fuerte. Grandes cantidades de insectos devoraban las hojas de las plantas reduciendo la tasa de captación de energía lumínica para la fotosíntesis y el intercambio gaseoso, otros comían partes de los tallos, destruyendo algunas veces las yemas, otras los haces conductores, y las heridas que dejaban aumentaban la probabilidad de infecciones micóticas y virales, disminuyendo significativamente su eficacia biológica. Como respuesta a esta presión, las plantas desarrollaron estrategias de defensa basadas en los denominados metabolitos secundarios.[85] Tales compuestos son moléculas orgánicas no esenciales que actúan a diferentes niveles sobre los herbívoros, ya sea como antinutrientes (inhiben la absorción de nutrientes) o como toxinas. Si bien su producción reduce considerablemente el ataque de los insectos herbívoros, también repercutiría negativamente sobre la planta, ya que la inversión energética necesaria para la producción de estos productos representa un costo que se debía balancear con las probables pérdidas por herbivoría. Por otro lado, este mecanismo inmediatamente generó una presión selectiva sobre las poblaciones de insectos herbívoros, los cuales comenzaron a generar resistencia genéticamente determinada a tales compuestos.[86]
Puesto que la estrategia de defensa química no era suficiente para contrarrestar a los herbívoros debido a que éstos generan resistencia, las plantas fueron desarrollando otras tácticas de protección, en especial del polen y de las semillas, estructuras fuertemente depredadas por los insectos por la gran cantidad de nutrientes que contienen. Para las plantas tal depredación representaba una pérdida muy grande de eficacia biológica ya que impactaba directamente sobre la reproducción de la especie. Por esta razón, las plantas comenzaron a desarrollar conjuntos de hojas modificadas a manera de brácteas para proteger las estructuras reproductivas.[84]
 Las inflorescencias de las Bennettitales son muy similares a flores.
Las inflorescencias de las Bennettitales son muy similares a flores.Las teorías actualmente aceptadas plantean que las flores cíclicas se formaron a partir de la diferenciación y especialización de estos conjuntos de brácteas foliares. La primera evidencia que se encontró para respaldar esta teoría fue el registro fósil de Bennettitales, un grupo de plantas primitivas que durante el Carbonífero originaron prototipos de flores.[84] Este grupo de plantas se originó contemporáneamente con las cicadáceas y las ginkgoáceas (Gimnospermas) y se cree que tuvieron una evolución paralela.[83] [82]
Ya para el Cretácico, las protoflores de Bennettitales habían aumentado su tamaño de 0,5 cm a más de 1012 cm, lo cual incrementó notablemente la amplitud de visitantes florales, entre los cuales se incluían algunos mamíferos pequeños que actuaron también como polinizadores. En estas flores primitivas la polinización era difusa (varios grupos de animales polinizan una misma flor), pero la coevolución produjo mayores grados de especialización y hoy en día la mayoría de las especies de plantas son polinizadas por un solo grupo de insectos.[83]
Si bien el proceso coevolutivo ha llevado al desarrollo de las flores cíclicas en las angiospermas como una respuesta a la presión de herbivoría por parte de los insectos fitófagos, también ha desarrollado estrategias reproductivas adaptadas a las nuevas estructuras. Las flores atraen a los insectos con el néctar o con otro tipo de recompensas florales (como ayuda reproductiva) y esta constante visita por parte de los insectos ha llevado a la selección a favorecer los procesos de polinización entomófila. Puesto que las plantas tienen una movilidad restringida, dependen de características externas para el traslado de los granos de polen de una planta a otra, y de esta manera garantizar la reproducción y la polinización cruzada para evitar la erosión genética por endogamia. Para este objeto, los insectos resultaron ser el vector de polinización más eficiente porque visitan varias flores durante el forrajeo y llevan el polen de una a otra flor.[82]
Mediante los procesos de selección recíproca se han seleccionado favorablemente las flores con características de forma, color y fragancia atractivas para los insectos, puesto que las flores que atraen una mayor cantidad de insectos obtienen una mayor dispersión de su polen y dejan una mayor descendencia, lo cual incrementa la eficacia biológica de la especie. De esta manera se han ido fijando alelos que codifican para ciertos colores, formas y fragancias que han logrado constituir fuertes asociaciones mutualistas (asociaciones en las cuales ambas partes se benefician de la interacción) con los insectos, quienes dispersan el polen a cambio de la recompensa floral. Es esta presión de selección a favor del desarrollo de estrategias reproductivas combinadas la que ha llevado al desarrollo y expresión de tan inmensa variabilidad de flores en las angiospermas, incluso provocando la variación del plan de simetría de las flores con el cambio de organización actinomorfa a cigomorfa, puesto que las flores cigomorfas tienen mayores posibilidades de interacción con los polinizadores.[82] [87]
El origen de las flores hermafroditas y de los modos de reproducción de las angiospermas
 Distilia en Primula vulgaris. Las flores Pin (A) presentan los filamentos de los estambres cortos y los pistilos largos. Las flores Thrum (B) muestran, en cambio, pistilos cortos y filamentos largos. Referencias: 1.- Pétalos, 2.-Sépalos, 3.- Anteras de los estambres y 4.- Pistilo. Las únicas polinizaciones legítimas son las que se producen en cruzamientos Pin x Thrum o Thrum x Pin.
Distilia en Primula vulgaris. Las flores Pin (A) presentan los filamentos de los estambres cortos y los pistilos largos. Las flores Thrum (B) muestran, en cambio, pistilos cortos y filamentos largos. Referencias: 1.- Pétalos, 2.-Sépalos, 3.- Anteras de los estambres y 4.- Pistilo. Las únicas polinizaciones legítimas son las que se producen en cruzamientos Pin x Thrum o Thrum x Pin.La coevolución de las flores de las angiospermas con los insectos polinizadores está gobernada por una serie de principios evolutivos, como fuera señalado por el eminente botánico y evolucionista George Ledyard Stebbins. El principio de la "eficiencia de la polinización" sugiere que la coevolución de las flores con los insectos está basada en la posibilidad de que durante su visita el insecto deposite polen sobre el estigma de la flor mientras, al mismo tiempo, está buscando más polen para alimentarse. Además, los insectos no tendrían incentivo en visitar flores pistiladas que no los recompensaran con polen o con néctar. Por esta razón, las flores deben tener tanto pistilo como anteras para que la visita sea efectiva. Se sigue que la flor hermafrodita, que permite la captación y cesión simultánea de polen, es la estructura más eficaz para sostener en el tiempo la adaptación coordinada entre las plantas y los insectos polinizadores y la razón por la cual es el tipo predominante entre las angiospermas.[88]
La gran ventaja de la zoofilia es, evidentemente, el modo mucho más orientado en como se realiza el transporte del polen por los animales que vuelan de flor en flor. De este modo, fue posible para las plantas reducir significativamente la cantidad necesaria de polen producido, que en el caso de la polinización anemófila tiende a ser muy alto. Además, resultó posible para las angiospermas ocupar hábitats en los que no hay viento, como por ejemplo el interior de los bosques, y vivir en poblaciones poco densas.[14]
En las gimnospermas, el polen no es recibido por un estigma papiloso-viscoso sino que es capturado directamente en la micrópila del óvulo por una "gota polinizante", además, los óvulos descubiertos son mucho más sensibles a las inclemencias del clima. La flor de las angiospermas, entonces, ha permitido un grado mucho mayor de independencia respecto de la humedad ambiental, lo cual, sumado a las ventajas del hermafroditismo y de la zoofilia, explica porqué estas plantas lograron la supremacía frente a las gimnospermas y otros grupos de vegetales que poseían flores unisexuales y anemófilas.[14]
