- Endosimbiosis seriada
-
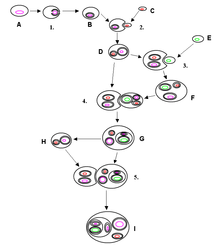 Endosimbiosis seriada
Endosimbiosis seriada
La Endosimbiosis seriada (Serial Endosymbiosis Theory) o teoría endosimbiótica, describe la aparición de las células eucariotas como consecuencia de la sucesiva incorporación simbiogenética de diferentes bacterias de vida libre (procariotas), tres en el caso de animales y hongos y cuatro en el caso de los vegetales.
La endosimbiósis seriada fue propuesta por Lynn Margulis en diferentes artículos y libros: On origen of mitosing cells (1967), Origins of Eukaryotic Cells (1975) y Symbiosis in Cell Evolution (1981),[1] llegándose a conocer por el acrónimo inglés SET (Serial Endosymbiosis Theory). En la actualidad se acepta que las eucariotas surgieron como consecuencia de los procesos simbiogenéticos descritos por Margulis, una vez ha quedado demostrado el origen simbiogenético de las mitocondrias y los cloroplastos de los eucariontes.[2] [3]
Lynn Margulis publicó un artículo en The Biological Bulletin que prueba la incorporación simbiótica de una espiroqueta para formar los flagelos y cilios de los eucariontes,[4] único paso sobre el que, al día de hoy, existen discrepancias.[5]
Contenido
Antecedentes del origen simbiogenético de las eucariotas
En 1883, el biólogo alemán Andreas Schimper propuso que la capacidad fotosintética de las células vegetales podía proceder de cianobacterias aun presentes en la naturaleza y con iguales capacidades.
A principios del siglo XX, en 1909, el ruso Kostantin S. Mereschovky presentó la hipótesis según la cual el origen de los cloroplastos tendría su origen en procesos simbióticos.[6]
Las teorías de Margulis fueron prefiguradas ya en el siglo xix por el naturalista ruso Konstantin Merezhkovsky (1855-1921). Este científico olvidado, que nació antes de la publicación de El origen de las especies y tenía ya 27 años cuando murió Darwin, fue el primer autor que propuso la extravagante idea de la simbiogénesis, según la cual algunos órganos, e incluso algunos organismos, no surgían en la evolución por el gradual mecanismo de la selección natural, sino mediante asociaciones simbióticas entre una especie animal o vegetal y algún tipo de microbio. Merezhkovsky llegó a postular que el núcleo de la célula eucariota provenía de un antiguo microorganismo, lo que posiblemente es erróneo, al menos dicho así, sin más matices. En cualquier caso, sus ideas no tuvieron la menor repercusión.
Javier Sampedro, Deconstruyendo a Darwin.[7]Merezhkovsky llegó a proponer que el núcleo de las células eucariotas tuvo su origen en una bacteria de vida libre; lo que hoy se considera erroneo,[8] [9] pero que lo convierte en uno de los primeros en proponer el origen simbiótico de las células eucariotas. Los trabajos de Merezhkovsky pasaron inadvertidos. Años después, Ivan Wallin (anatomista estadounidense) llegó a la misma conclusión, publicando en 1927 el libro Simbiosis y el origen de las especies. En este caso, sus conclusiones fueron tenidas por absurdas, costándole su prestigio profesional.
En Francia, el biólogo Paul Portier, en 1918, también llegó a conclusiones parecidas sobre el origen simbiótico de las eucariotas. En este caso, Portier sufrió los ataques del entonces influyente microbiólogo August Lumiére.[10]
Estos trabajos, minusvalorados en su tiempo, permanecieron olvidados hasta que Margulis, apoyándose en ellos, poniendo énfasis en las capacidades de las bacterias y la potencialidad de la simbiosis, formulara en 1967 la teoría endosimbiótica.[11]
La teoría
 Modelos quiméricos del origen de Eukarya.[12] [13] Arriba: 1-Fusión, 2-Simbiosis, 3-Ingestión y endosimbiosis. Abajo: Teoría de la endosimbiosis serie: 4-Fusión de una arquea y una espiroqueta, 5-Adquisición de mitocondrias. 6-Adquisición de cloroplastos.
