- Orchidaceae
-
 Orquídeas
Orquídeas
Flores de cuatro especies de orquídeas:
Phalaenopsis hieroglyphica (arriba, izquierda)
Ophrys tenthredinifera (arriba, derecha)
Paphiopedilum concolor (abajo, izquierda)
Maxillaria tenuifolia (abajo, derecha).Clasificación científica Reino: Plantae División: Angiospermae Clase: Monocotyledoneae Orden: Asparagales Familia: Orchidaceae
Juss., nom. cons.Diversidad 800 géneros y más de 20.000 especies. Subfamilias Comprende cinco subfamilias:[1]
Las orquídeas u orquidáceas (nombre científico Orchidaceae) son una familia de plantas monocotiledóneas que se distinguen por la complejidad de sus flores y por sus interacciones ecológicas con los agentes polinizadores y con los hongos con los que forman micorrizas.
La familia comprende aproximadamente 25.000 (algunas fuentes informan de 30.000) especies, y quizá otros 60.000 híbridos y variedades producidas por los horticultores, por lo que resulta ser una de las familias con mayor riqueza de especies entre las angiospermas. Pueden ser reconocidas por sus flores de simetría fuertemente bilateral, en las que la pieza media del verticilo interno de tépalos —llamada labelo— está profundamente modificada, y el o los estambres están fusionados al estilo, al menos en la base.[1]
Las orquídeas constituyen un grupo extremadamente diverso de plantas, que pueden tener desde unos pocos milímetros de longitud (ciertas especies de los géneros Bulbophyllum y Platystele) hasta constituir gigantescas agregaciones de varios cientos de kilogramos de peso (algunas especies de Grammatophyllum) o presentar longitudes de hasta 13,4 m, como es el caso de Sobralia altissima, una orquídea recién descrita en 1999 en el Perú.[2] [3] Del mismo modo, las flores de las orquídeas varían en tamaño desde menos de 1 mm y difícilmente visibles a simple vista (Platystele) pasando por las grandes flores de 15 a 20 cm de diámetro en muchas especies de los géneros Paphiopedilum, Phragmipedium y Cattleya hasta los 76 cm de las flores de Phragmipedium caudatum. La fragancia de sus flores no es menos variable, desde el delicado aroma de Cattleya hasta el repulsivo hedor de las flores de ciertas especies de Bulbophyllum .[3]
Se encuentran en la mayor parte del mundo, si bien son especialmente abundantes en los trópicos. No obstante, su capacidad de adaptación les ha permitido conquistar un sinnúmero de nichos ecológicos, desde los más secos y calientes del planeta hasta los más húmedos y fríos ya que, literalmente, se distribuyen desde las regiones polares hasta el ecuador.[3]
La familia ha sido reconocida por los sistemas clásicos de clasificación de plantas, como el sistema de Cronquist,[4] así como por los más modernos, como el sistema de clasificación APG II y el sistema de clasificación APG III.[5] [6]
Contenido
Descripción
Hábito
Véase también: Hábito (planta)
 Cephalanthera longifolia , una orquídea terrestre.
Cephalanthera longifolia , una orquídea terrestre. Archivo:Pseudobulbe+.JPGVariabilidad en la forma de los pseudobulbos de distintas orquídeas: 1.Hoffmannseggella 2.Laelia 3.Ada 4.Cattleya 5.Guarianthe 6.Sobralia 7.Thunia 8.Bulbophyllum 9.Cirrhopetalum 10 a 12.Dendrobium 13. Flickingeria 14. Epidendrum 15.Oncidium, 16.Cymbidium 17.Stanhopea 18.Lycaste 19.Catasetum 20.Gongora 21.Coryanthes.
Archivo:Pseudobulbe+.JPGVariabilidad en la forma de los pseudobulbos de distintas orquídeas: 1.Hoffmannseggella 2.Laelia 3.Ada 4.Cattleya 5.Guarianthe 6.Sobralia 7.Thunia 8.Bulbophyllum 9.Cirrhopetalum 10 a 12.Dendrobium 13. Flickingeria 14. Epidendrum 15.Oncidium, 16.Cymbidium 17.Stanhopea 18.Lycaste 19.Catasetum 20.Gongora 21.Coryanthes. Crecimiento monopodial en Vanilla planifolia.
Crecimiento monopodial en Vanilla planifolia. Crecimiento simpodial en Lycaste xytriophora, obsérvense los pseudobulbos.
Crecimiento simpodial en Lycaste xytriophora, obsérvense los pseudobulbos.Las orquídeas son plantas herbáceas, perennes (raramente anuales), terrestres o epífitas, ocasionalmente trepadoras, algunas veces saprófitas o, raramente, micoheterotróficas. Con respecto a las orquídeas epífitas, se dice que pueden llegar a ser eternas. De hecho, en la naturaleza, su supervivencia está ligada a la vida del árbol que las sostiene. Se conocen plantas recolectadas a mediados del siglo XIX que todavía están creciendo y floreciendo en muchas colecciones.[7]
Los tallos son rizomas o cormos en las especies terrestres. En las especies epífitas, en cambio, los tallos se hallan engrosadas en la base formando pseudobulbos que sirven para almacenar agua y nutrientes y que, por lo general, están recubiertos por las vainas foliares membranosas que se secan con la edad.[8]
Existen dos tipos básicos de crecimiento dentro de la familia: el tipo simpodial, que origina tallos múltiples, y el tipo monopodial, que origina un solo tallo.[9] El tipo simpodial de crecimiento es el más común dentro de la familia. La mayoría de estas orquídeas presentan pseudobulbos que funcionan como reservorios de agua y nutrientes. La planta sostiene los pseudobulbos casi verticalmente y el crecimiento y desarrollo posterior de nuevos tallos se produce horizontalmente, entre los pseudobulbos preexistentes. Cada nuevo pseudobulbo se origina en la base de los anteriores y, con su crecimiento, origina nuevas hojas y raíces. Las hojas originadas en cada pseudobulbo pueden durar muchos años, proveyendo nutrientes para toda la planta, hasta que se tornan marrones y mueren. Aún sin hojas, cada pseudobulbo continúa sosteniendo el crecimiento y suministrando la energía necesaria para el crecimiento del resto de la planta y para la floración. Algunos ejemplos de orquídeas con este tipo de crecimiento son los géneros Cattleya, Dendrobium y Oncidium.[9] Las orquídeas con crecimiento monopodial, a diferencia de las anteriores, presentan un solo tallo principal que crece erecto e indefinidamente desde el centro de la planta.[10] Normalmente, el tallo va creciendo hacia arriba y se originan raíces en los nudos, las cuales crecen hacia abajo. La planta, conforme va creciendo, pierde las hojas inferiores a medida que se forman nuevas hojas en el extremo superior. Algunas especies de orquídeas con este tipo de crecimiento son aquellas pertenecientes a los géneros Ascocentrum, Phalaenopsis y Vanda.[9]
Raíz
Las orquídeas terrestres a veces presentan raíces tuberosas. En las orquídeas epífitas, en cambio, las raíces son aéreas y están muy desarrolladas, cuelgan de los árboles y son verdes y gruesas. Las raíces de las epífitas tienen una doble función, son las estructuras que se encargan de captar los nutrientes que la planta necesita y funcionan, además, como elementos de fijación. Las raíces en este tipo de orquídeas típicamente poseen una epidermis esponjosa, formado por muchas capas de células muertas a la madurez y con paredes celulares engrosadas, llamada velamen. El velamen constituye una vaina esponjosa y blanquecina que rodea por completo a la raíz. Si el tiempo está seco, sus células están llenas de aire; pero cuando llueve se llenan de agua.[7] Según algunos autores el velamen es un tejido que absorbe agua, según otros nunca se ha observado el paso de agua del velamen al córtex de la raíz. Su función principal parece ser la de protección mecánica, además de impedir la excesiva pérdida de agua de la raíz en períodos de deficiencia hídrica. Además, cuando el velamen se llena de agua se vuelve transparente permitiendo a la luz alcanzar el tejido verde de las raíces y, por ende, facilita la fotosíntesis.[11] [12] [13]
 Diferentes tipos de hojas en las orquídeas.
Diferentes tipos de hojas en las orquídeas.Hojas
Del rizoma o de los tallos aéreos nacen las hojas, las cuales son simples y de margen entero, generalmente alternas, espiraladas, dísticas o verticiladas, muchas veces plicadas, basales o a lo largo del tallo, a veces reducidas a vainas o a escamas, usualmente con venación paralela y envainadoras en la base. Pueden presentar pecíolo o ser sésiles y no presentan estípulas. Las especies adaptadas a períodos de sequía tienen hojas carnosas que cumplen la función de reserva de agua en épocas de escasez.[14] [15] [3]
Flor
Ninguna familia de plantas tiene una gama de flores tan variada. Las flores están altamente especializadas en relación a sus polinizadores. Son bien conocidas las variaciones estructurales que facilitan la polinización por una determinada especie de insecto, pájaro o murciélago.
 Flor de Cattleya labiata, obsérvese la disposición de las piezas del perianto en relación al esquema superior.
Flor de Cattleya labiata, obsérvese la disposición de las piezas del perianto en relación al esquema superior.Las flores pueden surgir, dependiendo del género y la especie, de la base de la hoja, del rizoma o de algún entrenudo del pseudobulbo. Son hermafroditas (raramente unisexuales), en general zigomorfas (de simetría bilateral), usualmente resupinadas (es decir, las partes florales giran 180° durante el desarrollo), muchas veces conspicuas y epíginas (es decir, las piezas del perianto se disponen por encima del ovario).
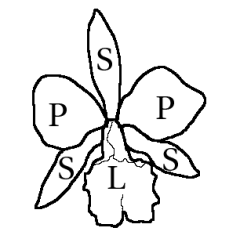
En la gran mayoría de los géneros, las flores están formadas por tres piezas externas llamadas sépalos, dos laterales y uno dorsal, y tres elementos internos llamados pétalos, el inferior modificado en un labio o labelo de tamaño mayor y color más intenso que los demás. Algunos autores interpretan al perianto de las orquídeas como un perigonio, formado por seis tépalos dispuestos en dos verticilos.[16] [17] Las diferentes piezas del perianto pueden estar separadas entre sí o fusionadas en la base.