En las flores hermafroditas se produce con facilidad la polinización directa dentro de la misma flor (autogamia) y, con ello, se eleva la consanguinidad o endocría que puede ser sumamente deletérea para muchas especies. Por esta razón, las angiospermas han desarrollado numerosas adaptaciones en sus flores orientadas a favorecer la polinización cruzada (alogamia) e, incluso, hacerla obligatoria. En relación con tales adaptaciones, el estilo y el estigma alcanzaron una importancia decisiva como "filtros" fisilógicos que pueden separar a los granos de polen capturados de acuerdo con la planta que les ha dado origen. En general, no solo impiden la germinación o la formación del tubo polínico en el caso que el polen sea de una especie distinta (incompatibilidad cruzada), sino también, y en la mayoría de las angiospermas, cuando dicho polen procede de la misma planta (autoincompatibilidad).[14] [89]
En distintos grupos de angiospermas se ha llegado, en muchos casos y de modo paralelo, a aumentar el efecto de la autoincompatibilidad por medio de la heterostilia. En las estirpes heterostílicas existen dos o tres tipos diferentes de flores (llamadas morfos). En una planta individual, cada una de las flores comparte el mismo morfo. Estos morfos difieren cualitativamente entre sí en las longitudes de los pistilos y de los filamentos de los estambres. El fenotipo para cada morfo está genéticamente determinado. Cuando una especie presenta dos morfos florales se denomina distílica (ejemplo, Primula), cuando presenta tres, se denomina tristílica (ejemplo, Lythrum). En uno de los morfos, llamado Pin, los estambres son cortos y los pistilos largos (las flores son macrostílicas). En el otro morfo, llamado Thrum, los estambres son largos y los pistilos cortos (flores brevistílicas). Casi todas las especies heterostílicas presentan autoincompatibilidad y los loci responsables de la misma están estrechamente ligados con aquellos genes responsables del polimorfismo floral, por lo que ambos caracteres se heredan en forma conjunta. La distilia está determinada por un solo gen con dos alelos, la tristilia por dos genes con dos alelos cada uno. Como fuera demostrado ya en 1862 por Charles Darwin, solo en el caso de la polinización cruzada entre dos morfos distintos se produce fecundación y fructificación, mientras que si el polen que llega a un estigma procede del mismo morfo, la reacción de autoincompatibilidad impide el crecimiento de los tubos polínicos.[90] [91]
La autoincompatibilidad y la separación espacial de los pistilos y estambres para impedir la autofecundación, como en el caso de la heterostilia, no son los únicos métodos que las angiospermas desarrollaron para favorecer la alogamia. Así, otro procedimiento es el de separar en el tiempo el momento de la maduración de los gametos femeninos y masculinos. En las plantas dicógamas, por ejemplo, los estambres y estigmas alcanzan la madurez para producir y recibir el polen en distintos momentos. Si los estambres maduran antes que los estigmas se dice que hay protandria, en caso contrario, hay protoginia.[89]
La unisexualidad secundaria se las flores en forma de monoecia, dioecia o poligamia se ha producido en las angiospermas sobre todo en relación con la anemofilia, pero permite también evitar la autogamia y la consanguinidad.[14]
En la mayoría de las angiospermas, no obstante, se han originado estirpes derivadas con autogamia facultativa y, finalmente, obligada. Los requisitos necesarios para ello son la desaparición de la autoincompatibilidad y el transporte orientado del polen hacia el estigma de la propia flor, sea por descenso, por movimiento de curvatura de los estambres o por el cierre de la corola. La polinización realizada en la fase de pimpollo conduce a la cleistogamia, en la que las flores no llegan a abrirse, un mecanismo bastante usual en las gramíneas. La autogamia hace posible la producción de descendientes a partir de individuos aislados de cualquier otro ejemplar de la misma especie. Por ello se halla difundida entre las plantas pioneras (las que conquistan nuevos hábitats), las malezas y las especies insulares. Muchas veces la autogamia constituye la única posibilidad de reproducción sexual en ambientes extremos en los que no abundan los animales polinizadores (regiones alpinas, árticas o desérticas). Las especies obligadamente autógamas suelen poseer flores inconspicuas, carentes de fragancia y de néctar y en las que a menudo se ha reducido el tamaño o el número de pétalos y de estambres y ha disminuido la producción de polen o el tamaño de las anteras. Muchas veces la autogamia y la alogamia coexisten en el mismo individuo en un equilibrio ventajoso. Por ejemplo, en muchas especies de la familia de las compuestas al abrirse las flores es posible la fecundación cruzada y solo hacia el final de la antesis, si todavía no ha ocurrido la fecundación, las ramas del estilo se curvan sobre si mismas para tocar el polen de la propia flor y autopolinizarse. En muchas otras especies (Viola, Oxalis, Bromus) se da un equilibrio análogo entre la formación en el mismo individuo de flores normales (llamadas chasmógamas) y otras cleistógamas, muy reducidas. Tales especies, entonces, aseguran la producción de descendientes por medio de la autogamia pero, al mismo tiempo, tratan de generar mayor variabilidad genética por medio de la fecundación cruzada.[14] [89]
Desarrollo floral
Un momento crítico en el ciclo de vida de las plantas con flores es aquel en el que realizan la transición desde la fase juvenil a la fase reproductiva. Para conseguir reproducirse con éxito, es imprescindible, por una parte, que la floración se produzca en sincronía con otros miembros de la misma especie y, por otra parte, que la planta haya alcanzado cierto nivel de crecimiento que le permita sobrellevar el gasto energético implicado en el proceso de formación de flores, frutos y semillas. Por lo tanto, la planta ha de integrar información ambiental, así como endógena, antes de sufrir la transición. Entre los factores externos que más influyen en la floración se encuentran la luz (tanto la intensidad lumínica como la duración del lapso diario de horas de luz o fotoperíodo) y la temperatura. Este último factor desempeña un doble papel. Por un lado, las temperaturas más altas incrementan las tasas de crecimiento y diferenciación en las plantas, por lo que aceleran la transición al período de floración. Por otro lado, muchas plantas necesitan ser expuestas transitoriamente a temperaturas muy bajas durante la fase vegetativa o juvenil para poder florecer, proceso denominado vernalización. Además de estos factores externos, también juegan un papel importante factores endógenos como el reloj circadiano y algunas hormonas como las giberelinas, el ácido salicílico y el ácido abscísico. Así, la planta modelo Arabidopsis thaliana florece más rápidamente cuando los días se alargan (se dice que es una planta de día largo), ya ha sufrido la vernalización (es decir, ha estado expuesta al frío durante el invierno), y la temperatura ambiente incrementa. Es decir, al igual que un sinnúmero de otras especies, la floración de Arabidopsis se induce en las condiciones que se observan en la primavera.[92]
El análisis genético de mutantes con el tiempo de floración alterado ha permitido diseccionar la inducción de la floración en al menos cuatro vías de señalización: la primera se activa cuando los días son largos; la segunda funciona fundamentalmente cuando los días son cortos y depende de las giberelinas; la tercera participa en la regulación de la floración en respuesta a la temperatura ambiental; y la cuarta está implicada en la vernalización.[93]
Cuando la planta llega al estado reproductivo del desarrollo, alguno o todos los meristemas apicales en su vástago cesan de iniciar hojas normales y comienzan a producir partes florales de acuerdo con la secuencia característica de la especie en cuestión y con un número variable de brácteas ubicándose entre las hojas y la flor. En este proceso, los meristemas apicales pasan de tener un crecimiento indeterminado a un tipo de crecimiento determinado, ya que la formación de la flor es generalmente el acontecimiento final de la actividad de un meristema apical. En las plantas anuales, el advenimiento del estado reproductivo significa también el acercamiento a la consumación del ciclo de vida en su totalidad. En las plantas perennes, la floración se repite un número variable de veces que depende de la longevidad de la especie.
