- Magnoliophyta
-
 Magnoliofitas o Angiospermas
Magnoliofitas o Angiospermas
Flores de PrimulaClasificación científica Reino: Plantae División: Magnoliophyta (Magnoliofitas) o Angiospermae (angiospermas) Clases - Amborellales
- Nymphaeales
- Austrobaileyales
- Chloranthales
- Magnoliidae
- Monocotyledoneae - Monocotiledóneas
- Ceratophyllales
- Eudicotyledoneae
sensu APG III (2009),[1] cuando son especificados órdenes en lugar de clases, se asume que están incluidos en su propia clase monotípica.
una variación: Chloranthales incluido en Magnoliidae sensu APWeb (desde el 2008 hasta la fecha de edición de este artículo al menos, agosto del 2010)[2]
- Magnoliopsida o Dicotiledóneas, parafilético con respecto a las monocotiledóneas
- Liliopsida o Monocotiledóneas
en sistemas de clasificación más antiguos como el de Cronquist (1981,[3] 1988,[4] ) el de Engler,[5] etc.Magnoliophyta es el nombre de un taxón de plantas ubicado en la categoría taxonómica de división, que en el sistema de clasificación de Cronquist (1981,[3] 1988[4] ) coincide en su circunscripción con las que en otros sistemas de clasificación son las llamadas angiospermas (en latín Angiospermae, como en el sistema de clasificación de Engler[5] y el moderno sistema de clasificación APG III del 2009,[1] en inglés angiosperms, como en el sistema de clasificación APG II del 2003[6] ). Las flores de las angiospermas se diferencian de las flores del resto de las espermatofitas en que poseen verticilos o espirales ordenados de sépalos, pétalos, estambres y carpelos, y los carpelos encierran a los óvulos y reciben el polen sobre su superficie estigmática en lugar de directamente sobre el óvulo como en gimnospermas [nota 1] .
En algunos sistemas de clasificación se las llama "magnoliofitas" (nombre científico Magnoliophyta), debido a que se creía hasta hace poco tiempo que las magnolias eran las flores más parecidas a la "angiosperma ancestral" (la primera planta de estas características, de la que habría descendido todo el resto de las angiospermas).
El término "angiospermas" proviene de dos palabras griegas: αγγειον (angíon- vaso, ánfora) y σπέρμα (sperma, semilla); así, este término compuesto significa "semillas envasadas", en referencia a que sus óvulos (y posteriormente sus semillas) están encerrados por la hoja fértil portadora de los óvulos o carpelo. De esta forma, el grano de polen para fecundar al óvulo, debe contactar una superficie del carpelo preparada para ello (el "estigma") en lugar de caer directamente sobre el óvulo como en gimnospermas.
La estructura particular de sus flores no es la única diferencia que poseen con el resto de las espermatofitas, otros caracteres morfológicos distintivos son: la reducción del gametofito femenino a sólo unas pocas células, la doble fecundación (con la formación de un tejido nutritivo característico, triploide, llamado endosperma, al tiempo que se fecunda al óvulo), y un xilema y floema distintivos, más recientes que los del resto de las traqueofitas, y más eficientes en muchos aspectos (aunque el xilema parece haber evolucionado ya dentro del clado, siendo las angiospermas más primitivas similares en sus tubos xilemáticos a las gimnospermas).
Las angiospermas son un grupo de espermatofitas reconocido como monofilético desde hace mucho tiempo debido a sus caracteres morfológicos distintivos, y cuya monofilia fue sostenida posteriormente por los análisis moleculares de ADN.
La diversificación que sufrió este grupo es asombrosa. En el registro fósil aparecen a principios del Cretácico (hace unos 130 millones de años), momento a partir del cual aparecen enormes cantidades de fósiles de especies muy diversas, como si hubieran aparecido con toda su diversidad de golpe, lo que Darwin llamó en su momento un "abominable misterio". Aun hoy cerca del 90% de las plantas terrestres pertenecen a este grupo. Con unas 257.000 especies vivientes (Judd et al. 2002), las angiospermas son las responsables de la mayor parte de la diversidad en espermatofitas, en embriofitas y en viridofitas.
Así como posee muchas especies, este grupo se caracteriza por poseer una enorme diversidad de hábitos, y haber ocupado prácticamente todos los nichos ecológicos posibles. Hay plantas arbustivas y herbáceas, las hay terrestres y acuáticas, se encuentran tanto en los desiertos como en los pantanos, en el nivel del mar como en lo alto de las montañas. Su diversidad de especies es mucho más alta en zonas tropicales y húmedas (alrededor del 60% de las especies son de zonas tropicales y un 75% tiene un crecimiento óptimo en climas tropicales), donde dominan completamente el paisaje, y va disminuyendo su número hacia las latitudes altas, llegando a poseer una representación empobrecida en las floras más frías como la de la tundra (que todavía hoy está dominada por las coníferas).
Los miembros de esta división son la fuente de la mayor parte de los alimentos consumidos por el hombre, así como de muchas materias primas y productos naturales. El grueso de la alimentación mundial procede de sólo quince especies.