Los sépalos, o tépalos externos, son usualmente petaloideos (similares a pétalos), imbricados. A veces los dos sépalos laterales se encuentran fusionados en un solo elemento llamado «sinsépalo». Los pétalos, o tépalos internos, están siempre separados, a veces presentan puntos, manchas y colores muy variados. El llamado «labelo» es el pétalo medio, es de tamaño mayor que los dos pétalos laterales y su forma es extremadamente variable. Es la pieza más compleja y, en cierto modo, un órgano característico de las orquídeas. Puede ser lobulado, y entonces se dice que existe un lóbulo central y dos laterales (Orchis, Dactylorhiza). En otras oportunidades, como en Epipactis, se diferencian transversalmente dos partes que se denominan «hipoquilo» la basal y «epiquilo» la distal. Puede tener áreas brillantes, crestas, quillas u otras protuberancias que se suelen denominar como «callo» o «callus». También es frecuente que desarrolle un espolón dirigido hacia atrás o hacia abajo en donde se aloja un nectario. Este espolón puede ser largo y fino (Gymnadenia, Orchis), o como un saco redondeado (Coeloglossum viride). También hay especies en que el espolón no tiene néctar o puede haber nectarios no incluidos en el espolón.
El androceo está usualmente formado por uno o dos estambres (a veces tres), si es uno solo deriva del estambre medio del verticilo externo ancestral y usualmente con dos estaminodios vestigiales derivados de los estambres laterales de un verticilo interno ancestral. En algunas subfamilias, como en Apostasioideae o Cypripedioideae, hay dos o tres estambres fértiles. Cuando son dos, han derivado de los dos estambres laterales del verticilo interno ancestral, y cuando son tres, se han originado de los dos laterales del verticilo interno y del estambre medio del verticilo externo. El androceo se halla fusionado al estilo y al estigma, los cuales se hallan altamente modificados, formando una estructura conocida como «columna», «ginostemo» o «ginostegio». Las tecas de las anteras se disponen en la porción del ginostemo denominada «clinandro» o «androclino». El polen es granular, en tétradas o aglutinado en grupos de dos a ocho masas suaves o duras llamadas polinias. Estas polinias presentan un apéndice filiforme —llamado «caudícula»— que se une con una masa pegajosa —«retináculo» o «viscidium»— sobre el «rostelo», estructura derivada del estigma con forma de lóbulo alargado y que se sitúa sobre la porción receptiva del estigma. El conjunto de polinios, caudículas y retináculos se denomina «polinario», el cual es la unidad de transporte del polen durante la polinización. Las anteras son de dehiscencia longitudinal y su conectivo muchas veces se halla modificado en un «opérculo» que cubre la antera hasta la polinización.
El gineceo está formado por tres carpelos fusionados entre sí, con el ovario ínfero, que puede presentar un lóculo o tres, y numerosos óvulos (hasta millones) de placentación usualmente parietal, pero ocasionalmente de placentación axilar.
Las orquídeas son, en general, productoras de néctar, sustancia que utilizan como recompensa a los polinizadores. Los nectarios son variables en posición y tipo. Por ejemplo, se encuentran en el espolón del labelo, o en los ápices de los sépalos, o en las paredes internas del gineceo. Las especies que no producen néctar son autógamas o apomícticas, es decir, no necesitan de polinizadores para producir semillas.[18]
Inflorescencia
Las orquídeas llevan sus flores de diversos modos. Aún dentro del mismo género, las diferentes especies pueden tener distintos modos de disponer las flores en inflorescencias, las cuales son indeterminadas y, a veces, reducidas a una única flor, terminal o axilar. La mayoría de las orquídeas tienen inflorescencias que llevan dos o más flores, las que usualmente nacen de un eje floral más o menos alargado que comprende un tallo denominado pedúnculo y una porción que lleva las flores, llamada raquis. En la mayoría de las especies las flores se disponen en un racimo erecto y alargado, con las flores arregladas en una espiral laxa alrededor del raquis (como, por ejemplo, en Cymbidium). En esos racimos las flores individuales se enlazan con el eje floral a través de un corto tallito llamado pedicelo. Puede ser que las flores se articulen con el raquis directamente, sin pedicelo, y —en ese caso— la inflorescencia se denomina espiga, como puede observarse en los géneros Peristylus y Neuwiedia. Un grupo de orquídeas pertenecientes al género Bulbophyllum, bastante espectacular por su floración, presenta el raquis tan contraído que todas las flores parecen salir del mismo punto, como en una umbela. Algunas otras orquídeas (Oncidium, Renanthera), finalmente, presentan inflorescencias ramificadas que se denominan panículas.[7] [19]
Fruto y semilla
El fruto es una cápsula loculícida, que se abre mediante tres o seis ranuras longitudinales (a veces una sola); en raras ocasiones, el fruto de las orquídeas es una baya.
Las semillas son diminutas y numerosas. El tegumento es crustoso o membranoso, sin fitomelaninas, con sólo la capa externa persistente y los tejidos internos colapsados. Las semillas son muchas veces membranosas y aladas, los que les permite ser dispersadas por el viento. El embrión es muy pequeño y no se halla acompañado por endosperma, ya que este tejido aborta muy temprano en el desarrollo embrionario.
Etimología e historia
 Ilustración de Orchis ustulata, una orquídea de hábito terrestre. Obsérvese el parecido de sus tubérculos con los testículos de un mamífero. Esa semejanza es a la que hace referencia el nombre "orquídea".
Ilustración de Orchis ustulata, una orquídea de hábito terrestre. Obsérvese el parecido de sus tubérculos con los testículos de un mamífero. Esa semejanza es a la que hace referencia el nombre "orquídea".La palabra orquídea deriva del griego ορχις (orchis = testículo), vocablo que se encontró por primera vez en los manuscritos de la obra De causis plantarum del filósofo griego Teofrasto y que datan aproximadamente del año 375 antes de Cristo.[10] Tal vocablo hace referencia a la forma de los tubérculos de las especies del género Orchis, orquídeas de hábito terrestre cuyos tubérculos dobles parecen testículos,[20] como puede apreciarse en la imagen de la derecha.
Fueron conocidas y apreciadas por los seres humanos desde la Antigüedad. Existen escritos chinos de 1.500 años de antigüedad donde se hace referencia al cultivo de las orquídeas. En la antigua Grecia se le atribuían propiedades curativas y afrodisíacas.[10] Los aztecas utilizaban una orquídea —la vainilla— para enriquecer una bebida espesa hecha a base de cacao, destinada a los nobles y a los guerreros y era conocida con el nombre de xocoatl.[21] [22]
En Europa, el interés por ellas se despertó hacia 1731 cuando floreció la primera orquídea tropical del Nuevo Mundo, Bletia purpurea (sin. Bletia verecunda), en la colección del almirante inglés Charles Wager quien la obtuvo del Jardín botánico de Chelsea.[23] Desde ese momento, se suscitó un interés sin igual por la adquisición y cultivo de orquídeas exóticas, en particular por los miembros de las clases sociales más acomodadas, quienes debían construir un orquideario como una obligación acorde con su estatus. De hecho, cuando una orquídea florecía en tales colecciones, el evento daba lugar a grandes fiestas y la noticia cubría las primeras planas de la prensa. Para satisfacer este consumo de orquídeas raras y exóticas, durante muchos años los recolectores profesionales provenientes en su mayoría de Francia e Inglaterra se dedicaron a saquear sin misericordia los bosques americanos, poniendo a muchas especies en peligro de extinción. A principios del siglo XX, no obstante, la era de la denominada «orquideomanía» llegaba a su fin. El costo para calefaccionar los invernaderos en los que se debían cultivar estas plantas era extremadamente alto y la carestía energética —agudizada por la primera guerra mundial— dificultó el mantenimiento de los orquidarios privados. Con la depresión de 1929, el cultivo de orquídeas a gran escala definitivamente pasó a manos de empresarios comerciales.[24] [25]
Ecología
Distribución y hábitat
Las orquídeas conforman la familia más grande de las plantas con flores, con alrededor de 20.000 especies divididas en unos 800 géneros distribuidos por todo el mundo. Son una familia cosmopolita, que se halla distribuida desde dentro del Círculo polar ártico hasta Tierra del Fuego y las islas al sur de Australia. Se hallan ausentes solamente en los desiertos verdaderos y en los polos. Son más diversas en las regiones tropicales, donde frecuentemente son epífitas.[26] No obstante, la mayoría de las especies se encuentran en los trópicos y subtrópicos, desde el nivel del mar hasta los 5000 msnm, en casi todos los ambientes. En algunos ecosistemas son el elemento dominante, particularmente en hábitats deficientes en nutrientes. Solamente existen dos ambientes en la tierra donde no prosperan estas plantas, los polos y los desiertos de arena. Son más diversas en las regiones tropicales, donde frecuentemente son epífitas.[27] La mayor cantidad de especies se distribuyen en las regiones tropicales, particularmente en áreas montañosas, las cuales representan barreras naturales y aíslan a las diversas poblaciones de plantas, lo que ocasiona la formación de un elevado número de endemismos. Algunas áreas con una marcada predominancia de orquídeas son las islas y el área continental del sudeste asiático y la región montañosa de Colombia y Ecuador. El tercer sitio con un gran número de especies es la masa atlántica brasilera con, aproximadamente, 1.500 especies descritas.[28] Otras áreas importantes son las montañas del sur del Himalaya en la India y China, las montañas de América Central y el sudeste africano, notablemente la isla de Madagascar.