Una flor puede originarse en el ápice del vástago principal o en aquellos de las ramas laterales, o en ambos sitios a la vez. En muchas especies, el cambio hacia la floración comprende la formación de una inflorescencia. Un fenómeno observado a menudo durante la iniciación del estado reproductivo es un brusco y rápido alargamiento del eje. Tal crecimiento es particularmente impactante en plantas de hábito en roseta durante el período vegetativo, como por ejemplo las gramíneas o las plantas bulbosas.[94]
Los órganos florales se inician como hojas normales por divisiones periclinales de células localizadas más o menos profundamente por debajo de la protodermis o en la misma protodermis.[95] [96] [97] Las divisiones iniciales periclinales son seguidas por otras, incluyendo anticlinales, y el primordio se convierte en una protuberancia. Los sépalos se parecen más a las hojas normales en su iniciación y desarrollo. El tipo de crecimiento de los pétalos también es más o menos similar al de las hojas. Los estambres se originan como estructuras gruesas y cortas, desarrollándose el filamento subsecuentemente por crecimiento intercalar. El desarrollo del gineceo varía en detalle en relación con la unión de los carpelos entre sí y con otras partes florales. Si los carpelos no están unidos, el primordio del carpelo individual se inicia en un punto por divisiones periclinales en el ápice, las que luego se propagan adaxialmente. El crecimiento hacia arriba, comparable al marginal en una hoja, eleva al carpelo como una estructura en forma de bolsa. En los ovarios sincárpicos los carpelos se originan como primordios individuales o conjuntamente como una unidad estructural.[94]
Biología molecular del desarrollo floral
 Diagrama de la expresión diferencial de los genes A, B y C en los distintos verticilos de una flor de Arabidopsis.
Diagrama de la expresión diferencial de los genes A, B y C en los distintos verticilos de una flor de Arabidopsis.La definición de flor brindada al principio de este artículo se encuadra dentro del contexto de la denominada "teoría foliar", la cual sugiere que todas las piezas florales son hojas estructuralmente modificadas y funcionalmente especializadas en la reproducción o la protección. La formulación de esta teoría comenzó en 1790 cuando Goethe publicó su obra "Un intento para interpretar la metamorfosis de las plantas" ("Versuch die Metamorphose der Pflanzen zu erklaren").[98] En este libro, que consistía en 123 párrafos numerados, Goethe escribió:
"...también deberíamos decir que un estambre es un pétalo contraído, como que un pétalo es un estambre en expansión; o que el sépalo es una hoja caulinar contraída aproximándose a cierto estado de refinamiento, como que una hoja caulinar es un sépalo expandido por el influjo de jugos más bastos".[99]Casi 200 años después de esta publicación, la biología molecular y la biología evolutiva del desarrollo (o informalmente evo-devo, del inglés evolutionary developmental biology) han inventado las herramientas para decodificar la estructura molecular básica del desarrollo floral. Uno de los hitos de estas disciplinas ha sido la descripción del denominado modelo "ABC", un modelo genético simple y elegante para explicar la determinación de la identidad de los órganos florales,[100] cuya confirmación ha sido el descubrimiento de la profunda homología y conservación de los genes involucrados en el modelo, pertenecientes a taxa que han divergido hace varios millones de años[101] [102]
El modelo ABC del desarrollo floral fue propuesto como modelo explicativo del conjunto de mecanismos genéticos que dan lugar al establecimiento de identidades de órgano en Arabidopsis thaliana, como representante de las rósidas, y de Antirrhinum majus, perteneciente al linaje de las astéridas. Ambas especies poseen cuatro verticilos (sépalos, pétalos, estambres y carpelos), definidos por la expresión diferencial en cada verticilo de una serie de genes homeóticos. El modelo ABC establece que la identidad de los órganos presentes en esos cuatro verticilos florales es consecuencia de la interacción de por lo menos tres tipos de productos génicos de funciones distintas. De este modo, los sépalos se caracterizan por la expresión de genes A exclusivamente; en los pétalos, en cambio, se coexpresan los genes A y B; en los estambres, los genes de función B y C establecen su identidad; y en los carpelos, únicamente se requiere de la actividad de genes de función C. Además, los genes tipo A y C son antagonistas recíprocos.[103]
El hecho de que estos genes homeóticos determinan la identidad de órgano se pone de manifiesto cuando un gen representante de una determinada función, por ejemplo la A, no se expresa. Así, en Arabidopsis dicha pérdida produce la aparición de una flor compuesta por un verticilo de carpelos, otro de estambres, otro de estambres, y otro de carpelos.[103] Se ha propuesto, además de las mencionadas funciones A, B y C, la existencia de unas suplementarias: la D y la E. La función D especificaría la identidad de óvulo, como una función reproductiva separada del desarrollo de los carpelos y posterior en aparición a la determinación de estos.[104] La función E correspondería a un requisito fisiológico propio de todos los verticilos florales, aunque, en inicio, fue descrita como necesaria para el desarrollo de los tres verticilos más internos (función E sensu stricto).[105] No obstante, su acepción sensu lato sugiere que es requerido en los cuatro verticilos.[106] De este modo, cuando se pierde la función D, los óvulos pasan a ser estructuras semejantes a hojas, y cuando se pierde la función E sensu stricto, los órganos florales de los tres verticilos más internos se transforman en sépalos,[105] mientras que, de perderse la función E sensu lato, todos los verticilos son semejantes a hojas.[106] [107]
El modelo ABC, entonces, explica los aspectos más importantes del desarrollo floral, desde los más teóricos (una puesta al día de las ideas de Goethe) hasta los más prácticos, tales como la manipulación de los genes ABC para producir flores con cualquier tipo de combinación de piezas florales.[108] [98]
Simbolismo
En todas las culturas alrededor del mundo, hasta donde se tienen registros históricos, las flores han provisto información emocional entre los seres humanos. En las cuevas de neandertales se ha hallado polen (la parte de la flor más difícil de corromper a través del tiempo) y se ha sugerido que las flores eran parte del ceremonial para rendir homenaje a sus difuntos,[109] si bien tal significado todavía está en discusión.[110]
En la antigüedad las flores no sólo eran consideradas fuente de placer para dioses y hombres, y motivo de inspiración para artistas y poetas, sino que poseían significado de trascendencia mística y cósmica. En el Japón, donde se cultiva el arte del arreglo floral (ikebana), la flor se considera el resumen del ciclo vital de todas las criaturas y su efímera duración. La flor que Buda exhibe, en ocasiones, sustituye toda palabra y enseña: es la imagen explícita de la perfección a alcanzar, de la iluminación y la expresión de lo inexpresable. En Grecia antigua por estar asociadas a las mariposas se les identificaba con las almas de los muertos. En el Taoísmo la flor de oro que emerge a partir de la coronilla es símbolo de la máxima iluminación espiritual.[111] En las sociedades americanas prehispánicas, por ejemplo, las flores ofrecieron una amplia gama de significados. Las antiguas representaciones de las flores no eran meramente decorativas, sino que formaban parte de un simbolismo religioso. Las flores de cuatro pétalos, por ejemplo, han tenido un significado polifacético en las culturas antiguas y actuales de Mesoamérica, y es uno de los símbolos persistentes en la mente y en el lenguaje de sus habitantes. Hay numerosas representaciones de flores de cuatro pétalos en Teotihuacán (estado de México, México) y en otros sitios tales como en Tlalancaleca (estado de Puebla, México). Las flores teotihuacanas también formaron parte de la iconografía de la pintura mural, a veces aludiendo a un lugar paradisíaco, otras veces refiriéndose al canto y a lo bello de las palabras[112] A algunas se les otorgó un carácter sagrado y sirvieron para fines ceremoniales y mágicos, como sucedió con el "nardo" u «omixóchitl» (Polianthes tuberosa), el "pericón" o «yauhtli» (Tagetes lucida) y el «cempoaxóchitl» (Tagetes erecta), hoy conocida en México como "cempasúchil" o "flor de muertos", flores que por su perfume tan fuerte han servido como medio de comunicación o atracción de los seres sobrenaturales, o como protección contra ellos.[113] Los lirios (Lilium), por ejemplo, han sido utilizados como símbolo de pureza durante cientos de años.