Contenido
Ciclo de vida de las angiospermas y morfología de la flor
- Véase también: Morfología floral
 Diagrama esquemático del ciclo de vida de las espermatofitas (Spermatophyta).
Diagrama esquemático del ciclo de vida de las espermatofitas (Spermatophyta).
Referencias:
n : generación haploide,
2n : generación diploide,
m! : mitosis,
M! : meiosis,
F! : fecundaciónLas angiospermas pueden a definirse como traqueofitas con las siguientes características:
- Como en todas las traqueofitas, hay dos generaciones multicelulares alternadas, llamadas gametofito y esporofito, siendo el gametofito taloide, y el esporofito organizado en tejidos y órganos. Como en todas las espermatofitas, la alternancia de generaciones gametofítica y esporofítica se da en forma enmascarada, ya que el gametofito se desarrolla por completo dentro de las estructuras del esporófito.
- Como en todas las traqueofitas, el esporofito es un "cormo" (con sistema vascular, raíz y vástago). Como en todas las eufilofitas, el vástago del esporofito crece gracias a su meristema apical, y se ramifica como un tallo principal con ramas laterales, y además tiene hojas ("eufilos").
- Como todas las espermatofitas, tienen un ciclo de vida heterospórico y el gametofito se desarrolla de forma endospórica (esto quiere decir que el esporofito da esporas femeninas dentro de las cuales se desarrolla el gametofito femenino, y esporas masculinas dentro de las cuales se desarrolla el gametofito masculino), los esporangios nacen siempre en las hojas ("hojas fértiles", los estambres y los carpelos), y las hojas fértiles siempre están en una estructura llamada flor (aquí definida como una rama de crecimiento limitado portadora de hojas fértiles). Como en todas las espermatofitas, el megasporangio (presente en el carpelo) produce una única megaspora que nunca lo abandona (megasporangio + megaspora + gametofito femenino + gameta femenina = óvulo). Como en todas las espermatofitas, el microsporangio (o saco polínico, presente en el estambre) produce en su interior "granos de polen" (microsporas con un único gametofito masculino dentro) que son liberados al exterior a la espera de que los agentes externos los transporten hasta las vecindades del óvulo. Como en todas las espermatofitas, el grano de polen al ser estimulado emite un tubo polínico para fecundar al óvulo.
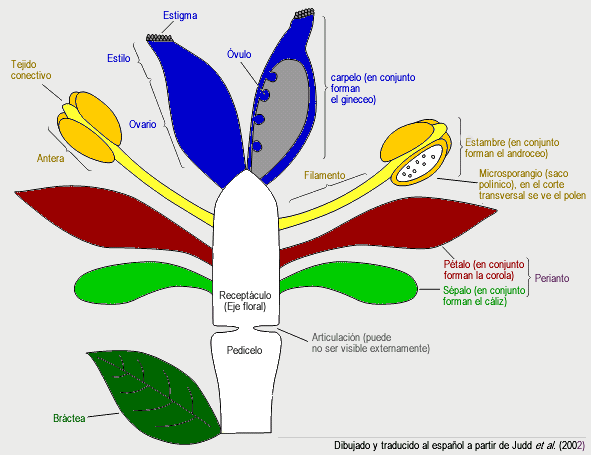
- La flor de angiosperma se caracteriza por poseer un pedicelo que nace en la axila de una bráctea, que remata en un receptáculo portador de los verticilos o espirales de sépalos, pétalos, estambres y carpelos en ese orden. Los sépalos y pétalos son hojas estériles no presentes en el resto de las espermatofitas. Por lo general los sépalos son verdes y fotosintéticos, y tienen una función de protección del capullo cuando la flor todavía está en formación, mientras que los pétalos son vistosos y coloridos, y tienen una función de atracción de los agentes polinizadores, por lo general animales. Por lo general los estambres constan de un filamento y una antera, y la antera posee 4 microsporangios (sacos polínicos) unidos por tejido conectivo. El gametófito masculino está reducido a tres células (dos son espermáticas y la tercera forma el tubo polínico). Por lo general los carpelos son hojas cerradas que mantienen a los óvulos dentro, se diferencian en una superficie preparada para recibir al grano de polen llamada estigma, un cuello (estilo) preparado para hacer crecer al tubo polínico hacia los óvulos, y una cámara que alberga a los óvulos llamada ovario. El gametofito femenino ("saco embionario") en general está reducido a siete células. Los óvulos están unidos a la pared del ovario mediante un tejido a veces llamado "placenta", y la forma de unión (o "tipos de placentación") es útil para la determinación de las familias. No hay que confundir esta placentación con la placenta que une al embrión con el gametofito femenino (característica de todas las embriofitas).
- En las angiospermas se produce el fenómeno de doble fecundación. Esto quiere decir que los dos núcleos espermáticos que libera el grano de polen fecundan uno al óvulo (produciendo el cigoto que dará el futuro esporofito adulto) y el otro a una célula diploide, produciendo un tejido 3n (triploide) de reserva llamado endosperma.