Modelos quiméricos del origen de Eukarya.[12] [13] Arriba: 1-Fusión, 2-Simbiosis, 3-Ingestión y endosimbiosis. Abajo: Teoría de la endosimbiosis serie: 4-Fusión de una arquea y una espiroqueta, 5-Adquisición de mitocondrias. 6-Adquisición de cloroplastos.La teoría endosimbiótica describe el paso de las células procariotas (bacterias o arqueas, no nucleadas) a las células eucariotas (células nucleadas constituyentes de todos los pluricelulares) mediante incorporaciones simbiogenéticas.
Margulis describe este paso en una serie de tres incorporaciones mediante las cuales, por la unión simbiogenética de bacterias, se originaron las células que conforman a los individuos de los otros cuatro reinos (protistas, animales, hongos y plantas).
Según la estimación más aceptada, hace 2.000 millones de años (aunque una horquilla posible podría descender a la cifra de 1.500 millones de años) la vida la componían multitud de bacterias diferentes, adaptadas a los diferentes medios. Margulis destacó también, la que debió ser una alta capacidad de adaptación de estas bacterias al cambiante e inestable ambiente de la Tierra en aquella época. Hoy se conocen más de veinte metabolismos diferentes usados por las bacterias frente a los dos utilizados por los pluricelulares: el aeróbico, que usa el oxígeno como fuente de energía -único metabolismo utilizado por los animales- y la fotosíntesis -presente en las plantas-. Para Margulis, tal variedad revela las dificultades a las que las bacterias se tuvieron que enfrentar y su capacidad para aportar soluciones a esas dificultades.
A mediados de los sesenta, Margulis formuló lo que se conoce como «Teoría de la endosimbiosis serial», que propone que la primera célula eucariota de la Tierra, aquella célula de la que provenimos todos los animales y las plantas, se formó mediante la fusión de tres bacterias preexistentes completas, con los genes de cada una incluidos, por supuesto. Una de esas bacterias aportó los andamios de microtúbulos, otra ciertas capacidades metabólicas peculiares y la tercera (que se sumó más tarde a las otras dos) se convirtió en las actuales mitocondrias. Esa célula eucariota primigenia empezó a proliferar, y una de sus descendientes sufrió aún otra experiencia traumática: se tragó a una bacteria fotosintética de la que provienen los actuales cloroplastos.
Javier Sampedro, Deconstruyendo a Darwin.[14]Primera incorporación simbiogenética:
En primer lugar, un tipo de bacteria amante del azufre y del calor, llamada arqueobacteria fermentadora (o termoacidófila), se fusionó con una bacteria nadadora. Juntos, los dos componentes integrados de la fusión se convirtieron en el nucleocitoplasma, la sustancia base de los ancestros de las células animales, vegetales y fúngicas. Este temprano protista nadador era, como sus descendientes actuales, un organismo anaerobio. Envenenado por el oxígeno, vivía en arenas y lodos donde abundaba la materia orgánica, en grietas de las rocas, en charcos y estanques donde este elemento estaba ausente o era escaso.
Lynn Margulis, Una revolución en la Evolución, Cap.: Individualidad por incorporación.[15]- Una bacteria consumidora de azufre, que utilizaba el azufre y el calor como fuente de energía (arquea fermentadora o termoacidófila), se habría fusionado con una bacteria nadadora (espiroqueta) habiendo pasado a formar un nuevo organismo y sumaría sus características iniciales de forma sinérgica (en la que el resultado de la incorporación de dos o más unidades adquiere mayor valor que la suma de sus componentes). El resultado sería el primer eucarionte (unicelular eucariota) y ancestro único de todos los pluricelulares. El núcleoplasma de la células de animales, plantas y hongos sería el resultado de la unión de estas dos bacterias.