 Lirios en un fresco de la edad de bronce que representa un paisaje primaveral. Excavaciones de Akrotriri, Santorini, Grecia.
Lirios en un fresco de la edad de bronce que representa un paisaje primaveral. Excavaciones de Akrotriri, Santorini, Grecia.Existen pinturas en palacios de Grecia de más de 3.000 años de antigüedad que atestiguan ese uso. Esta asociación entre la pureza y ciertos lirios pasó más tarde a la cristiandad, donde a la Virgen María se la representaba con flores de lirio (Lilium candidum, "Madonna Lily" en inglés) en sus brazos. El símbolo de la flor de lis estuvo originalmente basado en la flor de una especie de iris (Iris pseudacorus) y aparecía en pinturas religiosas egipcias e indias mucho antes que fuera adoptado como emblema de los reyes de Francia desde el Siglo V. Las flores han servido no solo como símbolos religiosos sino también del poder real desde hace miles de años y han sido admiradas y utilizadas por su belleza desde tiempos inmemoriales y por muchas civilizaciones.[114] [115]
Además de por su belleza, simbolismo religioso y su asociación con el poder, las flores también han sido utilizadas como medio de comunicación. El romanticismo de fines del siglo XIX, rescató el llamado lenguaje de las flores que fuera desarrollado en el siglo previo por los burgueses alemanes, con el objeto de comunicar mensajes específicos a amigos y amantes, para lo que se escribieron libros que explicaban el significado de cada flor. Ejemplos de este "simbolismo floral" son: lila, «corramos al altar antes que la juventud se acabe»; rosa, «bella como una flor»; altramuz, «encanto celestial e inteligencia hallé en tu corazón»; girasol, «tu amor es para mi vida lo que el sol para esta flor»; cebolla, «me repugnas»; saúco, «cada vez te siento más fría»; pétalo de rosa roja, «sí»; de rosa blanca, «no». El profundo y cautivante significado que la flor representa universalmente no ha sido dejado de lado por novelistas y poetas de todos los tiempos quienes se han valido de ella para titular muchas de sus obras: Las flores del mal de Charles Baudelaire, Por el sendero de las muchachas en flor de Marcel Proust, El nombre de la rosa de Umberto Eco, Doña Flor y sus dos maridos de Jorge Amado, entre decenas de ejemplos.[111]
Respuestas socio-emocionales a las flores
En la actualidad, se espera que las flores transmitan simpatía, contrición o culpa, romance o acercamiento sexual y celebración, desde orgullo hasta alegría.[116] Las flores también se utilizan para expresar sentimientos religiosos y en algunas religiones se considera una ruta directa a la comunicación espiritual.[117] Además, por supuesto, muchas flores se utilizan para adorno personal, tanto los pimpollos en sí mismos como sus esencias bajo la forma de perfumes. De hecho, la gran mayoría de las fragancias comerciales para uso personal tienen una nota floral. Independientemente de algunos usos básicos, tales como las flores de uso culinario o las de uso medicinal, la mayoría de las especies que el ser humano cultiva por sus flores no tienen otro propósito más que el emocional. Un uso que, para poner un ejemplo, supone un mercado de casi 5.000.000.000 de dólares en Estados Unidos de América en el año 2001.[118]
Si bien desde un análisis superficial se puede argüir que las flores son deseables para los humanos debido a asociaciones aprendidas con eventos sociales, la universalidad del uso de las flores a través de distintas culturas, etapas históricas y la ausencia de un sustituto adecuado para los muchos usos que hacemos de ellas, sugiere que debe existir algo más que esta simple asociación. De hecho, por más de 5.000 años los seres humanos hemos cultivado flores a pesar de que no existe una recompensa conocida para este comportamiento costoso desde el punto de vista del trabajo y la energía que supone realizarlo. En tres estudios diferentes se ha demostrado que las flores son un poderoso inductor de emociones positivas en nuestra especie. En el primer estudio, las flores siempre suscitaban la sonrisa de Duchenne inmediatamente después de obsequiárselas a mujeres,[c]La plantilla {{Note label}} está obsoleta, véase el nuevo sistema de referencias. quienes informaron que habían tenido mejor humor desde ese momento hasta tres días más tarde, cuando fueron encuestadas. En un segundo estudio, una flor entregada a mujeres o varones en un ascensor produjo una conducta social más positiva que otros estímulos. En un tercer estudio, las flores entregadas a personas de más de 55 años suscitaron un humor más positivo y una mejora en la memoria. Las flores tuvieron un efecto inmediato y a largo plazo sobre las reacciones emocionales, humor, conducta social, e incluso la memoria, tanto en mujeres como en varones. De hecho, se ha formulado la hipótesis de que las flores pueden influenciar o iniciar el comportamiento socio-emocional de los seres humanos ya que estarían adaptadas a un nicho emocional en nuestra especie.[118]
Floricultura
 Una rosa azul, obtenida mediante colorantes artificiales. Este tipo de rosas se está tratando de obtener por medio de ingeniería genética.
Una rosa azul, obtenida mediante colorantes artificiales. Este tipo de rosas se está tratando de obtener por medio de ingeniería genética.La floricultura es el arte y la técnica del cultivo de plantas para obtener flores y su comercialización en florerías y viveros. Pueden establecerse dos tipos fundamentales de explotaciones, la particular y la comercial. La primera, asociada con la jardinería, persigue como objetivo el cultivo de plantas con flores por su belleza y por satisfacción anímica, y la llevan a cabo desde aficionados hasta entusiastas coleccionistas que se han especializado en una especie o grupos de especies de plantas. La floricultura comercial, en cambio, se refiere a los cultivos de plantas para obtener flores que persiguen una finalidad de lucro. Sus productos pueden estar destinados a la ornamentación, a la industria o a la medicina e involucran otras actividades comerciales estrechamente relacionadas, tales como la producción de semillas, de bulbos y de una infinidad de elementos imprescindibles para esta actividad intensiva, que van desde los fertilizantes y agroquímicos (insecticidas, fungicidas y herbicidas) hasta las macetas y sustratos para el cultivo.[119] [120] [121]
El mercado internacional de flor cortada está fundamentalmente centrado en tres mercados consumidores: Estados Unidos, la Unión Europea y Japón, con un valor de facturación de 955, 6.500 y 3.800 millones de euros respectivamente. Cada uno de esos mercados produce un alto porcentaje de su demanda de flor cortada, pero además, importan una considerable cantidad desde una diversidad de países. Así los Estados Unidos importan el 59% de sus requerimientos, la UE el 10% y Japón solo el 6%. Los principales proveedores del mercado estadounidense son Colombia, Ecuador y los Países Bajos. Para la UE, los principales proveedores son Kenia, Israel y Colombia y para el mercado japonés, Tailandia, Colombia y los Países Bajos. Las especies más destacadas en los mercados internacionales son las rosas, los claveles, los crisantemos y varias especies de plantas bulbosas, como los tulipanes y las azucenas. Los Países Bajos constituyen el mercado concentrador más importante del mundo, y registran más del 50 por ciento del total de las importaciones y el 85 por ciento de las exportaciones de flores de la Unión Europea.[122]
La causa del crecimiento sostenido del mercado de flor cortada en las últimas décadas es, básicamente, el cambio en las pautas de consumo de la población con ingresos medios y altos, que utiliza este producto como elemento decorativo del hogar, regalos empresariales y para solaz personal. Una de las consecuencias del crecimiento de los mercados de flor de corte es la cuantiosa inversión que muchas empresas e instituciones realizan para obtener nuevas variedades de flores, ya sea por medio de mejoramiento genético clásico o por medio de sofisticadas técnicas de ingeniería genética[123]
Véase también
- Angiospermae
- Botánica
- Desarrollo floral
- Flora
- Síndrome floral
- Autoincompatibilidad
Notas
a. ↑ La plantilla {{Note label}} está obsoleta, véase el nuevo sistema de referencias. La definición más concisa de flor es "un braquiblasto que lleva los antófilos", es decir un eje corto de crecimiento determinado que lleva las hojas especializadas en la función de reproducción (estambres y carpelos) y de protección de los gametos (sépalos y pétalos). "Crecimiento determinado" quiere decir que el meristema apical que da origen a la flor cesa su actividad inmediatamente después que ha producido todas las partes florales.[2]
b. ↑ La plantilla {{Note label}} está obsoleta, véase el nuevo sistema de referencias. Un ejemplo de esto es la lluvia de azufre en la época de floración de las coníferas.