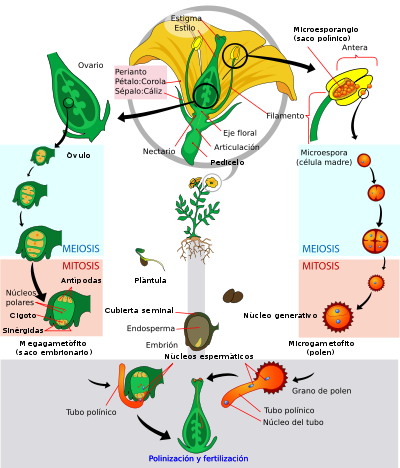 Ciclo de vida.
Ciclo de vida.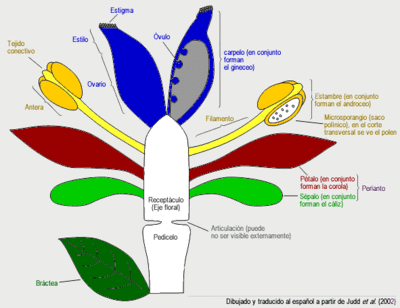 Dibujo esquemático de una flor de angiosperma ancestral con todas sus partes. Las flores de angisopermas se diferencian de las flores del resto de las espermatofitas en que poseen verticilos o espirales ordenados de sépalos, pétalos, estambres y carpelos, los carpelos encierran a los óvulos y reciben el polen sobre su superficie estigmática en lugar de directamente sobre el óvulo como en gimnospermas. Dibujado y traducido al español a partir de Judd et al. (2002).
Dibujo esquemático de una flor de angiosperma ancestral con todas sus partes. Las flores de angisopermas se diferencian de las flores del resto de las espermatofitas en que poseen verticilos o espirales ordenados de sépalos, pétalos, estambres y carpelos, los carpelos encierran a los óvulos y reciben el polen sobre su superficie estigmática en lugar de directamente sobre el óvulo como en gimnospermas. Dibujado y traducido al español a partir de Judd et al. (2002).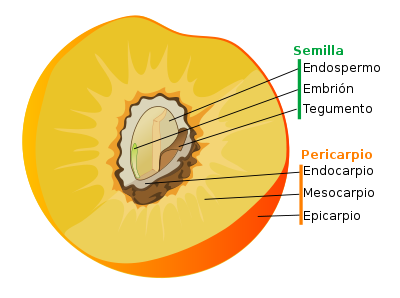 Fruto de angiosperma.
Fruto de angiosperma.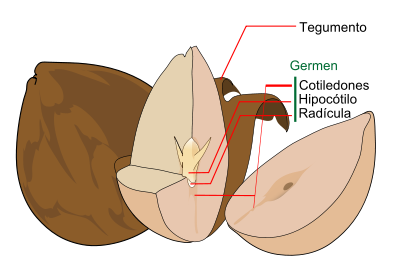 Semilla de angiosperma.
Semilla de angiosperma.Otros caracteres morfológicos
Otras características morfológicas son:
- Muchas angiospermas poseen vasos xilemáticos en lugar de traqueidas, carácter derivado dentro del grupo. En los vasos, el agua puede fluir sin necesidad de atravesar una membrana, lo que los vuelve muy eficientes en el transporte de fluidos dentro del esporofito pero probablemente también más propensos a recibir daño (en especial por embolias de aire) cuando están sujetos a estrés hídrico.
- El floema de las angiospermas difiere del de todas las demás plantas en que los elementos del tubo criboso (que son células vivas pero sin núcleo, encargadas del transporte de azúcares) están acompañadas por una o más células acompañantes, que nacen de la misma célula madre que el elemento.
Sistemática de angiospermas
Taxones aceptados
Tradicionalmente las angiospermas fueron clasificadas en monocotiledóneas y dicotiledóneas (también llamadas Liliopsida y Magnoliopsida). Hoy se sabe que las dicotiledóneas no son monofiléticas, y que este sistema de clasificación no refleja la filogenia de las angiospermas.
Últimos avances en filogenia
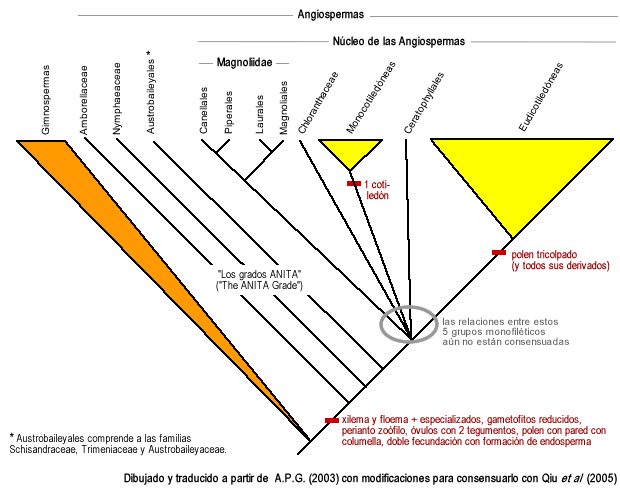
Ya en 1999, Qiu y sus colaboradores[7] publicaron en la prestigiosa revista Nature que, por los estudios moleculares del ADN, las angiospermas más antiguas (en inglés, basal angiosperms), eran, de más antigua a menos antigua: Amborellaceae, Nymphaeaceae, y un orden nuevo de 3 familias al que llamaron Austrobaileyales, que comprende a las familias Schisandraceae, Trimeniaceae y Austrobaileyaceae. Determinaron que uno a uno estos tres clados se fueron desprendiendo de la rama principal de las angiospermas, a estas 5 familias hoy se las conoce con el nombre de los grados ANITA ("ANITA grade" en inglés).