Ecuador es el país que presenta la mayor riqueza de especies, llegando a totalizar 3.549 taxones de orquídeas registrados, seguido por Colombia con 2.723, Nueva Guinea con 2.717 y Brasil con 2.590.[27] Borneo, Sumatra, Madagascar, Venezuela y Costa Rica son también países con un elevado número de especies.[27]
Su capacidad para adaptarse es notable, ya que pueden crecer tanto a nivel del mar como en los páramos elevados. Muchas viven sobre los árboles (epífitas), otras lo hacen sobre las rocas (litófitas), otras sobre la tierra y algunas especies se desarrollan incluso en ambientes subterráneos. A pesar de lo que mucha gente cree, no son parásitas, ya que no se alimentan del árbol donde viven, sino que lo usan como medio de soporte y como vehículo para alcanzar la luz del sol. Algunas sólo miden unos pocos centímetros y otras pueden tener el porte de un árbol. Sus flores pueden ser tan diminutas que resulta imposible observarlas a simple vista, mientras que otras llaman poderosamente la atención.[27]
Biología de la polinización
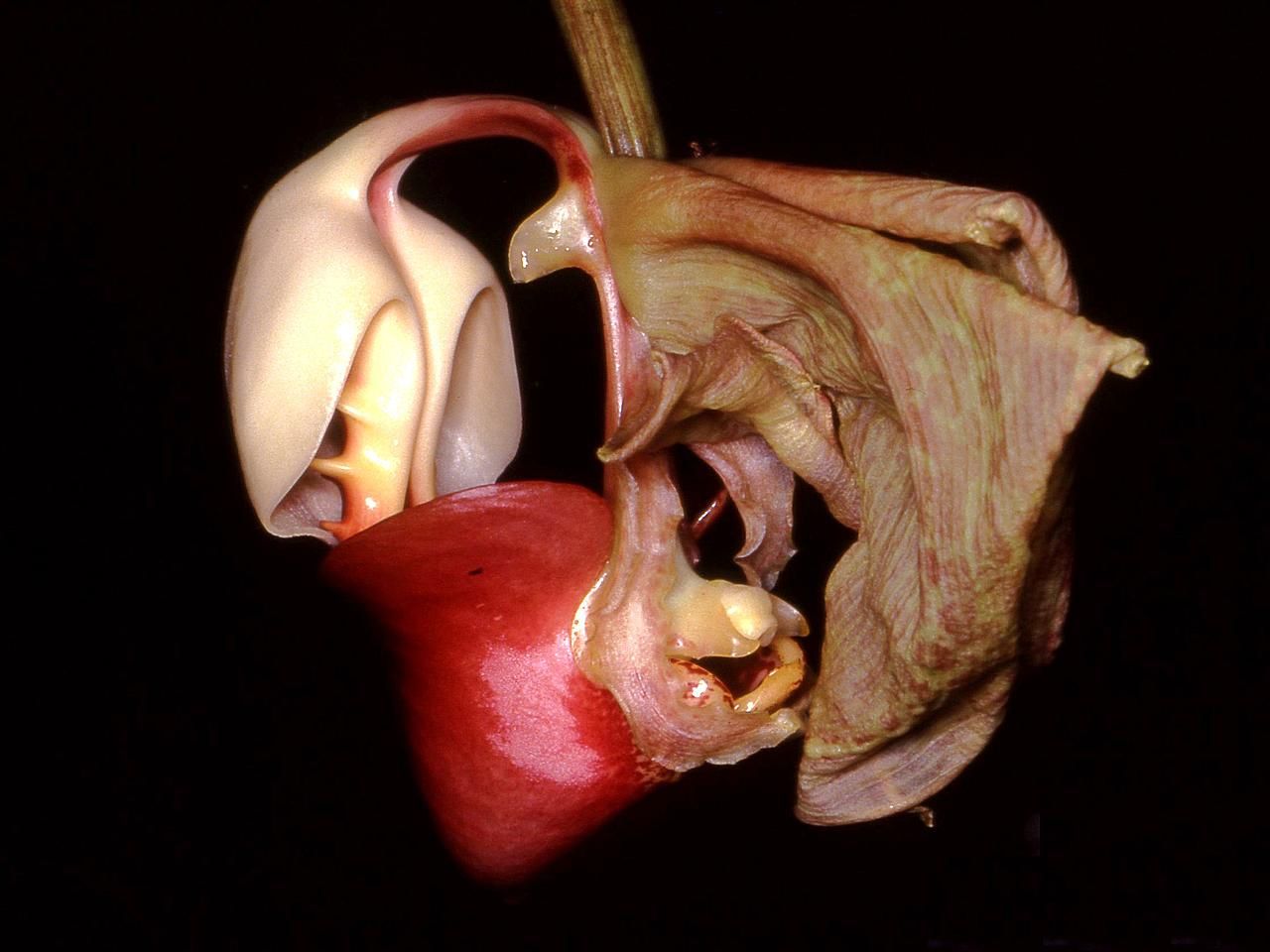
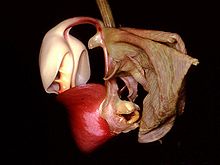 Las formas, colores y fragancias de las orquídeas son el resultado de su coevolución con los animales polinizadores. En la imagen se observa una flor de Coryanthes leucocorys, especie que ha desarrollado uno de los mecanismos más fascinantes de atracción de polinizadores por medio de una bolsa llena de líquido.
Las formas, colores y fragancias de las orquídeas son el resultado de su coevolución con los animales polinizadores. En la imagen se observa una flor de Coryanthes leucocorys, especie que ha desarrollado uno de los mecanismos más fascinantes de atracción de polinizadores por medio de una bolsa llena de líquido. Ilustración de la polinización de Angraecum sesquipedale por una polilla hipotética de larga proboscis. El dibujo fue realizado en 1867 por Alfred Russel Wallace, siguiendo la predicción realizada por Charles Darwin de 1862. En 1903 fue descubierta la polilla en Madagascar y bautizada como Xanthopan morganii praedicta.
Ilustración de la polinización de Angraecum sesquipedale por una polilla hipotética de larga proboscis. El dibujo fue realizado en 1867 por Alfred Russel Wallace, siguiendo la predicción realizada por Charles Darwin de 1862. En 1903 fue descubierta la polilla en Madagascar y bautizada como Xanthopan morganii praedicta.Por lo general, las orquídeas florecen una sola vez al año, siempre por la misma época, la cual está determinada por factores ambientales tales como la disminución o elevación de la temperatura, el incremento de las horas de luz, los cambios de estación y las variaciones en la humedad ambiental. Las flores pueden permanecer abiertas desde un día (el caso de Sobralia) hasta más de tres meses (como en Paphiopedilum y Phalaenopsis). Los híbridos artificiales pueden florecer dos o más veces al año.[29]
El 97% de las especies de orquídeas necesitan de un polinizador para que se lleve a cabo la transferencia de los granos de polen de una planta a los pistilos de otro individuo y, por ende, para que se produzca la fecundación y la formación de las semillas.[29] Se debe tener en cuenta que el polen de las orquídeas se halla agrupado en masas compactas llamadas polinias (singular: polinario), de tal modo que por sí solo, o por acción del viento, el polen no se puede dispersar de una flor a otra por lo que los polinizadores son imprescindibles para asegurar su reproducción sexual.[29] Estos polinizadores son muy variados y, según cuál sea la especie en cuestión, pueden ser moscas, mosquitos, abejas, avispas, coleópteros y aves (especialmente colibríes).[12] [13] [29]
La zoofilia que caracteriza a las orquídeas presupone que los animales polinizadores visiten las flores de manera regular y se detengan en ellas el tiempo suficiente; que las anteras y el estigma sean rozados o tocados con cierta frecuencia y que el primero quede adherido a los visitantes de modo tan perfecto que pueda llegar con la debida seguridad a los estigmas de otras flores. El resultado de la zoofilia depende esencialmente de que los animales puedan reconocer las flores desde una cierta distancia y de que se vean compelidos a visitar durante un cierto tiempo las flores de la misma especie. Las flores zoófilas, entonces, deben poseer productos atractivos (cebos, como el polen y el néctar), medios de reclamo (tales como olores y colores) y, además, polen viscoso o adherente.[12]
Muchas especies de orquídeas recompensan a los polinizadores con alimento (como por ejemplo, néctar, pelos alimenticios o aceites) y otros compuestos, tales como ceras, resinas y fragancias. Estas recompensas, a su vez, refuerzan la conducta de los polinizadores. No obstante, la especialización en un solo tipo de polinizador para asegurar una transferencia más eficiente de polen, determinó una creciente especialización morfológica y estructural en las flores de las orquídeas para garantizar la atracción de una sola especie de insecto.[29] [30] [31]
Por esa razón, las flores de las orquídeas son de formas extremadamente variadas y pueden atraer a una amplia variedad de insectos (abejas, avispas, moscas, mariposas, polillas) así como a pájaros, murciélagos o sapos para la polinización. Algunas atraen visitantes generalistas, pero muchas están bastante especializadas, atrayendo sólo a una o unas pocas especies como polinizadores. Polen, néctar, o fragancias florales pueden ser empleadas como recompensadores de la polinización, mientras que algunas flores (por ejemplo Cypripedium) manipulan a sus polinizadores y no proveen ninguna recompensa, y algunas especies de Ophrys y Cryptostylis mimetizan la forma y el olor de las hembras de abejas, avispas, o moscas, y son polinizadas cuando los machos tratan de aparearse con la flor (fenómeno llamado pseudocopulación).[29]
Generalmente, el labelo funciona como una plataforma de aterrizaje y provee señales visuales o táctiles que orientan al polinizador. La polinia se adjunta al cuerpo del polinizador, y muchas veces es depositada en el estigma (usualmente una depresión en la parte de abajo de la columna) de la siguiente flor visitada. El género Coryanthes tiene un labelo como un bolsillo que se llena con un fluido secretado por la columna, una abeja que cae en este fluido debe viajar a través de un túnel, forzando la deposición del polinario en su cuerpo. La transferencia de polen dentro de la polinia es una aparente adaptación para asegurar la fertilización de muchos del tremendo número de óvulos. En algunas especies la polinización es un evento bastante poco común, y las flores pueden permanecer funcionales y vistosas por muchos días. El marchitamiento del perianto ocurre rápidamente solo después de la fertilización. Angraecum sesquipedale, una orquídea de Madagascar, también es conocida por su biología de la polinización. Esta especie presenta un espolón de 20 a 35 cm de largo, y es polinizada por una mariposa esfíngida, Xanthopan morganii praedicta con una probóscide de esa longitud, un hecho que Charles Darwin había predicho antes del descubrimiento de tal polinizador.[32] [29]
Biología de la germinación
En la mayoría de las especies las pequeñas semillas, que son como polvo, son dispersadas por el viento y requieren nutrientes provistos por un hongo micorrícico para poder germinar. Algunos miembros de Cypripedioideae y Vanilloideae poseen frutos carnosos que fermentan in situ liberando compuestos fragantes (por ejemplo la vainillina) que atraen a pájaros y mamíferos, que actúan como agentes de dispersión.[33]
Estas semillas están formadas por un embrión constituido por pocas células (entre 100 y 200), cubiertas por una testa muy dura. El número de semillas puede variar de 13.000 a 4.000.000 por cápsula. El rango de peso de una semilla de orquídea varía de 0,3 a 14 µg y miden de 0,25 a 1,2 mm de largo y 0,009 a 0,27 mm de ancho. Estas semillas no poseen endosperma y consisten de un pequeño embrión suspendido dentro de una membrana, comúnmente transparente, aunque en ocasiones pigmentada. Las formas de las semillas pueden ser muy variables, existiendo elípticas, filiformes, fusiformes, redondas, globulares o prominentemente aladas. Todas estas características aparentemente maximizan la fecundidad y la efectividad en la dispersión por el viento de las semillas de orquídeas.[34] [33] [35] La germinación de estas semillas tiene lugar por medio de un proceso que es diferente al de la mayor parte de las angiospermas, porque los embriones de orquídeas son, desde un punto de vista anatómico y estructural, extremadamente reducidos y simples. Los embriones de orquídeas germinan y crecen hasta producir una masa de células llamada «protocormo». Estos protocormos, con sus «rizoides» (estructuras en forma de raíces) pueden o no de inmediato comenzar a fotosintetizar. No obstante, para que el protocormo sobreviva, se desarrolle y se transforme en retoño, primero debe establecer una relación simbiótica con un hongo.[36]
El papel que desempeña el hongo es el de suministrar azúcares al protocormo (especialmente a aquellos que no poseen clorofila). El hongo obtiene el azúcar de secciones del substrato (suelo u otro objeto sólido que sirva de organismo huésped a la planta) de la orquídea, es decir, la corteza de un árbol o del suelo. El protocormo, a su vez, provee al hongo con ciertas vitaminas y un hábitat donde vivir. El hongo vive en área del protocormo y del substrato. Con el tiempo, el joven retoño comenzará a producir sus propios nutrientes y la simbiosis no será más necesaria.[37] [38] [39] [40]