c. ↑ La plantilla {{Note label}} está obsoleta, véase el nuevo sistema de referencias. Muchos investigadores han sugerido que la sonrisa de Duchenne indica una emoción espontánea y genuina ya que la mayor parte de las personas no pueden contraer a voluntad los músculos involucrados en la expresión facial de este tipo de sonrisa.[124]
Referencias
- ↑ University of Hamburg. Department of Biology. Botany on line.Flower Morphology of Plants. Consultado el 5 de abril de 2009;
- ↑ a b c d e f g h i j k Font Quer, P. (1982). Diccionario de Botánica. 8ª reimpresión. Barcelona: Editorial Labor, S. A. ISBN 84-335-5804-8.
- ↑ a b c Damerval, C.; Nadot, S. 2007. Evolution of Perianth and Stamen Characteristics with Respect to Floral Symmetry in Ranunculales. Ann. Bot. 100: 631-640.
- ↑ Barnard, G. 1961. The interpretation of the angiosperm flower. Aust. J. Sci. 24: 64-72.
- ↑ Carlquist, S. 1969. Towards acceptable evolutionary interpretations of floral anatomy. Phytomorphology 19:332-362.
- ↑ Foster, A.S. & Gifford, E.M. 1974. Comparative morphology of seed plants. San Francisco, Freeman & Co.
- ↑ University of Hamburg. Department of Biology. Botanic on line.Flower Morphology of Plants. Consultado el 5 de abril de 2009.
- ↑ J. Marquez-Guzman, M. Engleman, A. Martinez-Mena, E. Martinez and C. Ramos. Anatomia Reproductiva de Lacandonia schismatica (Lacandoniaceae). Annals of the Missouri Botanical Garden, Vol. 76, No. 1 (1989), pp. 124-127
- ↑ Ronse De Craene LP, Soltis PS, Soltis DE. 2003. Evolution of floral structures in basal angiosperms. International Journal of Plant Sciences 164.
- ↑ Endress PK. Origins of flower morphology. In: The character concept in evolutionary biology. Wagner GP, ed. (2001) San Diego, CA: Academic Press. 493–510.
- ↑ Endress PK. Patterns of floral construction in ontogeny and phylogeny. Biological Journal of the Linnean Society (1990) 39:153–175
- ↑ Endress PK, Doyle JA. Floral phyllotaxis in basal angiosperms: development and evolution. Current Opinion in Plant Biology (2007) 10:52–57
- ↑ González, A.M. «Flor, disposición de las piezas florales» (en español). Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Consultado el 10 de abril de 2009.
- ↑ a b c d e f g h i j k l m n ñ o Strassburger, E. 1994. Tratado de Botánica. 8va. edición. Omega, Barcelona, 1088 p. ISBN 84-7102-990-1
- ↑ a b c González, A.M.. «Flor, simetría y sexualidad floral» (en español). Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Consultado el 10 de abril de 2009.
- ↑ Judd, W. S.; C. S. Campbell, E. A. Kellogg, P. F. Stevens, M. J. Donoghue (2007). «Cannaceae». Plant Systematics: A Phylogenetic Approach, Third edition.. Sunderland, Massachusetts: Sinauer Associates. pp. 306. ISBN 978-0-87893-407-2.
- ↑ Simpson, Michael G. (2005). «Cannaceae». Plant Systematics.. Elsevier Inc.. pp. 198-203. ISBN 0-12-644460-9 ISBN 978-0-12-644460-5.
- ↑ a b Endress PK. Symmetry in flowers: diversity and evolution. International Journal of Plant Sciences (1999) 160:S3–S23
- ↑ Rudall PJ, Bateman RM. Evolution of zygomorphy in monocot flowers: iterative patterns and developmental constraints. New Phytologist (2004) 162:25–44
- ↑ González, A.M. «Perianto» (en español). Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Consultado el 10 de abril de 2009.
- ↑ a b Font Quer, P. (1960). Botánica Pintoresca.. Barcelona: Editorial Ramón Sopena.
- ↑ González, A.M. «Perianto, cáliz» (en español). Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Consultado el 10 de abril de 2009.
- ↑ a b c d e f Gola, G., Negri, G. y Cappeletti, C. 1965. Tratado de Botánica. 2da. edición. Editorial Labor S.A., Barcelona, 1110 p.
- ↑ a b c d e f g h i j k Valla, J.J. 2005. Botánica: morfología de las plantas superiores. Buenos Aires, Hemisferio Sur. ISBN 950-504-378-3
- ↑ González, A.M. «Perianto, corola» (en español). Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Consultado el 10 de abril de 2009.
- ↑ González, A.M. «Anatomía Floral. Cáliz y Corola» (en español). Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Consultado el 10 de abril de 2009.
- ↑ González, A.M. «Perianto» (en español). Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Consultado el 10 de abril de 2009.
- ↑ Richtmyer, F. K. 1923. The reflection of ultraviolet by flowers. Journal of the Optical Society of America, vol. 7, issue 2, p.151
- ↑ Lutz, F. E. 1924. Hunting stingless bees where east seems to be west. Nat. Hist 24:494–508.
- ↑ Daumer, K. 1958. Blumenfarben, wie sie die Bienen sehen. Journal of Comparative Physiology A: Neuroethology, Sensory, Neural, and Behavioral Physiology 41:49-110.
- ↑ R. S. Thompson, D. Jacques, E. Haslam and R. J. N. Tanner. Part I. Introduction; the isolation, structure, and distribution in nature of plant procyanidins. J Plant proanthocyanidins. Chem. Soc., Perkin Trans. 1, 1972, 1387 - 1399, DOI: 10.1039/P19720001387
- ↑ Frederick H. Utech, and Shoichi Kawano.1975. Spectral polymorphisms in angiosperm flowers determined by differential ultraviolet reflectance. Journal of Plant Research 88:9-30.
- ↑ Strassburger, E. 1994. Tratado de Botánica. 8va. edición. Omega, Barcelona, 1088 p.
- ↑ Chittka, L.; Shmida, A.; Troje, N.; Menzel, R. (1994). «Ultraviolet as a component of flower reflections, and the colour perception of Hymenoptera». Vision Research 34 (11): pp. pp. 1489–1508. http://www.biomotionlab.ca/Text/chittka_VR_94.pdf. Consultado el 3 de mayo de 2009.
- ↑ a b c González, A.M. «Androceo» (en español). Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Consultado el 14 de abril de 2009.