Hoy en día está aceptado que Amborellaceae es la familia más antigua de angiospermas, hermana de todo el resto. Este punto fue rechazado por un trabajo posterior en que se analizaba el genoma completo de los plástidos de varios taxones, pero el trabajo fue posteriormente severamente criticado en sus fallas metodológicas (Soltis y Soltis 2004).[8]
En el 2002 Judd et al. en la segunda edición de su libro Plant Systematics, reconocieron el desprendimiento de estas familias antiguas del clado más reciente de angiospermas, y al clado más reciente lo llamaron Núcleo de las angiospermas (Core Angiosperms en inglés), nombre con el que se conoce al clado hoy en día.
Dentro del núcleo de las angiospermas, varios estudios genéticos, moleculares y también morfológicos hechos hasta el 2006, han confirmado la presencia de 5 clados claramente definidos: las monocotiledóneas, Chloranthaceae, Ceratophyllaceae, un clado que reúne a varias de las familias que Cronquist había llamado Magnoliidae y cuyo nombre se decidió conservar, y un clado que comprende a todo el resto de las angiospermas (aproximadamente el 75% de las especies de angiospermas), que anteriormente se conocía como "el de las dicotiledóneas tricolpadas", y hoy en día se conoce como "Eudicotiledóneas" ("Eudicots" en inglés).
El clado a que se llamó Magnoliidae se divide a su vez en dos clados hermanos, uno que reúne a los Piperales con los Canellales (Canellaceae y Winteraceae), y otro que reúne a los Laurales con los Magnoliales. Estos dos clados, y estos 4 órdenes de los que están compuestos, está consensuado que son monofiléticos.
El Angiosperm Phylogeny Group (A.P.G.) reunió la información molecular y genética que se había acumulado en los últimos años sobre la filogenia de angiospermas, y con ello creó un primer árbol taxonómico en 1998, y un segundo árbol taxonómico en el 2003. Se puede decir que éste fue el primer intento de convertir en taxones a los avances en filogenia. Como novedad se puede remarcar que en el árbol del 2003, se evitó el uso de la nomenclatura taxonómica clásica por arriba de orden, y los taxones superiores a orden fueron llamados con nombres "informales". En general el árbol del 2003 (conocido como APG II) es bien aceptado por la comunidad científica, aunque sólo unos puntos de él están consensuados por todos los trabajos cladísticos hechos hasta la fecha (ver por ejemplo Qiu et al. 2005),[9] y hay que tener en cuenta que todavía no está determinado el árbol filogenético (y por lo tanto tampoco el taxonómico) definitivo.
En resumen, un árbol filogenético consensuado hasta la fecha sería así:
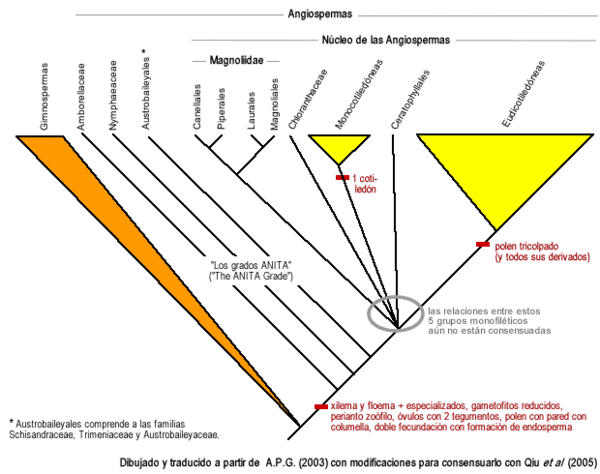
Angiospermas: árbol filogenético consensuado a 2006. Todos los grupos explicitados en el gráfico son monofiléticos, aunque aún falta dilucidar las relaciones entre algunos de ellos. Dibujado y traducido al castellano a partir de APG II 2003, dejando como incógnitas las relaciones que aún no están consensuadas. Taxonomía de angiospermas
- Introducción teórica en Taxonomía
Un sistema de clasificación que incorporó los últimos avances en filogenia a la clasificación, es el Sistema de clasificación APG III publicado en el 2009 (que reemplazó al APG II del 2003 y al anterior APG de 1998). Las correcciones al sistema de clasificación APG III, realizadas con los trabajos que fueron publicados posteriormente al 2009, se pueden encontrar en el APWeb o Angiosperm Phylogeny Website[10] (en inglés), mantenido por uno de los miembros del APG III (P. F. Stevens).