Evolución y filogenia
Se han sugerido muchos orígenes posibles para las orquídeas, no obstante, la familia de las hipoxidáceas (o plantas similares a ellas, ya extintas) parecen ser sus progenitores más probables. A pesar de ser la familia de angiospermas más diversa sobre la tierra, las orquídeas no poseen un registro fósil adecuado por lo que muchos aspectos de su historia evolutiva permanecen oscuros.[41] No obstante, en 2007 se ha informado el hallazgo en la República Dominicana del polinario de una orquídea (la que fue denominada Meliorchis caribea) preservado en ámbar y adherido al mesoescutelo de una especie extinta de abeja (Proplebeia dominicana). Ese fósil proviene del Mioceno, aproximadamente 15 a 20 millones de años atrás. Este descubrimiento constituye no sólo el primer fósil de orquídea descubierto sino también el primer antecedente fósil de las interacciones entre las plantas y sus polinizadores. Asimismo, este descubrimiento sumado a análisis cladísticos sobre datos de la morfología, indican que el ancestro más reciente de las orquídeas existentes vivió en el Cretácico superior, entre 76 a 84 millones de años atrás.[42] La monofilia de las orquídeas está sustentada tanto por la morfología como por los análisis de secuencias de ADN (Dressler 1981,[43] 1993,[44] Dressler y Chase 1995,[45] Burns-Balogh y Funk 1986,[46] Judd et ál. 1993,[47] Chase et ál. 2000,[48] Fay et ál. 2000,[49] Freudenstein et ál. 2004[50] ). Asimismo, las relaciones filogenéticas dentro de la familia, motivo de activas investigaciones en las últimas décadas que todavía continúan, han sido dilucidadas mediante análisis cladísticos de la morfología y de secuencias conservadas del ADN (Dressler 1983,[51] 1993,[44] Chase 1986,[52] 1988,[53] Chase y Hills 1992,[54] Chase y Palmer 1992,[55] Cameron et ál. 1999,[56] Kores et ál. 2000,[57] Whitten et ál. 2000,[58] Salazar et ál. 2003,[59] Burns-Balogh y Funk 1986,[46] Cameron 2006,[60] Dressler 1986,[61] 1993,[44] Judd et ál. 1993,[47] Dressler y Chase 1995,[45] Cameron et ál. 1999,[56] Freudenstein y Rasmussen 1999,[62] Cameron y Chase 2000,[63] Freudenstein et ál. 2000,[64] 2004,[50] Molvray et ál. 2000,[65] van den Berg et ál. 2005[66] ).
La filogenia de las orquídeas se halla suficientemente aclarada en la actualidad, lo que permite la reconstrucción de los estados ancestrales y el análisis de la evolución de varios caracteres adaptativos novedosos, los que incluyen los síndromes de polinización altamente especializados, la colonización de hábitats epifíticos y la presencia de metabolismo ácido de las crasuláceas.[67] El metabolismo ácido de las crasuláceas (conocido como CAM, acrónimo inglés para Crassulacean Acid Metabolism) es una ruta fotosintética, muy distribuida taxonómicamente, la cual ha evolucionado en plantas que habitan ambientes con limitaciones de agua y de dióxido de carbono, los cuales incluyen el dosel de los bosques tropicales con disponiblidad hídrica estacional o intermitente.[68] La fotosíntesis C3 es el estado ancestral de la familia y el metabolismo CAM ha evolucionado al menos diez veces de modo independiente, con varias reversiones al estado original. Un gran evento de radiación adaptativa hacia el metabolismo CAM ocurrió dentro de Epidendroideae, el clado con mayor riqueza de especies epífitas de todos los grupos de plantas conocidos, el cual estuvo ligado a la rápida evolución de una gran cantidad de especies durante el período Terciario. De hecho, se ha demostrado que existe una asociación muy estrecha entre el metabolismo CAM y el hábito epifítico, caracteres que habrían evolucionado paralelamente hace unos 65 millones de años, en correspondencia con la progresiva aridificación y reducción de la concentración de dióxido de carbono en la atmósfera durante el Terciario. Aparentemente, la riqueza de especies en las orquídeas está ligada a la colonización de hábitats epifíticos, y la capacidad para hacerlo estuvo relacionada con la adquisición del metabolismo CAM, como un medio de llevar a cabo la fotosíntesis con bajo consumo de agua.[69] No obstante, el cambio del hábito de crecimiento terrestre al hábito epifítico no solo requiere del metabolismo CAM, sino también de la presencia de varios otros atributos adaptativos. Así la presencia de raíces finas, pequeñas y trepadoras es esencial para la adhesión a un sustrato con escasa estabilidad, como las ramas de los árboles.[70] La suculencia y las raíces con velamen se requieren para soportar los períodos prolongados de deficiencia hídrica al estar fijadas a sustratos con escasa o nula capacidad de retención de agua.[71] Finalmente, la densidad poblacional de los organimos epifiticos es en general muy baja,[72] por lo que requieren de sistemas de polinización altamente especializados para lograr la transferencia eficiente del polen entre plantas diferentes.[73] [74] Todas estas adaptaciones morfológicas y fisiológicas se han desarrollado en las orquídeas en múltiples ocasiones, lo que ha determinado la gran riqueza de especies de esta familia y que casi el 70% de las mismas sean epífitas.[75]
Los géneros Apostasía y Neuwiedia (pertenecientes a la subfamilia Apostasioideae) son considerados hermanas del resto de los miembros de la familia (Dressler 1993,[44] Judd et ál. 1993,[47] Dressler y Chase 1995,[45] Neyland y Urbatsch 1996,[76] Cameron et ál. 1999,[56] Freudenstein y Rasmussen 1999[62] ). Estos dos géneros, y especialmente Neuwiedia, han retenido muchos caracteres ancestrales, como por ejemplo la presencia de dos (Apostasia) o tres (Neuwiedia) estambres en sus flores, los que están sólo ligeramente fusionados al estilo y los granos de polen, independientes entre sí y no pegajosos. Las restantes orquídeas, en cambio, presentan polen pegajoso o con los granos de polen fusionados entre sí en el momento en el que son liberados de los estambres. Dentro del grupo de orquídeas con polen pegajoso, las subfamilias Cypripedioideae y Vanilloideae son los clados basales. La primera de ellas (que incluye, por ejemplo, a Cypripedium y Paphiopedilum) es claramente monofilética y sus miembros comparten algunos caracteres exclusivos, como su labelo en forma de saco (con la forma de una zapatilla) y la antera media modificada en un estaminodio con forma de escudo y dos estambres funcionales. Vanilloideae (que contiene por ejemplo a Vanilla, Pogonia y Cleistes) se distingue porque sus flores tienen sólo un estambre funcional y tampoco presentan polinias.
Las restantes subfamilias poseen un solo estambre funcional, los granos de polen se encuentran agrupados formando polinias y el filamento del estambre se encuentra completamente fusionado con el estilo (Dahlgren et ál. 1985,[77] Burns-Balogh y Funk 1986,[46] Judd et ál. 1993[47] ). Este clado, que incluye a todas las especies con polen agrupado en polinias, tiene flores con sólo un estambre funcional (se dicen monandras) y los dos estambres laterales están transformados en esbeltos estaminodios o faltan completamente).
Los análisis de morfología (Freudenstein y Rasmussen 1999[62] ) y algunos análisis moleculares (Cameron et ál. 1999,[56] Molvray et ál. 2000[65] ) indican que las orquídeas monandras son monofiléticas, No obstante, otros análisis moleculares (Cameron 2006,[60] Cameron y Chase 2000,[63] Freudenstein et ál. 2004[50] ), sostienen que la reducción del número de estambres funcionales ha ocurrido en dos oportunidades durante la evolución de la familia. Entre las orquídeas monandras con polinia se reconocen dos grandes subfamilias, Epidendroideae y Orchidoideae (incluyendo Spiranthoideae). Los miembros de Epidendroideae comparten la apomorfía de presentar una antera picuda e incumbente (es decir, la antera se halla curvada sobre el ápice de la columna), mientras que los integrantes de Orchidoideae comparten las apomorfías de un ápice de la antera agudo, tallos suaves y hojas convolutas.[78] [44] El árbol filogenético de las subfamilias es el que sigue (Judd et ál. 2007, modificado de Cameron et ál. 1999,[56] Kocyan et ál. 2004[79] ):
Orchidaceae polen pegajoso polinia Orchidoideae (incluyendo Spiranthoideae)
Taxonomía
Algunos autores, como por ejemplo Dahlgren et ál. (1985), reconocieron tres familias de orquídeas basándose en el número de anteras del androceo: Apostasiaceae, con dos o tres anteras sólo parcialmente fusionadas al gineceo; Cypripediaceae, con dos anteras fusionadas al gineceo y Orchidaceae, la familia más rica en cantidad de especies, las cuales presentan una única antera fusionada al gineceo.[77] No obstante, todas las evidencias moleculares hasta la fecha muestran que Orchidaceae definida o circunscripta de ese modo sería polifilética.[80] Los datos morfológicos y moleculares indican, en cambio, que Orchidaceae debería definirse en forma amplia, por lo que una nueva clasificación de la familia que se corresponde con la filogenia conocida fue publicada por Chase y colaboradores en 2003, en la que se reconocen cinco subfamilias,[80] las que se describen a continuación:
Apostasioideae
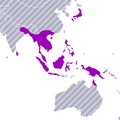 Mapa de distribución de las orquídeas apostasioides.