- ↑ González, A.M.. «Androceo, número de estambres» (en español). Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Consultado el 14 de abril de 2009.
- ↑ a b c González, A.M.. «Flor, estilo y estigma» (en español). Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Consultado el 15 de abril de 2009.
- ↑ Sassen M. M. A (1974). «The stylar transmitting tissue». Acta Botanica Neerlandica (23): pp. 99-108.
- ↑ Jensen W. A. & D. B. Fisher (1970). «Cotton embryogenesis: the pollen tube in the stigma and style». Protoplasma (69): pp. 215-235.
- ↑ C. Labarca, M. Kroh, and F. Loewus (2002). «The Composition of Stigmatic Exudate from Lilium longiflorum: Labeling Studies with Myo-inositol, D-Glucose, and L-Proline». Plant Physiol (46): pp. 150-156. http://www.plantphysiol.org/cgi/content/abstract/46/1/150?ijkey=440d892be29dca63e25cf3e2ccf0239c4f3d7c1e&keytype2=tf_ipsecsha.
- ↑ Eames A. J. (1961). «Morphology of the angiosperms». McGraw-Hill (Nueva York, Nueva York, EE. UU.) (69): pp. 215-235.
- ↑ Kocyan A. & P. K. Endress (2001). «Floral structure and development of Apostasia and Neuwiedia (Apostasioideae) and their relationships to other Orchidaceae.». International Journal of Plant Sciences (162): pp. 847-867.
- ↑ Heslop Harrison & K. R. Shivanna. 1997. The receptive surface of the angiosperm stigma. Ann. Bot. 41: 1233-1258
- ↑ Takhtajan, A. 1991. Evolutionary Trends in Flowering Plants. Columbia University Press, New York, ISBN 0-231-07328.
- ↑ a b González, A.M.. «Flor, óvulo y placentación» (en español). Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Consultado el 20 de abril de 2009.
- ↑ González, A.M.. «Flor, fórmula y diagrama floral» (en español). Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Consultado el 10 de abril de 2009.
- ↑ University of Berkeley. Department of evolutionary biology. What makes a petal a petal?. Consultado el 14 de abril de 2009.
- ↑ a b c Dimitri, M. 1987. Enciclopedia Argentina de Agricultura y Jardinería. Editorial ACME, Buenos Aires.
- ↑ Günter Gerlach. 2003.La subtribu Stanhopeinae: sus notables mecanismos de polinización, la química de sus aromas florales e implicaciones en sistemática y taxonomía Lankesteriana 7: 104-106.
- ↑ González, A.M.. «Reproducción de las plantas con flores. Evolución floral» (en español). Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Consultado el 10 de abril de 2009.
- ↑ Robert A. Raguso. 2008. Wake Up and Smell the Roses: The Ecology and Evolution of Floral Scent. Annual Review of Ecology, Evolution, and Systematics 39, 549-569
- ↑ González, A.M.. «Reproducción de las plantas con flores. Atracción de los agentes polinizadores» (en español). Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Consultado el 10 de abril de 2009.
- ↑ a b Raisman, J. y González, A.M.. «Evolución floral» (en español). Hipertextos del área de la biología. Reproducción de las plantas con flores.. Argentina: Universidad Nacional del Nordeste. Consultado el 1 de mayo de 2009.
- ↑ Barth, F. (1985) Insects and Flowers. The Biology of a Partnership. Princeton University Press. Princeton, NJ. ISBN 0-691-08368-1
- ↑ L. van der Pijl. 1961. Ecological Aspects of Flower Evolution. II. Zoophilous Flower Classes. Evolution, Vol. 15, No. 1, pp. 44-59
- ↑ Proctor, M., Yeo, P. & Lack, A. (1996). The Natural History of Pollination. Timber Press, Portland, OR. ISBN 0-88192-352-4
- ↑ Meeuse, B. & Morris, S. (1984) The Sex Life of Flowers. The Rainbird Publishing Group ltd. London. ISBN 0-87196-907-6
- ↑ Pijl, L. van der y C. H. Dodson. 1969. Orchid flowers. Their pollination and evolution. University of Miami Press. Coral Gables. pp. 101-122.
- ↑ Chase, M. W. 1985. Pollination of Pleurothallis endosthachys. Am. Orch. Soc. Bull. 54:431-434
- ↑ Johnson, S. D. y T. J. Edwards. 2000. The structure and function of orchid pollinaria. Pl. Syst. Evol. 222: 243-269.
- ↑ Borba, E. L., J. M. Felix, V. N. Solferini y J. Semir. 2001b. Fly-pollinated Pleurothallis (Orchidaceae) species have high phenetic variability: evidence from isozyme markers.Am. J. Bot. 88: 419-428.
- ↑ Borba, E. L. y J. Semir. 2001. Pollinator specificity and convergence in fly pollinated Pleurothallis (Orchidaceae) species: a multiple population approach. Ann. Bot. 88: 75-88.
- ↑ a b Perez, F., Arroyo, M., Medel, R. & M. Herskovitz. 2006. Ancestral reconstruction of flower morphology and pollination systems in Schizanthus (Solanaceae). American Journal of Botany 93(7): 1029–1038.
- ↑ Raisman, J. y González, A.M. «El gametofito masculino» (en español). Hipertextos del área de la biología. Reproducción de las plantas con flores.. Argentina: Universidad Nacional del Nordeste. Consultado el 1 de mayo de 2009.
- ↑ Raisman, J. y González, A.M.. «El gametofito femenino» (en español). Hipertextos del área de la biología. Reproducción de las plantas con flores.. Argentina: Universidad Nacional del Nordeste. Consultado el 1 de mayo de 2009.
- ↑ Raisman, J. y González, A.M.. «Doble fecundación» (en español). Hipertextos del área de la biología. Reproducción de las plantas con flores.. Argentina: Universidad Nacional del Nordeste. Consultado el 1 de mayo de 2009.
- ↑ Cocucci, A. E. 1995. Proceso sexual en angiospermas. Academia Nacional de Ciencias, Córdoba (Argentina), 46p.
- ↑ Ge Sun, David L. Dilcher, Shaoling Zheng, Zhekun Zhou. 1998. In Search of the First Flower: A Jurassic Angiosperm, Archaefructus, from Northeast China. Science Vol. 282. no. 5394, pp. 1692 - 1695
- ↑ G. Sun, Q. Ji, D.L. Dilcher et al., Archaefructaceae, a New Basal Angiosperm Family. Science 296, 899.
- ↑ E.M. Friis, K.R. Pedersen, and P.R. Crane. 2005. Cretaceous angiosperm flowers: Innovation and evolution in plant reproduction Curr. Opin. Plant Biol. 8, 5
- ↑ E.M. Friis, K.R. Pedersen, and P.R. Crane, Palaeogeogr. Palaeoclim. Palaeoecol. 232 (2), 251 (2006).
- ↑ D.L. Dilcher, G. Sun, Q. Ji et al., 2007. An early infructescence Hyrcantha decussata (comb. nov.) from the Yixian Formation in northeastern China. Proc. Nat. Acad. Sci. USA 104 (22), 9370.
- ↑ Friis, E.M., J.A. Doyle, P.K. Endress & Q. Leng 2003. Archaefructus. Angiosperm precursor or specialized early angiosperm? Trends in Plant Sciences 8: 369–373.
- ↑ Ji, Q., H. Li, L.M. Bowe, Y. Liu & D.W. Taylor 2004. Early Cretaceous Archaefructus eoflora sp. nov. with Bisexual Flowers from Beipiao, Western Liaoning, China.PDF (3.11 MiB) Acta Geologica Sinica 78(4): 883–896.