Ecología y Evolución de las angiospermas
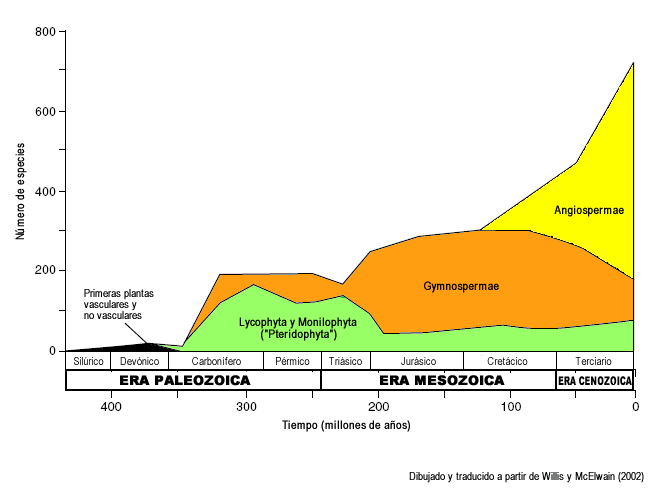
En una carta que le escribió a un amigo, Charles Darwin se refirió a aparición repentina de las angiospermas como un "abominable misterio". En los registros fósiles más antiguos, las primeras plantas vasculares datan de hace 400 millones de años. Luego en el Devónico y en el Carbonífero predominaron las lycophytas, sphenophytas y progimnospermas, que fueron dominantes hasta hace unos 300 millones de años. Las primeras plantas con semilla aparecieron en el Devónico tardío, y eso llevó a la típica flora del Mesozoico dominada por gimnospermas. Los fósiles más antiguos de angiospermas encontrados hasta la fecha datan aproximadamente de hace 140 millones de años (principios del Cretácico), y el número de especies encontrado aumenta de forma llamativa con el tiempo geológico, en comparación a otras plantas no angiospermas. Se diversificaron rápidamente desde hace unos 100 millones de años, llegando a dominar los biomas del globo salvo regiones polares hace unos 65-75 millones de años, dominancia que se mantiene hasta la fecha. Para esa época ya existía la mayor parte de las familias de angiospermas que conocemos hoy en día, e incluso algunos de los géneros actuales.
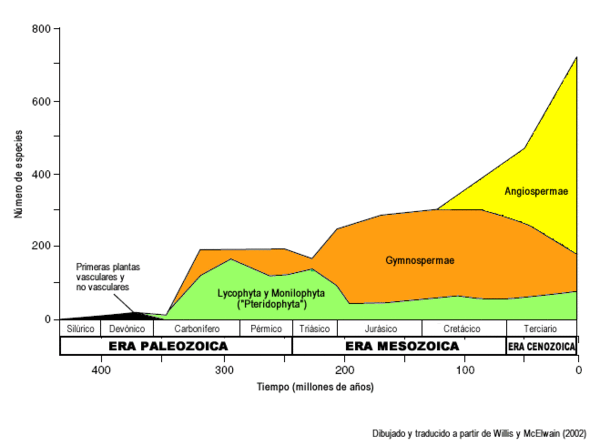
Registro fósil de aparición de los diferentes clados de plantas vasculares. Nótese la intensa radiación de las angiospermas en comparación con los helechos y gimnospermas. Dibujado y traducido a partir de Willis y McElwain (2002). Hasta la fecha de edición de este artículo, se especula que las primeras angiospermas eran pequeñas plantas adaptadas a vivir en la sombra, en lugares perturbados y probablemente también húmedos (Feild, Arens y Dawson 2003),[11] hipótesis llamada en inglés "the dark and disturbed hypotesis". Las evidencias provienen del análisis de las "angiospermas basales" actuales. Con respecto a la evidencia en el registro fósil, Willis y McElwain señalan que los frutos fósiles más antiguos encontrados (pertenecientes a partes de hojas, flores, frutos y polen de los clados ceratophyllales, juglandales y ranunculídeos, entre otros) poseen semillas lo suficientemente pequeñas como para suponer que las primeras angiospermas se dispersaban y colonizaban los ambientes a través de lo que en ecología se conoce como "estrategia r", la estrategia de los primeros colonizadores de los ambientes perturbados (Willis y McElwain 2002). Hay que tener en cuenta que para hacer este análisis, Feild, Arens y Dawson asumieron a Chloranthaceae como una familia de angiospermas basales, y la incluyeron en su análisis, sin embargo la ubicación de Chloranthaceae en el árbol filogenético aún no recibe consenso (Qiu et al. 2005).[9]
Lugar de origen de las angiospermas
Aparentemente, según el registro fósil las angiospermas se originaron a latitudes bajas (cerca del Ecuador), en las que ya estaban bien distribuidas hace unos 130 millones de años, y se volvieron florísticamente prominentes hace 120 millones de años. Luego, hace unos 100 millones de años, se dispersaron hacia latitudes más altas. En el registro fósil de hace 70 millones de años, el 60-80% del polen encontrado en bajas latitudes es de angiospermas, mientras que sólo el 30-50% del polen encontrado en altas latitudes es de angiospermas, correspondiendo el resto del polen encontrado a gimnospermas y pteridofitas. Aparentemente esta reticencia de las angiospermas para dominar la flora en las latitudes más altas se debió, en parte, a las limitaciones estacionales de luz y humedad que hay a altas latitudes, limitaciones que deben haber ofrecido pocas oportunidades a las especies colonizadoras de angiospermas para reemplazar a la antigua vegetación, bien establecida y adaptada, de gimnospermas y pteridofitas (Willis y McElwain, 2002). Esta disminución de la diversidad con el aumento de la latitud, probablemente haya tenido más que ver con restricciones en la biología de las angiospermas existentes en ese momento, que con bajos índices de dispersión (Willis y McElwain, 2002). De hecho, aún hoy la diversidad de especies de angiospermas es más baja que la de gimnospermas en biomas boreales como el de la tundra. Si bien las primitivas angiospermas no parecen haber sido especies invasivas en las altas latitudes, ese escenario parece haber cambiado más tarde, con la aparición de angiospermas más recientes para las cuales las altas latitudes parecen haber ofrecido un escenario óptimo para dominar el ambiente y especiarse posteriormente. Por ejemplo, el sur de Gondwana parece haber sido el origen del género Nothofagus, que apareció hace unos 70 millones de años y aún hoy en día es un componente importante de la flora del Hemisferio Sur.