- A las características iniciales de ambas células se le sumaría una nueva morfología más compleja con una nueva y llamativa resistencia al intercambio genético horizontal. El ADN quedaría confinado en un núcleo interno separado del resto de la célula por una membrana.
- Sobre este primer paso, al día de hoy, todavía existen discrepancias. A finales de los años ochenta y principio de los noventa diversos trabajos no admitían las homologías propuestas entre los flagelos de los eucariontes y de las espiroquetas.[16] [17] [18] [19] Margulis defiende que las asociaciones entre espiroquetas y protistas apoyan su teoría, y "la comparación de genes y genomas arqueobaterianos con secuencias de eucariontes han demostrado la relación filogenética de ambos grupos".[20] No obstante, desde su formulación por Margulis, han surgido innumerables interrogantes. Margulis admite que este es el punto de su teoría con más dificultades para defenderse y Antonio Lazcano, en 2002, previene que para comprender el origen de este primer paso, se acepte o no su origen simbiogenético, "es indispensable secuenciar no sólo los genomas de una gama representativa de protistas sino también reconocer la importancia del estudio de la biología de estos organismos".[20]
- Ya en los años setenta surgió, como alternativa al origen simbiogenético de este primer paso, la hipótesis de que éste se hubiese producido mediante invaginaciones,[21] propuesta que no contradice el paradigma neodarviniano y que, aún hoy, se considera plausible por amplios sectores del mundo académico.
- Recurrentemente se han propuesto diferentes hipótesis, también simbiogéneticas, en las que el propio núcleo sería resultado de la incorporación de otro simbionte, como en el caso de las mitocondrias y los cloroplastos.[22]
Segunda incorporación simbiogenética:
Después de que evolucionara la mitosis en los protistas nadadores, otro tipo de microorganismo de vida libre fue incorporado a la fusión: una bacteria que respiraba oxígeno. Surgieron células todavía más grandes, más complejas. El triplemente complejo respirador de oxígeno (amante del calor y del ácido, nadador y respirador de oxígeno) se volvió capaz de engullir alimento en forma de partículas. Estas células con núcleo, seres complejos y asombrosos que nadaban y respiraban oxígeno, aparecieron por primera vez sobre la Tierra quizá tan pronto como hace unos 2.000 millones de años. Esta segunda fusión, en la que el anaerobio nadador adquirió un respirador de oxígeno, condujo a células con tres componentes cada vez más preparadas para soportar los niveles de oxígeno libre que se acumulaban en el aire. Juntos, el delicado nadador, la arqueobacteria tolerante al calor y al ácido y el respirador de oxígeno, formaban ahora un único y prolífico individuo que produjo nubes de prole.
Margulis, Una revolución en la Evolución, Cap.: Individualidad por incorporación.- Este nuevo organismo todavía era anaeróbico, incapaz de metabolizar el oxígeno, ya que este gas suponía un veneno para él, por lo que viviría en medios donde este oxigeno, cada vez más presente, fuese escaso. En este punto, una nueva incorporación dotaría a este primigenio eucarionte de la capacidad para metabolizar oxigeno. Este nuevo endosombionte, originariamente bacteria respiradora de oxigeno de vida libre, se convertiría en las actuales mitocondrias y peroxisomas presentes en las células eucariotas de los pluricelulares, posibilitando su éxito en un medio rico en oxígeno como ha llegado a convertirse el planeta Tierra. Los animales y hongos somos el resultado de esta segunda incorporación.