- ↑ Xin Wang, Shaolin Zheng- 2007. The Earliest Perfect Flower: a perfect flower typical of angiosperms, including androecium, gynoecium and tepals, from western Liaoning, China marks the earliest record of undoubted flowers in the early Cretaceous (>125 Ma). Nature Precedings : hdl:10101/npre.2007.1320.1 : Posted 15 Nov 2007
- ↑ Sun, G.; Ji, Q.; Dilcher, D.L.; Zheng, S.; Nixon, K.C.; Wang, X. (2002). «Archaefructaceae, a New Basal Angiosperm Family». Science 296 (5569): pp. 899. doi:.
- ↑ a b Frohlich, M.W.; Chase, M.W. (2007). «After a dozen years of progress the origin of angiosperms is still a great mystery.». Nature 450 (7173): pp. 1184–9. doi:.
- ↑ a b Wing, S.L.; Boucher, L.D. (1998). «Ecological Aspects Of The Cretaceous Flowering Plant Radiation». Annual Reviews in Earth and Planetary Sciences 26 (1): pp. 379–421. doi:.
- ↑ a b Feild, T.S.; Arens, N.C.; Doyle, J.A.; Dawson, T.E.; Donoghue, M.J. (2004). «Dark and disturbed: a new image of early angiosperm ecology» (abstract). Paleobiology 30 (1): pp. 82–107. http://paleobiol.geoscienceworld.org/cgi/content/abstract/30/1/82.
- ↑ Scagel R., Bandoni R., Rouse G., Schofield W., Stein J. & T. Taylor. (1977). El Reino vegetal: los grupos de plantas y sus relaciones evolutivas. Ediciones Omega, Barcelona, pp 570, 573-575.
- ↑ L. van der Pijl. 1960. Ecological Aspects of Flower Evolution. I. Phyletic Evolution. Evolution, Vol. 14, No. 4, pp. 403-416
- ↑ a b c d e Fontúrbel, F. Rol de la coevolución planta–insecto en la evolución de las flores cíclicas en las angiospermas. Ciencia Abierta (Chile). Consultadoel 10 de abril de 2009.
- ↑ a b c d Fontúrbel, F. & D. Mondaca. (2000). Coevolución insecto–planta en la polinización. Revista Estudiantil de Biología, 1 (1): 18–27
- ↑ a b c d Jolivet, P. (1992). Insects and plants: parallel evolution and adaptations (2ª edición). Florida: Sandhill Crane Press. pp. 157–163. ISBN 9781877743108.
- ↑ González, A.M. & J. Raisman. «Compuestos secundarios de las plantas» (en español). Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Consultado el 1 de abril de 2009.
- ↑ Paré, P. & J. Tumlinson. (1999). Plant volatiles as a defense against insect herbivores. Plant Physiology, October, 121: 325–331.
- ↑ Friis EM, Endress PK. 1996 I Flower evolution. Prog. Bot 57. 253-280
- ↑ Stebbins, G.L. (1970). «Adaptive radiation of reproductive characteristics in angiosperms, I: pollination mechanisms». Annual Review of Ecology and Systematics 1 (1): pp. pp. 307–326. doi:.
- ↑ a b c Frankel, R. & Galun, E. (1977). Pollination mechanisms, reproduction, and plant breeding. Serie: Monographs on Theoretical and Applied Genetics no. 2.. Berlín: Springer-Verlag. ISBN 0387079343.
- ↑ Darwin, C. R. 1862. On the two forms, or dimorphic condition, in the species of Primula, and on their remarkable sexual relations. Journal of the Proceedings of the Linnean Society of London (Botany) 6: 77-96. Reimpresión del trabajo original
- ↑ Ganders, F. R. (1979). "The biology of heterostyly." New Zealand Journal of Botany 17: 607-635.
- ↑ Blázquez, Miguel A.. «Regulación del tiempo de floración por luz y temperatura» (en español). BioPress.net. Consultado el 8 de mayo de 2009.
- ↑ Simpson GG, Gendall AR & Dean C. 1999. When to switch to flowering. Annu. Rev. Cell Dev. Biol. 15: 519-50.
- ↑ a b ESAU, K. 1988. Anatomía de las plantas con semilla. Ed. Hemisferio Sur. ISBN 950-004-233-9
- ↑ Tepfer, S. 1953. Floral anatomy and ontogeny in Aquilegia formosa and Ranunculus repens. Univ. Calif. Pubs. Bot. 25:513-648.
- ↑ Tucker, S.C. 1959. Ontoeny of the inflorescence and the flower of Drymis winteri. Univ. Calif. Pubs. Bot. 30:257-336.
- ↑ Barnard, C. 1957. Floral histogenesis in the monocotyledons. I. The Gramineae, Aust. J. Bot. 5:115-128.
- ↑ a b Dornelas, M.C.; Dornelas, O. 2005. Da folha à flor: revisitando os conceitos de Goethe sobre a ¨metamorfose¨ das plantas. Braz. J. Plant Physiol., Londrina, v. 17, n. 4,
- ↑ Goethe J.W. von (1790) Versuch die Metamorphose der Pflanzen zu erklaren. Gotha, Ettlinger; párrafo 120."
- ↑ Coen ES, Meyerowitz EM (1991) The war of the whorls: genetic interactions controlling flower development. Nature 353:31-37.
- ↑ Theissen G, Becker A, Di Rosa A, Kanno A, Kim JT, Munster T, Winter KU, Saedler H. (2000) A short history of MADS-box genes in plants. Plant Mol. Biol. 42:115-149.
- ↑ Irish VF, Litt A (2005) Flower development and evolution: gene duplication, diversification and redeployment. Curr. Opin. Gen. Dev. 15:1-7.
- ↑ a b Bowman, J.L et al.. Expression of the Arabidopsis floral homeotic gene AGAMOUS is restricted to specific cell types late in flower development. Plant Cell 3, 749-758
- ↑ L Colombo, J Franken, E Koetje, J van Went, H J Dons, G C Angenent, et A J van Tunen. The petunia MADS box gene FBP11 determines ovule identity. Plant Cell. 1995 November; 7(11): 1859–1868.
- ↑ a b Soraya Pelaz, Gary S. Ditta, Elvira Baumann, Ellen Wisman and Martin F. Yanofsky. B and C floral organ identity functions require SEPALLATA MADS-box genes Nature 405, 200-203 (11 May 2000) | doi:10.1038/35012103
- ↑ a b Gary Ditta, Anusak Pinyopich, Pedro Robles, Soraya Pelaz and Martin F. Yanofsky (9 de noviembre de 2004). «The SEP4 Gene of Arabidopsis thaliana Functions in Floral Organ and Meristem Identity». Current Biology (21): pp. pp. 1935-1940. http://www.sciencedirect.com/science?_ob=ArticleURL&_udi=B6VRT-4DRMNRG-T&_user=10&_rdoc=1&_fmt=&_orig=search&_sort=d&view=c&_acct=C000050221&_version=1&_urlVersion=0&_userid=10&md5=0e4f4dc169aec5f88d3c985d7e7f352a.
- ↑ Weigel D, Meyerowitz EM (1994) The ABCs of floral homeotic genes. Cell 78:203-209.
- ↑ Dornelas MC (2000) Construindo flores: O controle molecular da arquitetura floral. Biotecnol. Ciênc. Desenvolv. 12:44-46.
- ↑ Solecki, R. S. (1971). Shanidar: The First Flower People. NY: Knopf.
- ↑ Sommer, J. D. (1999). The Shanidar IV "Flower Burial": A Reevaluation of Neanderthal Burial Ritual, Cambridge Archaeological Journal, 9: 127-129.
- ↑ a b Molina Prieto, L.F, Uribe Botero, E. y Osorio Olarte, J. Las flores de los jardines de Santa Fé de Bogotá. Publicación digital en la página web de la Biblioteca Luis Ángel Arango del Banco de la República. Consultado el 29 de abril de 2009.