El enigma de la tardía aparición de las angiospermas: el "abominable misterio" de Darwin
Con respecto a la tardía aparición de las angiospermas en el registro fósil (aproximadamente 300 millones de años después que la aparición de las primeras plantas vasculares), y su explosión de especies después de eso (lo que Darwin llamó "un abominable misterio" en una carta enviada a un amigo), las hipótesis que se manejan actualmente son:
1.- Sesgos en el registro fósil
Significaría que las angiospermas se originaron mucho antes que el primer registro fósil encontrado, pero las primeras angiospermas aparentemente son más difíciles de encontrar que las gimnospermas y pteridofitas de la misma época, debido principalmente a que vivían en un ambiente con un bajo potencial de preservación de los fósiles. Los análisis moleculares que se han hecho (los llamados "relojes moleculares" o "molecular clock" en inglés) apoyan en parte esta hipótesis, pero son duramente cuestionados debido a falencias en su metodología y en las hipótesis en que se basan, por lo que los resultados son observados con escepticismo por la comunidad científica, y aún son tema de debate.
2.- Evolución como respuesta al cambio ambiental
Según esta hipótesis las angiospermas aparecieron tardíamente, pero su evolución fue rápidamente desencadenada por un cambio profundo en las condiciones ambientales, tanto fisicoquímicas como bióticas (co-evolución con grupos animales por ejemplo), que en muy poco tiempo las pusieron en gran ventaja adaptativa con respecto a las gimnospermas y pteridofitas de la época. Las hipótesis que sostienen esto son 3: la que supone que el cambio ambiental fisicoquímico global desencadenó la especiación, la que supone una relación con los dinosaurios, y la que supone una relación con los insectos. Son las siguientes:
2.1.- La hipótesis del cambio ambiental fisicoquímico
Las evidencias señalan que un buen número de cambios en la composición florística del Cretácico (hace 140-80 millones de años) coinciden con una época de cambios ambientales globales, que incluye una actividad tectónica elevada, una disminución del oxígeno en los océanos, y una expansión del suelo oceánico con una definitiva ruptura de la Pangea con la consecuente formación de los continentes de África y Sudamérica, y volviendo distinguibles los platos de la India, Australia y la Antártida. También hubo cambios importantes en la composición de la atmósfera debido a la intensa actividad volcánica (principalmente aumento de la concentración de dióxido de carbono), probablemente un subsiguiente aumento de la temperatura global, un aumento de unos 100 metros en el nivel del mar, y la desaparición de los hielos polares.
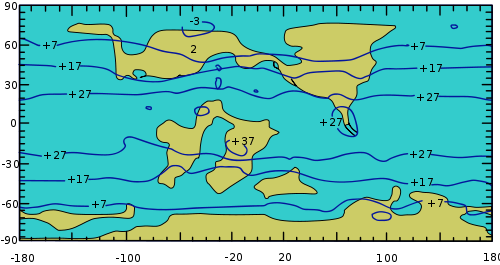 Superficie terrestre durante el Cretácico medio (hace unos 100 millones de años). Nótense las isotermas, que predicen que la temperatura global promedio era unos 4,8 °C más alta que ahora, y la ruptura de la Pangea: hacia el norte se observan Norteamérica y Europa, hacia el sur la Antártida, Australia y la India (aquí amplificados por la deformación del mapamundi), y en el centro los continentes de África y Sudamérica.