Mapa de distribución de las orquídeas apostasioides.Las orquídeas apostasioides se consideran el grupo de orquídeas más primitivo. Presentan dos o tres estambres en sus flores, las cuales son «regulares» y se parecen a las del género Hypoxis (de la familia Hypoxidaceae). Las hojas se disponen en forma espiralada en los tallos, son plegadas, resupinadas (salvo en Apostasia). El saco embrionario es bispórico, del tipo Allium. El número cromosómico básico es x= 24. Incluye solo dos géneros (Apostasia y Neuwiedia) y aproximadamente 16 especies.[1] [26] [81]
Sinonimia: Apostasiaceae Lindley, Neuwiediaceae Reveal & Hoogland.
Cypripedioideae
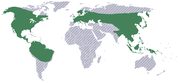 Mapa de distribución de la subfamilia Cypripedioideae.
Mapa de distribución de la subfamilia Cypripedioideae.Este segundo grupo de orquídeas representan un linaje independiente, con categoría taxonómica de subfamilia: las cipripedióideas. También retienen características primitivas, tales como la presencia de dos estambres en las flores. Comprende cinco géneros: Cypripedium, Mexipedium, Paphiopedilum, Phragmipedium y Selenipedium y cerca de 150 especies, las cuales se distribuyen en cinco tribus monotípicas. Están ampliamente distribuidas en Eurasia y a través de América.[1] [26]
Conocidas popularmente como «zapatillas de dama» debido a la abultada forma de zapatilla de su labelo que funciona como atrapa insectos, ya que el insecto es forzado a pasar con la espalda por el estaminodio, con lo que se recolectan o depositan los polinia. En estas orquídeas, dos anteras fértiles se disponen a cada lado de la columna. El estambre central es estéril y está curiosamente modificado como un escudo que impide el acceso directo de los polinizadores desde del frente de la flor a la parte central. Los otros dos estambres están escondidos detrás de este estaminodio. El labelo saculiforme ha evolucionado como una trampa para los polinizadores. Las paredes internas del labelo son muy resbalosas pero una escalera de pelos yace en el interior de la pared dorsal. Este conduce bajo el estigma ventral a una de las dos salidas en la base del labelo a cada lado de la columna.[82] [83] [19]
Sinonimia: Cypripediaceae Lindley.
Vanilloideae
Las orquídeas vanilóideas son un pequeño grupo que incluye a Vanilla, un género de aproximadamente 70 especies de lianas. Comprende 15 géneros y 180 especies que se distribuyen en la franja tropical y subtropical húmeda del globo y en los Estados Unidos de América.[84]
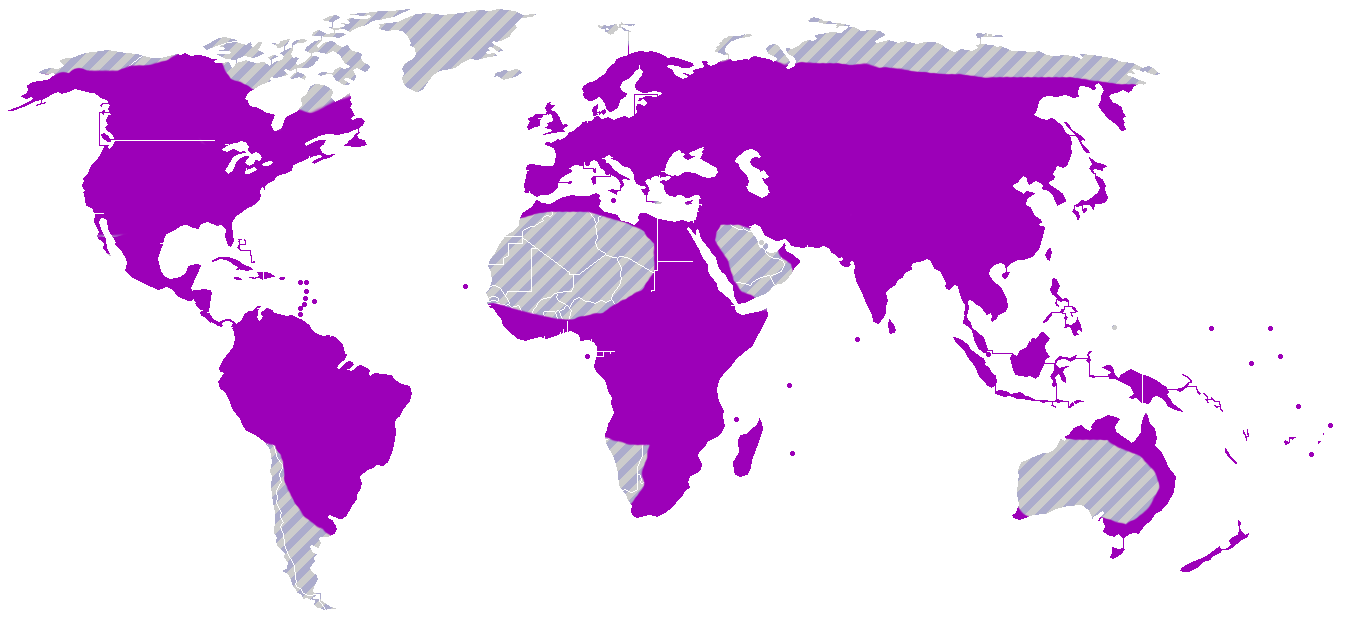
Orchidoideae
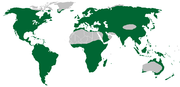 Mapa de distribución de la subfamilia Orchidoideae.
Mapa de distribución de la subfamilia Orchidoideae.Esta subfamilia incluye en su mayoría orquídeas terrestres con tubérculos o rizomas carnosos. El género tipo Orchis y las "orquídeas abeja" (Ophrys, que se denominan así porque su labelo parece el abdomen de una abeja) pertenecen a este grupo. Comprende 208 géneros y 3.630 especies distribuidas en todo el mundo, excepto en los desiertos más secos, en el círculo polar Ártico y en la Antártida. Los miembros representativos de Orchidoideae incluyen a Cynorkis, Diuris, Goodyera, Habenaria, Orchis, Platanthera, Spiranthes, y Zeuxine.[85]
Epidendroideae
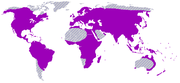 Mapa de distribución de la subfamilia Epidendroideae.
Mapa de distribución de la subfamilia Epidendroideae.Más de 500 géneros y cerca de 20.000 especies distribuidas en las mismas regiones de Orchidoideae, si bien incluyen algunas especies subterráneas del desierto australiano. Epidendroideae contiene numerosas epífitas tropicales, entre los géneros representativos se incluyen Bulbophyllum, Catasetum, Dendrobium, Epidendrum, Encyclia, Maxillaria, Oncidium, Pleurothallis y Vanda. La delimitación de los géneros en este grupo es notoriamente problemática, y los géneros más numerosos no son monofiléticos. La mayoría son epífitas tropicales (normalmente con pseudobulbos), pero algunas son terrestres e incluso unas pocas saprofitas.[86]
Importancia económica
Véase también: Vanilla Dibujo de la vainilla en el Códice Badiano (circa 1580) y descripción de su uso y propiedades, escrito en lengua náhuatl.
Dibujo de la vainilla en el Códice Badiano (circa 1580) y descripción de su uso y propiedades, escrito en lengua náhuatl.Tradicionalmente las orquídeas han sido utilizadas por distintos pueblos con fines ornamentales y medicinales. Los chinos fueron los primeros en cultivarlas desde, aproximadamente, el año 500 a. C. Más tarde, en el siglo V, los griegos las empleaban como plantas medicinales. En América, los aztecas las utilizaban como plantas medicinales, especias, alimenticias y ornamentales.[87] Una de las orquídeas empleadas por este pueblo fue la popular vainilla («tlilxóchitlo» en náhuatl, nombre científico, Vanilla planifolia), usada para aromatizar el chocolate,[88] y llevada a Europa por los conquistadores españoles a principios del siglo XVI y desde ahí a regiones tropicales como Madagascar. Este país se ha convertido en el primer productor del mundo de esta especia, utilizada como saborizante y aromatizante en todo el mundo.[88]
A pesar de la gran diversidad de la familia, pocas orquídeas son cultivadas por otra razón que no sea la belleza de sus flores. Además del ya mencionado cultivo de Vanilla para producir vanillina, algunas pocas especies se utilizan para la producción de aromatizantes del té (Jumellea) o del tabaco (Vanilla). En Turquía se utilizan los tubérculos de Anacamptis morio para la preparación de una bebida típica caliente que se bebe en los días fríos del invierno conocida como salep.[89]
El cultivo de las orquídeas por la belleza de sus flores evolucionó lentamente desde un simple pasatiempo hasta la explotación comercial. Las primeras orquídeas ornamentales llegaron a Europa, procedentes del Nuevo Mundo, en 1731. Sin embargo, no fue sino hasta 1821 cuando se inició su cultivo comercial en invernaderos cerca de Londres. Para 1913 se inauguró en Singapur la compañía «Sun Kee» para producir y comercializar flores cortadas de orquídeas.[90] Actualmente, en Estados Unidos, Inglaterra, Francia, Japón, China, Tailandia, Australia, Hawái y Singapur se ha profundizado el interés por el cultivo y la explotación de orquídeas, con dos objetivos definidos. El primero es el de la producción de flor cortada para abastecer el mercado internacional de floricultura. El segundo objetivo es el de producir y comercializar plantas de diferentes tamaños, en particular las que se hallan cerca de la floración, para abastecer de plantas ornamentales el mercado interno de cada país.[87]
Tailandia es uno de los países más especializados en la producción de flores de orquídeas para abastecer la demanda de las principales ciudades alrededor del mundo,[91] con un monto de exportaciones de 40 millones de dólares para el año 2001.[92]
Entre los géneros de orquídeas más comúnmente cultivados para flor de corte o como plantas ornamentales se destacan Cattleya, Dendrobium, Epidendrum, Paphiopedilum, Phalaenopsis, Vanda, Brassia, Cymbidium, Laelia, Miltonia, Oncidium, Encyclia y Coelogyne. No obstante, la mayor proporción de cultivares actuales de orquídeas (los que se cuentan por más de 100.000) han surgido a través de hibridaciones artificiales entre dos o más especies, muchas veces de distintos géneros.[90]
Multiplicación
El método más simple de multiplicación, a menudo utilizado por los coleccionistas y por los comerciantes de pequeña escala, es la división del tallo. En varias especies de orquídeas, como las pertenecientes al género Dendrobium, el pseudobulbo es largo y articulado, está formado por muchos nudos en los cuales se desarrollan hijuelos. Desde la base de estos hijuelos se desarrollan raíces. Para multiplicar este tipo de orquídeas, entonces, solo se deben cortar los hijuelos enraizados, separarlos de la planta madre y trasplantarlos a otro recipiente. Las especies de orquídeas de mayor importancia comercial, tales como Cattleya, Laelia, Miltonia y Odontoglossum, pueden propagarse por división del rizoma en secciones, las que deben llevar de tres a cuatro pseudobulbos. Los denominados «bulbos traseros», aquellos que ya han perdido el follaje, se usan comúnmente para propagar clones de Cymbidium. Estos bulbos se remueven de la planta y se colocan en otro recipiente con un sustrato adecuado para que formen raíces.[93]
Cuidados
Luz si, pero no sol directo, la temperatura entre 18 y 25 °C., nunca inferior a 16 °C. pues moriría.