Tercera incorporación simbiogenética:
En la adquisición final de la serie generadora de células complejas, los respiradores de oxígeno engulleron, ingirieron, pero no pudieron digerir bacterias fotosintéticas de color verde brillante. La «incorporación» literal tuvo lugar tras una gran lucha en la que las bacterias verdes no digeridas sobrevivieron y la fusión completa prevaleció. Con el tiempo las bacterias verdes se convirtieron en cloroplastos (paso 4, figura 1.1). Como cuarto miembro, estos productivos amantes del sol se integraron con los demás socios anteriormente independientes. Esta fusión final dio lugar a las algas verdes nadadoras. Estas antiguas algas verdes nadadoras no sólo son los ancestros de las células vegetales actuales; todos sus componentes individuales todavía están vivos y en buena forma, nadando, fermentando y respirando oxígeno.
Margulis, Una revolución en la Evolución, Cap.: Individualidad por incorporación.- Esta tercera incorporación originó el Reino vegetal, las recientemente adquiridas células respiradoras de oxígeno fagocitarían bacterias fotosintéticas y algunas de ellas, haciéndose resistentes, pasarían a formar parte del organismo, originando a su vez un nuevo organismo capaz de sintetizar la energía procedente del Sol. Estos nuevos pluricelulares, las plantas, con su éxito, contribuyeron y contribuyen al éxito de animales y hongos.
En la actualidad permanecen las bacterias descendientes de aquellas que debieron, por incorporación, originar las células eucariotas; así como aquellos protistas que no participaron en alguna de las sucesivas incorporaciones.
Presentación y alcance de la teoría
Margulis presentó en 1967 su teoría sobre el origen de las células eucariotas mediante un artículo en la revista Journal of Theoretical Biology: «Origin of Mitosing Cells»;. Antes, diferentes trabajos sobre esa misma teoría le habían sido rechazados en quince ocasiones y fue la directa intervención de su editor James F. DaNelly lo que posibilitase al fin su publicación.
Max Taylor, especializado el protistas, profesor de la Universidad de British Columbia, la bautizó con el acrónimo SET (Serial Endosyrnbiosis Theory), nombre por el que hoy es conocida.
Margulis también tuvo problemas para publicar un segundo texto más extenso ya en forma de libro: Origin of Eukaryotic Cells, el que en 1970 publicara Yale University Press.
Quien no se dedique a la ciencia profesional pensará que, cuando un investigador tiene una idea de ese tipo, le resultará más o menos fácil encontrar un editor que quiera publicársela en una revista técnica. Aunque sólo sea para que otros investigadores la lean y la refuten en el siguiente número. Pero no es así, ni siquiera cuando se trata de ideas más convencionales. En el caso de Margulis, encima, la idea era una absoluta herejía, y sus escasos precedentes, sobre ser arcaicos, no habían causado más que la ruina científica de sus proponentes. cuando no la burla general. El manuscrito fue rechazado quince veces por varias revistas, y sólo pudo publicarse en 1967. tras casi dos años de peregrinación, en el Journal of Theoretical Biology, una especie de último recurso para científicos desesperados (lo cual no le resta un ápice de interés a la revista. como demuestra este caso). […] Dos años después de la publicación de su artículo, Margulis se animó a extender su teoría en un libro y mandó el manuscrito a la prestigiosa editorial de Nueva York Academic Press. La editorial, como es costumbre, dio a leer el texto a terceros expertos, y su dictamen fue tan radicalmente negativo que, tras retener el manuscrito durante cinco meses, se lo devolvió a la autora sin pedir ni perdón.
Javier Sampedro[23]Margulis apoyó su teoría en numerosos datos sobre bioquímica y morfología (investigaciones de otros científicos y de ella misma), también la apoyó en datos paleontológicos y presentó un proceso coherente valiéndose de los descendientes (aun entre nosotros) de aquellas bacterias que plausiblemente lo protagonizaron.
En el momento en que Margulis presenta su teoría, para el evolucionismo únicamente existen animales y plantas, las bacterias solo eran del interés de una rama de la medicina: la bacteriología médica. Y únicamente algunos protozoos y hongos eran clasificados por diversos botánicos erróneamente como plantas.