Superficie terrestre durante el Cretácico medio (hace unos 100 millones de años). Nótense las isotermas, que predicen que la temperatura global promedio era unos 4,8 °C más alta que ahora, y la ruptura de la Pangea: hacia el norte se observan Norteamérica y Europa, hacia el sur la Antártida, Australia y la India (aquí amplificados por la deformación del mapamundi), y en el centro los continentes de África y Sudamérica.Las evidencias con respecto a la composición florística señalan que en esa misma época hubo un número de innovaciones clave en las angiospermas que las hicieron más resistentes a la sequía y por lo tanto mejor adaptadas a las condiciones ambientales nuevas, entre las que se incluyen hojas reducidas en tamaño, una capa externa de la semilla que las protegía de la sequedad, la aparición de haces vasculares más eficientes en la conducción de agua ("vasos xilemáticos"), y el hábito deciduo (aunque esta última característica también era observada en varias gimnospermas). Hasta el día de hoy no hay evidencia de que esos cambios ambientales globales (conocidos como el "episodio superpluma" o "superplume episode" en inglés) hayan desencadenado la diversificación de las angiospermas, sólo hay evidencia de que ocurrieron simultáneamente.
2.2.- La hipótesis de la coevolución entre las angiospermas y los dinosaurios
Esta hipótesis sostiene que los hábitos alimenticios de los dinosaurios pueden haber promovido la aparición de las angiospermas. Las evidencias señalan que en la época de la aparición de las angiospermas los dinosaurios eran principalmente saurópodos y estegosaurios, quienes tenían una amplia dieta de tejidos de coníferas, y deben haber representado una presión de herbivorismo importante en las copas de las coníferas adultas, presión que aseguraba el desarrollo de las gimnospermas juveniles. Hace unos 144 millones de años cambió la composición de la comunidad de herbívoros, apareciendo nuevos grupos de dinosaurios ("ornithischianos", masticadores eficientes) que probablemente incrementaron la mortalidad entre los juveniles de gimnospermas, creando claros en la cobertura de plantas y nuevos ambientes altamente perturbados. Las evidencias desde el punto de vista de las angiospermas muestran que las angiospermas primitivas parecen haber tenido un rápido ciclo de vida y una alta habilidad de colonización de hábitats perturbados. Sin embargo hoy en día esta hipótesis es rechazada por la comunidad científica, principalmente porque en un análisis del registro fósil, la aparición de los diferentes tipos de dinosaurios herbívoros no parece estar correlacionada con la diversificación de las angiospermas, ni en el tiempo ni en el espacio.
2.3.- La hipótesis de la coevolución entre las angiospermas y los insectos
La hipótesis señala que las flores del Cretácico temprano tenían caracteres que se vinculan a la polinización zoófila, como estambres con anteras cortas y poca producción de polen, y granos de polen cubiertos con un material "pollenkitt-like" (que los adhiere al cuerpo de los insectos que los visitan). También señalan que el polen encontrado era de mayor tamaño que el que es dispersado por el viento en forma efectiva. Además, aparentemente las primeras angiospermas ya presentaban signos de ser autoincompatibles, lo cual no hubiera permitido la autofecundación. Todas estas características sugieren que ya las primeras angiospermas eran polinizadas por insectos. Las ventajas adaptativas de la polinización por insectos son: la posibilidad de intercambio genético entre individuos muy lejanos y en poblaciones muy pequeñas. Esto la hace una hipótesis muy atractiva, pero los registros fósiles son ambiguos. Aparentemente los aparatos bucales de los insectos y las especies de angiospermas se diversificaron aproximadamente en la misma época, pero los análisis no muestran una relación muy significativa: los dípteros (moscas y mosquitos), los coleópteros (escarabajos) y los primeros himenópteros (avispas) aparecieron antes que las primeras angiospermas, algunos himenópteros y los lepidópteros (mariposas y polillas) aparecieron más o menos en la misma época que las angiospermas, y otras especies de himenópteros, incluyendo Anthophila (abejas), aparecieron hace 100 millones de años, por lo que son más recientes que las angiospermas. Por otro lado, hay cada vez más evidencias de que los "polinizadores avanzados" (abejas, avispas y moscas) deben haber jugado un rol importante en la evolución y radiación de ciertos grupos de angiospermas, pero la evolución de las angiospermas como grupo no puede ser explicada exclusivamente por su coevolución con los insectos.
2.4.- Altas tasas intrínsecas de especiación
Una explicación alternativa propone que las angiospermas simplemente tienen tasas altas de mutación, lo que genera una cantidad importante de plantas adaptadas a diferentes condiciones ambientales, lo que promueve su especiación por selección natural.
Las angiospermas del Cretácico tardío como reflejo de la flora actual
Una parte significativa de la flora actual tanto del Hemisferio Norte como del Hemisferio Sur, apareció por primera vez a fines del Cretácico (hace 100-65 millones de años), incluyendo a familias como Ulmaceae (hay registros fósiles del género Ulmus), Betulaceae, Juglandaceae, Fagaceae (especialmente Nothofagus) y Gunneraceae. La mayor parte de los árboles aparecidos en esta época, hoy en día tienen una distribución que es principalmente tropical o subtropical. Es interesante notar que la mayor parte de las angiospermas actuales son básicamente tropicales en sus requerimientos, con aproximadamente la mitad de todas las familias de angiospermas confinadas a regiones tropicales, y unos tres cuartos de las familias llegando a óptimos de crecimiento en ambientes tropicales. Por lo tanto, aunque algunas de ellas sean consideradas como de hábitats templados del hemisferio norte o del hemisferio sur, probablemente todavía tengan muchas de las características que les permitieron sobrevivir en el ambiente en que se originaron.