El riego, una vez a la semana se coloca la maceta en un plato y se llena el plato de agua, al rato quitar el plato o las raíces se pudrirán.
Se abona una vez al mes, siguiendo las mismas instrucciones que para el riego, es decir, diluyes el abono en el agua del plato y al rato cuando haya absorbido toda la humedad que necesita, retiras el plato.
Para conseguir una mayor y mejor floración, cuando las flores se secan, cortar por encima de la tercera yema y en pocos meses volverá a brotar con mas vigor.
La maceta, como en todas las plantas, se cambia cuando empiezan a asomar por debajo raíces, esa es señal de que ya no le queda tierra, ni sitio para las raíces.
Propagación a gran escala
Debido a la gran cantidad de semillas que se producen en cada fruto y a la posibilidad de cultivar los meristemas in vitro, las orquídeas también pueden ser multiplicadas a gran escala.[94]
Las semillas de las orquídeas son muy pequeñas y las que se hallan en un solo fruto pueden generar miles de nuevas plantas, cada una con características diferente de la otra. No obstante, las semillas contienen escasas reservas y no pueden germinar con sus propios recursos. De hecho, en la naturaleza deben asociarse a un hongo durante la germinación, el cual le provee los nutrientes que requiere para su crecimiento y desarrollo. Por esta razón, la manera más simple —si bien la menos eficiente— de multiplicar orquídeas a través de semillas consiste en esparcir las semillas sobre y alrededor de las raíces de orquídeas cultivadas en maceta y asegurarse de que tengan humedad constante en el sustrato. Las semillas germinan en unas semanas y crecen muy lentamente, de manera que una planta obtenida de este modo florece por primera vez cuando alcanza de cinco a diez años.[94] Este proceso se denomina «germinación simbiótica» y, hasta 1922, era el único método de propagación de orquídeas a través de semillas. En ese año Lewis Knudson de la Universidad Cornell publicó un trabajo en el que describía un método artificial para hacer germinar a las orquídeas sin la participación de un hongo.[95] Este método, llamado de «germinación asimbiótica», hace uso de técnicas de micropropagación para lograr la germinación y establecimiento de las plántulas en un medio de cultivo artificial y bajo condiciones estériles.[96]
La reproducción a través del cultivo in vitro de meristemas, o clonación, es más eficiente y consiste en quitar la punta de la raíz o el extremo de un brote, situarlo en un medio de cultivo adecuado bajo condiciones estériles. Bajo la influencia de los fitohormona los meristemas se convierten en una masa de tejido indiferenciado, capaces de dar lugar a nuevas plántulas. Las plántulas así obtenidas se separan unas de otras y se cultivan en tubos de ensayo independientes. Las plantas son clones perfectos de la planta original, por lo que éste es el método más aplicable a la propagación masiva de una variedad particular o de híbridos estériles.[97]
 Inflorescencia de Campylocentrum grisebachii, una de las orquídeas más pequeñas.
Inflorescencia de Campylocentrum grisebachii, una de las orquídeas más pequeñas.Orquídeas emblemáticas o extremas
- Bulbophyllum minutissimum, originaria de Australia, es la orquídea más pequeña, con solo 3 a 4 mm de altura.[26]
- Cattleya mossiae es la flor nacional de Venezuela.[98]
- Cattleya trianae es la flor nacional de Colombia.[99]
- Guarianthe skinneri es la flor nacional de Costa Rica.[100]
- Lycaste virginalis es la flor nacional de Guatemala.[101]
- Peristeria elata es la flor nacional de Panamá.[102]
- Rhyncholaelia digbyana es la flor nacional de Honduras.[103]
- Sobralia altissima, nativa del Perú, es la orquídea más alta del mundo.[104]
Véase también
- Anexo:Géneros de Orchidaceae
- Listado de Especies europeas de orquídea.
- Orquídeas del Perú
- Centro de Orquídeas de Arona
- Jardín de orquídeas de Kuala Lumpur
- Asociación de Productores y Cultivadores de Orquídeas de la Argentina
- Orquídeas en el Santuario Histórico de Machu Picchu
- Polinización en orquídeas
- La fecundación de las orquídeas
- Salep
- Vanilla
Referencias citadas
- ↑ a b c d Stevens, P. F. (2001). «Angiosperm Phylogeny Website (Versión 9, junio del 2008, y actualizado desde entonces)» (en inglés). Consultado el 12/1/2009.
- ↑ David E. Bennett Jr. y Eric A. Christenson (1999). «Sobralia altissima» (en inglés). Orchids 68 (11): pp. 1112-1113.
- ↑ a b c d Gustavo A. Romero-González, Germán Carnevali Fernández-Concha, Robert L. Dressler, Lawrence K. Magrath & George W. Argus. «Orchidaceae Jussieu» (en inglés). Flora of North America págs. FNA Vol. 26 Page 15, 16, 17, 26, 27, 490, 491, 617. Consultado el 20/3/2009.
- ↑ Cronquist, A. (1981). An integrated system of classification of flowering plants. Nueva York: Columbia University Press.
- ↑ Chase, M. (2003). «An Update of the Angiosperm Phylogeny Group Classification for the orders and families of flowering plants: APG II.» (pdf). Botanical Journal of the Linnean Society (141): pp. 399-436. http://www.blackwell-synergy.com/doi/pdf/10.1046/j.1095-8339.2003.t01-1-00158.x. Consultado el 12/1/2009.
- ↑ The Angiosperm Phylogeny Group III ("APG III", en orden alfabético: Brigitta Bremer, Kåre Bremer, Mark W. Chase, Michael F. Fay, James L. Reveal, Douglas E. Soltis, Pamela S. Soltis y Peter F. Stevens, además colaboraron Arne A. Anderberg, Michael J. Moore, Richard G. Olmstead, Paula J. Rudall, Kenneth J. Sytsma, David C. Tank, Kenneth Wurdack, Jenny Q.-Y. Xiang y Sue Zmarzty) (2009). «An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III.». Botanical Journal of the Linnean Society (161): pp. 105-121.
- ↑ a b c Royal Botanic Garden, Kew. Science & Horticulture. «Orchid structure: the inflorescence.» (en inglés). Major Groups, Families and Genera: Orchidaceae.. Consultado el 7/3/2009.
- ↑ ELY, Francisca, TORRES, Fresia, RADA, Fermín et al. Estudio morfo-anatómico de dos orquídeas de una selva nublada tropical. INCI. [online]. jun. 2007, vol.32, n.º 6, pp. 410-418. Último acceso 5 de julio de 2010. ISSN 0378-1844.
- ↑ a b c Scribd. «Orchid Growth Types: Sympodial or Monopodial» (en inglés). Consultado el 20/3/2009.
- ↑ a b c Freuler, Maria Julia (2008) (en español). Orquídeas. Editorial Albatros. pp. 8. ISBN 9502412184. http://books.google.co.ve/books?id=SjFbL4qd9-MC.
- ↑ Font Quer, P. (1982). Diccionario de Botánica. 8ª reimpresión. Barcelona: Editorial Labor, S. A.. ISBN 84-335-5804-8.
- ↑ a b c Strassburger, E. (1994). Tratado de Botánica. 8ª. edición. Omega, Barcelona, 1088 p.
- ↑ a b Gola, G., Negri, G. y Cappeletti, C. (1965). Tratado de Botánica. 2ª. edición. Editorial Labor S.A., Barcelona, 1110 p.
- ↑ Watson, L.; Dallwitz, M. J.. «Orchidaceae» (en inglés). The families of flowering plants: descriptions, illustrations, identification, and information retrieval. Version: 1st June 2007.. Consultado el 4/11/2007.
- ↑ Renz, J.. «Orchidaceae» (en inglés). Flora of Pakistan . Consultado el 20/3/2009.
- ↑ Simpson, Michael G. (2005). «Orchidaceae». Plant Systematics.. Elsevier Inc.. pp. 171-177. ISBN 0-12-644460-9 ISBN 13 978-0-12-644460-5.
- ↑ Judd, W. S.; C. S. Campbell, E. A. Kellogg, P. F. Stevens, M. J. Donoghue (2007). «Orchidaceae». Plant Systematics: A Phylogenetic Approach, Third edition.. Sunderland, Massachusetts: Sinauer Associates. pp. 273-274. ISBN 978-0-87893-407-2.
- ↑ MRM Neiland, CC Wilcock (1998). «Fruit set, nectar reward, and rarity in the Orchidaceae 1». American Journal of Botany 85 (12): pp. 1657–1657. doi:. http://www.amjbot.org/cgi/content/full/85/12/1657.