En este contexto, su trabajo es rechazado por la ortodoxia del evolucionismo; el paso de procariotas a eurocariotas no entrañaba ningún problema especial de comprensión para el neodarwinismo: lo mismo que una especie puede aumentar paulatinamente de tamaño, aumentar su complejidad y adquirir nuevas funcionalidades también paulatinamente; los procariotas habrían aumentado su tamaño, aumentado su complejidad y adquirido nuevas funcionalidades, todo paulatinamente y como consecuencia de errores en la replicación de su ADN.
Cuando Margulis plantea su teoría y surgen los primeros apoyos es cuando el neodarwinismo siente la necesidad de formular una teoría alternativa ajustada a su paradigma, y para ello utiliza el escenario descrito por ella para justificar tan paso: un medio "caótico", inestable y cambiante; un medio que justificaba una alta competencia favorecedora de tan radicales cambios.
La primera teoría alternativa a la de la endosimbiosis seriada formulada por Margulis la postuló Max Taylor (Universidad British Columbia) a principios de los años setenta. Una teoría basada en el modelo darwinista y que bautizó con el nombre de "filiación directa".
La teoría de Taylor, un buen ejemplo de la resistencia heroica del neodarwinismo a aceptar cualquier cosa que no encaje en sus preconcepciones, quedó refutada al poco de formularse, cuando se comprobó que el ADN de las mitocondrias y los cloroplastos es mucho más parecido al material genético de las bacterias que al del genoma nuclear eucariota. El mundo académico se vio forzado a aceptar la parte de la teoría de Margulis que hoy se enseña en todos los libros de texto: que las mitocondrias y los cloroplastos provienen, por simbiosis, de antiguas bacterias de vida libre. La idea convencional, sin embargo, persiste aún gracias a que la teoría de Margulis se suele presentar en una versión edulcorada que no capta el fondo de la cuestión.
Javier Sampedro[24]Mahler, en 1972, rechazó que las espiroquetas y el aparato mitótico fuesen consecuencia de procesos simbióticos considerando que "tanto la síntesis de las proteínas dirigida por el núcleo como la síntesis proteica dirigida por las mitocondrias, así como otras actividades metabólicas se deben a presiones selectivas divergentes". En 1975 Bogart formuló otra hipótesis según la cual, "todo el ADN de las células eucariotas se encontraba originariamente en el área nuclear, rodeado quizás por una membrana nuclear común y posteriormente se separaron bloques de genes procedentes de protonúcleo y se rodearon de sus propias membranas, con lo cual se formaron los plástidos y las mitocondrias".[25]
Desde entonces desde el neodarwinismo se han ofrecido numerosas teorías alternativas.
Existen otras teorías del origen simbiogenético de las eucariotas que también incluyen el núcleo, como en la teoría expuesta por Margulis. Timan Hartman del Ames Research Center de la NASA y Radney Gupta de la Universidad MacMaster defienden ese origen simbiótico del núcleo, pero mediante pasos y procesos diferentes a los defendidos por Margulis.[26] [27]
También existe, y tiene sus seguidores, una versión del paso de procariotas a eurocariotas que sólo admite como posible adquisición simbiótica la capacidad fotosintética; y en todo caso, desde el neodarwinismo, la capacidad motora de las eucariotas (insuficientemente demostrado su origen simbiogenético) se considera producto de mutaciones al azar.
Para Margulis el origen del núcleo se debe a la interacción de los dos primeros simbiontes que elaborarían esas membranas a modo de barrera que impidiera su total fusión.
La SET ha necesitado treinta años para ser aceptada (solo en parte) mayoritariamente como plausible, cuando se ha considerado demostrada la incorporación de tres de los cuatro simbiontes que Margulis propone como componentes de los eucariotas (la adquisición de bacterias nadadoras no se considera probada).