Véase también
- Clasificación de los organismos vegetales para una introducción a la taxonomía de las plantas.
- Sistema de clasificación APG III para la clasificación actual de las angiospermas.
Notas
- ↑ En algunos textos se considera que sólo las angiospermas son plantas con flores, ya que sólo consideran flores a las flores de angiospermas (que ancestralmente poseen verticilos o espirales ordenados de sépalos, pétalos, estambres y carpelos, y poseen los óvulos encerrados en los carpelos de forma que se pueda acceder a ellos a través del estigma). Aquí, siguiendo la denominación de Pio Font Quer en su Diccionario de Botánica, consideramos que una "flor" es toda rama de crecimiento limitado que posea hojas fértiles, por lo que todas las espermatofitas (tanto angiospermas como gimnospermas) poseen flores, como aquí definido.
Referencias citadas
- ↑ a b The Angiosperm Phylogeny Group III ("APG III", en orden alfabético: Brigitta Bremer, Kåre Bremer, Mark W. Chase, Michael F. Fay, James L. Reveal, Douglas E. Soltis, Pamela S. Soltis y Peter F. Stevens, además colaboraron Arne A. Anderberg, Michael J. Moore, Richard G. Olmstead, Paula J. Rudall, Kenneth J. Sytsma, David C. Tank, Kenneth Wurdack, Jenny Q.-Y. Xiang y Sue Zmarzty) (2009). «An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III.» (pdf). Botanical Journal of the Linnean Society (161): pp. 105-121. http://www3.interscience.wiley.com/journal/122630309/abstract.
- ↑ P. F. Stevens, 2001 en adelante, Angiosperm Phylogeny Website. Versión 8, junio del 2007, y actualizado desde entonces. http://www.mobot.org/MOBOT/research/APweb/
- ↑ a b Cronquist, A. (1981). An integrated system of classification of flowering plants.. Nueva York: Columbia University Press.
- ↑ a b Cronquist, A. (1981). The evolution and classification of flowering plants.. Nueva York: Columbia University Press.
- ↑ a b H. Melchior. 1964. A. Engler's Syllabus der Pflanzenfamilien (12ª edición)
- ↑ The Angiosperm Phylogeny Group. 2003. "An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG II". Botanical Journal of the Linnean Society, 141, 399-436. (pdf aquí )
- ↑ Qiu et al. 1999. "The earliest angiosperms: evidence from mitochondrial, plastid and nuclear genomes". Nature 402, 404-407
- ↑ Douglas E. Soltis y Pamela S. Soltis. 2004. Amborella not a "basal angiosperm"? Not so fast! American Journal of Botany 91 (6): 997-1001. (resumen aquí )
- ↑ a b Qiu et al. 2005. "Phylogenetic Analyses of Basal Angiosperms Based on Nine Plastid, Mitochondrial, and Nuclear Genes". International Journal of Plant Sciences, 166, 815–842. (resumen aquí )
- ↑ Error en la cita: El elemento
<ref>no es válido; pues no hay una referencia con texto llamadaAPW - ↑ Taylor S. Feild, Nan Crystal Arens, y Todd E. Dawson. 2003. "The Ancestral Ecology of Angiosperms: emerging perspectives from extant basal lineages". Int. J. Plant Sci. 164 (3 Suppl.):S129-S142. (resumen aquí )
Bibliografía
Morfología y General
- Judd, W. S. Campbell, C. S. Kellogg, E. A. Stevens, P.F. Donoghue, M. J. 2002. Plant systematics: a phylogenetic approach, Second Edition. Sinauer Axxoc, Estados Unidos.
Sistemática
- The Angiosperm Phylogeny Group. 2003. "An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG II". Botanical Journal of the Linnean Society, 141, 399-436. (pdf aquí )
Evolución
- Willis y McElwain. 2002. "Flowering Plant Origins". en: The Evolution of Plants, capítulo 6.
- Raven. 2004. "Evolution of the Angiosperms". en : Biology of Plants, capítulo 20 (pdf aquí )
Lecturas recomendadas
- Pamela S. Soltis, Peter K. Endress, Mark W. Chase, Douglas E. Soltis. 2005. Phylogeny & Evolution of Angiosperms. Sinauer Associates, Estados Unidos. 370 pp.
Enlaces externos
 Wikimedia Commons alberga contenido multimedia sobre Magnoliophyta. Commons
Wikimedia Commons alberga contenido multimedia sobre Magnoliophyta. Commons- Wikimedia Commons alberga contenido multimedia sobre MagnoliophytaCommons.
- P. F. Stevens, 2001 en adelante, Angiosperm Phylogeny Website. Versión 8, junio del 2007, y actualizado desde entonces. http://www.mobot.org/MOBOT/research/APweb/





Wikimedia foundation. 2010.