- ↑ a b Pridgeon, A.M.; Cribb, P.J.; Chase, M.W. & F. N. Rasmussen (1999): Genera Orchidacearum Vol. 1, Oxford U. Press. ISBN 0-19-850513-2
- ↑ López Trabanco, P. J. 2009. Estudio lingüístico de la fitonimia científica de las orquídeas desde una perspectiva multidisciplinaria. Boletín de lingüística vol. 21, n.º 32, pp. 67-94. ISSN 0798-9709.
- ↑ (en inglés) Ecott T., Travels in search of the ice cream orchid, Vanilla, p.7
- ↑ Bernardino de Sahagún, Historia general de las cosas de Nueva Espana, libro 8 (donde cita las diferentes maneras de tratar el cacao: ...cacao hecho con tlixochitl tierno...) y libro 11.
- ↑ Rigby, R. 2005. Highlights of Orchid History. Consultado el 20 de junio de 2010.
- ↑ Hirtz, A. 2004. Historia de las Orquídeas. Ecuador: terra incognita 3, septiembre-octubre de 2004
- ↑ Berliocchi, L. 2000. The Orchid in Lore and Legend. Timber Press Inc. ISBN 0-88192-616-7
- ↑ a b c d Royal Botanic Garden, Kew. Science & Horticulture. «What are ochids?» (en inglés). Major Groups, Families and Genera: Orchidaceae.. Consultado el 7/3/2009.
- ↑ a b c d Royal Botanic Garden, Kew. Science & Horticulture. «Orchid Fact File» (en inglés). Major Groups, Families and Genera: Orchidaceae.. Consultado el 7/3/2009.
- ↑ Guido Pabst & Fritz Dungs (1975) Orchidaceae Brasilienses vol. 1, Brucke-Verlag Kurt Schmersow, Hildesheim. ISBN 3871050106
- ↑ a b c d e f g van der Cingel, N. 2001. An atlas of orchid pollination: America, Africa, Asia and Australia. CRC Press. ISBN 90-5410-486-4, 9789054104865
- ↑ Dressler, Robert L. 1993, Phylogeny and classification of the orchid family. Cambridge University Press ISBN 0-521-45058-6, 9780521450584
- ↑ Proctor, Michael; Yeo, Peter, Lack, Andrew. 1996. The natural history of pollination. Portland, Oregon. Timber Prees. 479 p.
- ↑ On the various contrivances by which British and foreign orchids are fertilised by insects, and on the good effects of intercrossing. Londres: John Murray. 1862. http://darwin-online.org.uk/content/frameset?viewtype=text&itemID=F800&pageseq=1. Consultado el 1/4/2009.
- ↑ a b Arditti, J., D. Michaud, & P. Healey. 1979. Morphometry of Orchid Seeds. I. Paphiopedilum and Native California and Related Species of Cypripedium. American Journal of Botany 66: 1128-1137.
- ↑ Arditti, J. 1967. Factors affecting the germination of orchid seeds. Botanical Review. 33: 1-97.
- ↑ Arditti, J., D.Michaud, & P. Healey. 1980. Morphometry of orchid seeds. II. Native California and related species of Calypso, Cephalanthera, Corallorhiza, and Epipactis. American Journal of Botany 67: 347-360.
- ↑ Arditti, J., Karim, A., Arim, A. (2000). Numerical and physical properties of orchid seeds and their biological implications. New Phytol. 145, 367-421
- ↑ McKendrick, S., J. Leake, D. Taylor & D. Read. 2000. Symbiotic germination and development of myco-heterotrophic plants in nature: ontogeny of Corallorhiza trifida and characterization of its mycorrhizal fungi. New Phytologist 145:523-537.
- ↑ McKendrick, S., J. Leake, D. Taylor & D. Read. 2002. Symbiotic germination and development of the myco-heterotrophic orchid Neottia nidus-avis in nature and its requirement for locally distributed Sebacina spp. New Phytologist. 154(1): 233-247.
- ↑ Arditti J, Ernst R, Yam TW, Glabe C. 1990. The contribution of orchid mycorrhizal fungi to seed germination: a speculative review. Lindleyana 5: 249-255.
- ↑ Arditti J, Clements MA, Fast G, Hadley G, Nishimura G, Ernst R. (1982). Orchid seed germination and seedling culture. In: Arditti J, ed. Orchid Biology Reviews and Perspectives, vol I. Ithaca, NY, USA: Cornell University Press, 243-370.
- ↑ R Schmid, MJ Schmid (1977). «Fossil history of the Orchidaceae». Orchid Biology: Reviews and Perspectives (USA). http://www.fao.org/agris/search/display.do?f=./1978/v404/US7858767.xml;US7858767.
- ↑ Ramírez. S.R., Barbara Gravendeel, Rodrigo B. Singer, Charles R. Marshall, Naomi E. Pierce (2007). Dating the origin of the Orchidaceae from a fossil orchid with its pollinator. Nature 448, 1042-1045
- ↑ Dressler, R. L. (1981). The orchids: Natural history and classification.. Cambridge, MA.: Harvard University Press.
- ↑ a b c d e Dressler, R. L. (1993). Phylogeny and classification of the orchid family. Portland. OR.: Dioscorides Press.
- ↑ a b c Dressler, R. L.; Chase, M. W. (1995). «Whence the orchids?». En Rudall, P. J., Cribb, P. J., Cutler, D. F., y Humphries, C. J.. Monocotyledons: Systematics and evolution. (Royal Botanic Gardens edición). Kew. pp. 217-226.
- ↑ a b c Burns-Balogh, P.; Funk, V. A. (1986). «A phylogenetic analysis of the Orchidaceae.». Smithsonian Contr. Bot. (61): pp. 1-79.
- ↑ a b c d Judd, W. S.; Stern, W. L., y Cheadle, V. I. (1993). «Phylogenetic position of Apostasia and Neuwiedia (Orchidaceae).». Bot. J. Linn. Soc. (113): pp. 87-94.
- ↑ Chase, M. W.; Soltis, D. E., Soltis, P. S., Rudall, P. J., Fay, M. F., Hahn, W. H., Sullivan, S., Joseph, J., Molvray, M., Kores, P. J., Givnish, T. J., Sytsma, K. J., y Pires, J. C. (2000). «Higher-level systematics of the monocotyledons: An assessment of current knowledge and a new classification.». En Wilson, K. L. y Morrison, D. A.. Monocots: Systematics and evolution. (CSIRO Publ. edición). Collingwood, Australia. pp. 3-16.
- ↑ Fay, M. F. (2000). «Phylogenetic studies of Asparagales based on four plastid DNA regions.». En K. L. Wilson y D. A. Morrison. Monocots: Systematics and evolution. (Royal Botanic Gardens edición). Kollingwood, Australia: CSIRO. pp. 360-371.
- ↑ a b c Freudenstein, J. V.; van der Berg, C., Goldman, D. H., Kores, P. J., Molvray, M., y Chase, M. W. (2004). «An expanded plastid DNA phylogeny of Orchidaceae and analysis of Jacknife branch support strategy.». Amer. J. Bot. (91): pp. 149-157.
- ↑ Dressler, R. L. (1983). «Classification of the Orchidaceae and their probable origin.». Telopea 2: pp. 413-424.
- ↑ Chase, M. W. (1986). A reappraisal of the oncidioid orchids.. 11. pp. 477-491.
- ↑ Chase, M. W. (1988). «Obligate twig epiphytes: a distinct subset of Neotropical orchidaceous epiphytes.». Selbyana: pp. 24-30.
- ↑ Chase, M. W.; Hills, H. G. (1992). «Orchid phylogeny, flower sexuality, and fragrance seeking.». Bioscience 42: pp. 43-49.
- ↑ Chase, M. W.; Palmer, J. D. (1992). «Floral morphology and chromosome number in subtribe Oncidiinae (Orchidaceae): evolutionary insights from a phylogenetic analysis of chloroplast DNA restriction site variation.». En P. S. Soltis, D. E. Soltis, y J. J. Doyle. Molecular Systematics of Plants. Nueva York: Chapman and Hall. pp. 324-339.
- ↑ a b c d e Cameron, K. M.; M. W. Chase, W. M. Whitten, P. J. Kores, D. C. Jarrell, V. A. Albert, T. Yukawa, H. G. Hills, y D. H. Goldman (1999). «A phylogenetic analysis of the Orchidaceae: evidence from rbcL nucleotide sequences.». American Journal of Botany 86: pp. 208-224.
- ↑ Kores, P. J.; P. H. Weston, M. Molvray, y M. W. Chase (2000). «Phylogenetic relationships within the Diurideae (Orchidaceae): inferences from plastid matK DNA sequences.». En K. L. Wilson, D. A. Morrison. Monocots: Systematics and Evolution. Melbourne: CSIRO. pp. 449-456.
- ↑ Whitten, W. M.; N. H. Williams, y M. W. Chase (2000). «Subtribal and generic relationships of Maxillarieae (Orchidaceae) with emphasis on Stanhopeinae: combined molecular evidence.». American Journal of Botany 87: pp. 1842-1856.
- ↑ Salazar, G. A.; M. W. Chase, M. A. Soto Arenas, y M. H. Ingrouille (2003). «Phylogenetics of Cranichideae with an emphasis on Spiranthinae (Orchidaceae: Orchidoideae): evidence from plastid and nuclear DNA sequences.». American Journal of Botany 90: pp. 777-795.
- ↑ a b Cameron, K. M. (2006). «A comparison and combination of plastid atpB and rbcL gene sequences for inferring phylogenetic relationships within Orchidaceae.». Aliso (22): pp. 447-464.
- ↑ Dressler, R. L. (1986). «Recent advances in orchid phylogeny.». Lindleyana (1): pp. 5-20.
- ↑ a b c Freudenstein, J. V.; Rasmussen, F. N. (1999). «What does morphology tell us about orchid relationships? A cladistic analysis.». Amer. J. Bot. (86): pp. 225-248.
- ↑ a b Cameron, K. M.; Chase, M. W. (2000). «Nuclear 18S rDNA sequences of Orchidaceae confirm the subfamilial status and circumscription of Vanilloideae.». En Wilson, K. L. y Morrison, D. A.. Monocots: Systematics and evolution. (CSIRO Publ. edición). Collingwood, Australia. pp. 457-464.
- ↑ Freudenstein, J. V.; Senyo, D. M., y Chase, M. W. (2000). «Mitochondrial DNA and relationships in the Orchidaceae.». En Wilson, K. L. y Morrison, D. A.. Monocots: Systematics and evolution. (CSIRO Publ. edición). Collingwood, Australia. pp. 421-429.