- ↑ Heydn, D. 1983. Mitología y simbolismo de la flora en el México prehispánico, UNAM, México.
- ↑ Ana María L. Velasco Lozano, Debra Nagao. 2003. Mitología y simbolismo de las flores. Arqueología mexicana 4:29-35
- ↑ Rosella Rossi. 1990. Guía de Bulbos. Grijalbo. Barcelona.
- ↑ Hessayon, D.G. The Bulb Expert. Transworld Publishers Ltd. Londres, 1999
- ↑ Heilmeyer, M. (2001). The Language of Flowers: Symbols and Myths. New York, Prestel USA, 59pp. ISBN 3791323962
- ↑ Stenta, N. (1930). From other lands: the use of flowers in the spirit of the liturgy. Orate Fratres, 4(11): 462-469.
- ↑ a b J. Haviland-Jones, Holly Hale R., P. Wilson & T. R. McGuire (2005). «An Environmental Approach to Positive Emotion: Flowers» (PDF). Evolutionary Psychology 3: pp. pp. 104-132. http://www.epjournal.net/filestore/ep03104132.pdf.
- ↑ Vilarnau, E.G. & Guarro, E. 1958. Jardinería, cultivo de las flores. Editorial Albatros, Buenos Aires, Argentina. 435p.
- ↑ Rosende, T. 1974. Floricultura, tratado del arte de cultivar flores. Ed. Aymi, Buenos Aires. 122 p.
- ↑ Licever, R. 1946. Jardinería. Biblioteca de "La Chacra", Editorial Atlántida, Buenos Aires, 275 p.
- ↑ Anónimo. «Global flower bulb production» (en inglés). Consultado el 5 de mayo de 2009.
- ↑ Neil O. Anderson. 2007. Flower Breeding and Genetics: Issues, Challenges and Opportunities for the 21st Century. Springer, ISBN 1402065698
- ↑ Elkman, P., Friesen, W. V., and O'Sullivan, M. (1988). "Smiles when lying". Journal of Personality and Social Psychology, 54, pp. 414–420.
Enlaces externos
 Wikiquote alberga frases célebres de o sobre Flor. Wikiquote
Wikiquote alberga frases célebres de o sobre Flor. Wikiquote Wikimedia Commons alberga contenido multimedia sobre floresCommons.
Wikimedia Commons alberga contenido multimedia sobre floresCommons. Wikcionario tiene definiciones para flor.Wikcionario
Wikcionario tiene definiciones para flor.Wikcionario
Categorías:- Flores
- Morfología de la flor
- Términos botánicos
- Polinización
-












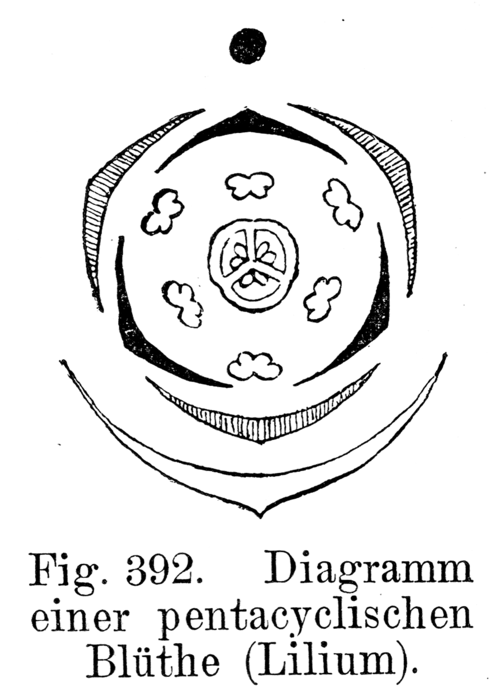






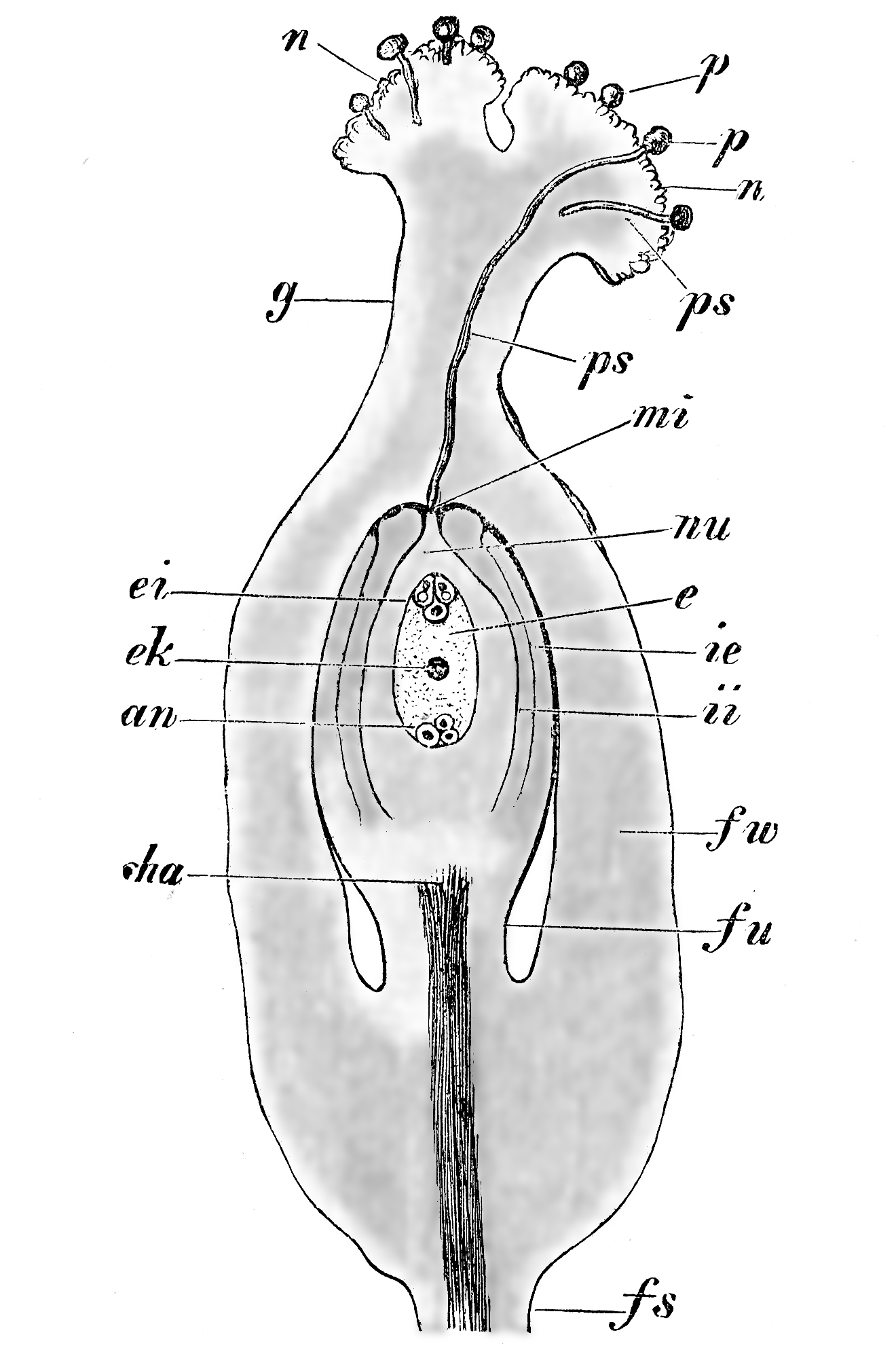






Wikimedia foundation. 2010.