- ↑ a b Molvray, N.; Kores, P. J., y Chase, M. W. (2000). «Polyphyly of mycoheterotrophic orchids and functional influences on floral and molecular characters.». En Wilson, K. L. y Morrison, D. A.. Monocots: Systematics and evolution. (CSIRO Publ. edición). Collingwood, Australia. pp. 441-448.
- ↑ van der Berg, C.; Goldmand, D. H., Freudenstein, J. V., Pridgeon, A. M., Cameron, K. M., y Chase, M. W. (2005). «An overview of the phylogenetic relationships within Epidendropideae inferred from multiple DNA regions and recircumscription of Epidendreae and Arethuseae (Orchidaceae).». Amer. J. Bot. (92): pp. 613-624.
- ↑ Chase MW, Cameron KM, Barrett RL, Freudenstein JV (2003) DNA data and Orchidaceae systematics: a new phylogenetic classification. In KW Dixon, SP Kell, RL Barrett, PJ Cribb, eds, Orchid Conservation. Natural History Publications, Kota Kinabalu, Borneo, pp 69–89
- ↑ LÜTTGE U. Ecophysiology of Crassulacean Acid Metabolism. Ann Bot. 2004;93:629-652.
- ↑ Katia Silvera, Louis S. Santiago, John C. Cushman, Klaus Winter. 2009. Crassulacean Acid Metabolism and Epiphytism Linked to Adaptive Radiations in the Orchidaceae. Plant Physiol. 149: 1838-1847
- ↑ Nieder, J. 2004 Distribution patterns of epiphytic orchids—present research, past causes and future consequences. In Proc. Eur. Orchid Conf. and Show, March 2003 (ed. J. Hermans & P. Cribb), pp. 241–258. London: The British Orchid Council and the Royal Horticultural Society.
- ↑ Chase, M. W. 1987 Obligate twig epiphytism in the Oncidiinae and other neotropical orchids. Selbyana 10, 24–30.
- ↑ Wolf, J. H. D. & Flamenco, A. 2003 Patterns in species richness and distribution of vascular epiphytes in Chiapas, Mexico. J. Biogeogr. 30, 1689–1707.
- ↑ Barbara Gravendeel, Ann Smithson, Ferry J. W. Slik and Andre Schuiteman. 2004. Epiphytism and pollinator specialization: drivers for orchid diversity? Phil. Trans. R. Soc. Lond. B 359, 1523–1535
- ↑ Benzing, D. H. 1990 Epiphytism: a preliminary overview. In Vascular epiphytes. General biology and related biota (ed. P. S. Ashton, S. P. Hubbell, D. H. Janzen, A. G. Marshall, P. H. Raven & P. B. Tomlinson), pp. 1–42. Cambridge University Press.
- ↑ Benzing, D. H. 1987. Vascular epiphytism: taxonomic participation and adaptive diversity. Ann. Miss. Bot. Gard. 74, 183–204.
- ↑ Neyland, R..; Urbatsch, L. E. (1996). «Phylogeny of subfamily Epidendroideae (Orchidaceae) inferred from ndhF chloroplast gene sequences.». Amer. J. Bot. (83): pp. 1195-1206.
- ↑ a b Dahlgren, R. M.; Clifford, H. T., Yeo, P. F. (1985). The families of the monocotyledons. (Springer-Verlag edición). Berlín.
- ↑ Stern, W. L.; Morris, M. W., Judd, W. S., Pridgeon, A. M., y Dressler, R. L. (1993). «Comparative vegetative anatomy and systematics of Spiranthoideae (Orchidaceae).». Bot. J. Linn. Soc. (113): pp. 161-197.
- ↑ Kocyan, A.; Qiu, Y. -L., Endress, P. K., y Conti, E. (2004). «A phylogenetic analysis of Apostasioideae (Orchidaceae) based on ITS, trnL-F and matK sequences.». Plant Syst. Evol. (247): pp. 203-213.
- ↑ a b Chase, M. W.; K. M. Cameron, R. L. Barrett, y J. V. Freudenstein (2003). «A phylogenetic classification of Orchidaceae». En K. M. Dixon, S. P. Kell, R. L. Barrett, y P. J. Cribb. Orchid Conservation. Kota Kinabalu, Sabah, Malasia: Natural History Publications. pp. 69-89.
- ↑ Stern, W. L., V. Cheadle, and J. Thorsch (1993). «Apostasiads, systematic anatomy, and the origins of Orchidaceae». Botanical Journal of the Linnean Society 111: pp. 411–445. doi:.
- ↑ Cash, C. 1991. The Slipper Orchids, Timber Press ISBN 0-88192-183-1.
- ↑ Cox, A.V., A. M. Pridgeon, V. A. Albert, and M. W. Chase. 1997. Phylogenetics of the slipper orchids (Cypripedioideae: Orchidaceae): nuclear rDNA ITS sequences. Plant Systematics and Evolution 208: 197-223. PDF
- ↑ Pridgeon, Alec M.; Cribb, Phillip J.; Chase, Mark.W; Rasmussen, Finn.N 2003. Genera Orchidacearum: Orchidoideae Vanilloideae. Oxford University Press ISBN 0-19-850711-9
- ↑ Bateman, R. Hollingworsth, P., Preston, J., Yi-Bo, L. Pridgeon, A. & Chase, M. (2003). Molecular phylogenetics and evolution of Orchidinae and selected Habenariinae (Orchidaceae). Botanical Journal of the Linnean Society 142 (1)
- ↑ Cameron, Kenneth, Chase, Mark, Whitten, W., Kores, Paul, Jarrell, David, Albert, Victor, Yukawa, Tomohisa, Hills, Harold, Goldman, Douglas. A phylogenetic analysis of the Orchidaceae: evidence from rbcL nucleotide. Am. J. Bot. 1999 86: 208-224
- ↑ a b Orozco Hernández, R. (1996). Las orquídeas. Correo del Maestro Núm. 7. Consultado el 29/7/2009.
- ↑ a b Ossenbach C. (2005). History of orchids in Central America. Part I : from prehispanic times to the independence of the new republics, in Harvard Papers in Botany, vol. 10, nº 2, pp. 197-202 : The History of Vanilla
- ↑ Hoehne, F. C. (1940). Orchidaceae, introdução. En: Flora Brasílica, Fascículo 1, Volumen 12.1; 1 a 12
- ↑ a b Orchidsasia.What are orchids?. Introduction to orchids. Consultado el 29/7/2009.
- ↑ Sahavacharin, O. (1998). Cut Flower Production in Thailand. Food and Agriculture Organization (FAO) of the United Nations. Regional Office for Asia and the Pacific, Bangkok, Tailandia. Consultado el 29/7/2009.
- ↑ Chitrapan Piluek & Siranut Lamseejan (2002). Melhoramento de Orquídeas por meio de Mutações introduzidas por Raios Gama. Consultado el 29/4/2009.
- ↑ Hartmann, H.T. y D.E. Kester. 1991. Propagación de plantas. Principios y prácticas. C.E.C.S.A. México. ISBN 968-26-0156-8
- ↑ a b Waldemar Silva. 1976. Cultivo de Orquídeas no Brasil. Nobel. ISBN 85-213-0383-1.
- ↑ Knudson, L. (1922). Nonsymbiotic Germination of Orchid Seeds Botanical Gazette, Vol. 73, N.º 1, pp. 1-25.
- ↑ Croezen, P. In vitro Orchid cultivation Orchidmania Organization. Consultado el 19/2/2010.
- ↑ Joseph Arditti, Robert Ernst. 1993. Micropropagation of Orchids. John Wiley & Sons. ISBN 978-0-471-54905-5
- ↑ Sinn, M (2006). Local & Holy: Venezuela’s National Flower. Orchid Digest
- ↑ Díaz Piedrahita, Santiago (2005). La flor de mayo, Cattleya trianae, flor nacional. Biblioteca Virtual del Banco de la República Edición original.
- ↑ Museo Nacional de Costa Rica. Los símbolos patrios costarricences . Consultado el 6/3/2010.
- ↑ Superintendencia de Administración Tributaria. Mi Bella Guatemala. Flor nacional. Consultado el 6/3/2010.
- ↑ República de Panamá. Página Oficial.
- ↑ Nelson, C. Flor nacional de Honduras. Universidad Nacional Autónoma de Honduras. Consultado el 3/7/2010.
- ↑ Rach, Nina. Información sobre Sobralia altissima
Bibliografía
- Judd, W. S.; C. S. Campbell, E. A. Kellogg, P. F. Stevens, M. J. Donoghue (2007). «Orchidaceae». Plant Systematics: A Phylogenetic Approach, Third edition.. Sunderland, Massachusetts: Sinauer Associates. pp. 273-274. ISBN 978-0-87893-407-2.
- Simpson, Michael G. (2005). «Orchidaceae». Plant Systematics.. Elsevier Inc.. pp. 171-177. ISBN 0-12-644460-9 ISBN 13 978-0-12-644460-5.
- Soltis, D. E.; Soltis, P. F., Endress, P. K., y Chase, M. W. (2005). «Asparagales». Phylogeny and evolution of angiosperms.. Sunderland, Massachusetts: Sinauer Associates. pp. 104-109.
- Van der Pijl, L., Dodson, C. (1966). Orchid flowers. Their pollination and evolution. University of Miami Press. ISBN 87024-069-2.
-
El contenido de este artículo incorpora material de una entrada de la Enciclopedia Libre Universal, publicada en español bajo la licencia Creative Commons Compartir-Igual 3.0.
Enlaces externos
 Wikimedia Commons alberga contenido multimedia sobre Orchidaceae. Commons
Wikimedia Commons alberga contenido multimedia sobre Orchidaceae. Commons Wikiespecies tiene un artículo sobre Orchidaceae. Wikispecies
Wikiespecies tiene un artículo sobre Orchidaceae. Wikispecies- Orquídea Cattleya bahiana en Planthogar
- Orquídea Cymbidium en Planthogar
- Orquídea Laeliocattleya en Planthogar
- Orquídea Sophrolaeliocattleya en Planthogar
- Base de datos de orquídeas del Jardín Botánico Lankester
- Base de datos de orquídeas
- Guía de orquídeas de Chile
- Orchidaceae de Brasil
- Nuestras amigas orquídeas.
- Manuales, textos y fotos de orquídeas.
- Orquídeas ibéricas














Wikimedia foundation. 2010.