Hoy me sorprende ver cómo en los institutos y en los textos universitarios se enseña una versión diluida, como si se tratara de una verdad descubierta. Encuentro, para mi desgracia aunque no para mi sorpresa, que la exposición es dogmática, lleva a la confusión, no está argumentada con lógica, y con frecuencia es francamente incorrecta
Margulis, Planeta Simbiótico.[28]Margulis no se ha limitado a formular la SET, con ella se acepta como demostrada parte de un hito de la evolución de la vida;[29] desde entonces, intenta extender el alcance de la simbiogénesis a otros ámbitos de la Evolución: la especiación y la adquisición de caracteres y organos de los organismos (Teoría simbiogenética).
Argumentos que se esgrimieron a favor y en contra de la teoría
Argumentos a favor
La evidencia de que las mitocondrias y los plastos surgieron a través del proceso de endosimbiosis son las siguientes:
- El tamaño de las mitocondrias es similar al tamaño de algunas bacterias.
- Las mitocondria y los cloroplastos contienen ADN bicatenario circular cerrado covalentemente - al igual que los procariotas- mientras que el núcleo eucariota posee varios cromosomas bicatenarios lineales.
- Están rodeados por una doble membrana, lo que concuerda con la idea de la fagocitosis: la membrana interna sería la membrana plasmática originaria de la bacteria, mientras que la membrana externa correspondería a aquella porción que la habría englobado en una vesícula.
- Las mitocondrias y los cloroplastos se dividen por fisión binaria al igual que los procariotas (los eucariotas lo hacen por mitosis). En algunas algas, tales como Euglena, los plastos pueden ser destruidos por ciertos productos químicos o la ausencia prolongada de luz sin que el resto de la célula se vea afectada. En estos casos, los plastos no se regeneran.
- En mitocondrias y cloroplastos los centros de obtención de energía se sitúan en las membranas, al igual que ocurre en las bacterias. Por otro lado, los tilacoides que encontramos en cloroplastos son similares a unos sistemas elaborados de endomembranas presentes en cianobacterias.
- En general, la síntesis proteica en mitocondrias y cloroplastos es autónoma.
- Algunas proteínas codificadas en el núcleo se transportan al orgánulo, y las mitocondrias y cloroplastos tienen genomas pequeños en comparación con los de las bacterias.. Esto es consistente con la idea de una dependencia creciente hacia el anfitrión eucariótico después de la endosimbiosis. La mayoría de los genes en los genomas de los orgánulos se han perdido o se han movido al núcleo. Es por ello que transcurridos tantos años, hospedador y huésped no podrían vivir por separado.
- En mitocondrias y cloroplastos encontramos ribosomas 70s, característicos de procariotas, mientras que en el resto de la célula eucariota los ribosomas son 80s.
- El análisis del RNAr 16s de la subunidad pequeña del ribosoma de mitocondrias y plastos revela escasas diferencias evolutivas con algunos procariotas.
- Una posible endosimbiosis secundaria (es decir, implicando plastos eucariotas) ha sido observado por Okamoto e Inouye (2005). El protista heterótrofo Hatena se comporta como un depredador e ingiere algas verdes, que pierden sus flagelos y citoesqueleto, mientras que el protista, ahora un anfitrión, adquiere nutrición fotosintética, fototaxia y pierde su aparato de alimentación.
Las bacterias, fusionadas en simbiosis, nos dejan pistas de su anterior independencia. Tanto las mitocondrias como los plastos son bacterianos en su tamaño y forma. Todavía más importante es que estos orgánulos se reproducen de manera que hay muchos presentes a la vez en el citoplasma pero nunca dentro del núcleo. Ambos tipos de orgánulos, los plastos y las mitocondrias, no sólo proliferan dentro de las células sino que se reproducen de forma distinta y en momentos distintos a los del resto de la célula en la que residen. Ambos tipos, probablemente 1.000 millones de años después de su fusión inicial, retienen sus propias reservas reducidas de ADN. Los genes del ácido desoxirribonucleico (ADN) de los ribosomas de las mitocondrias todavía recuerdan sorprendentemente a los de las bacterias respiradoras de oxígeno que viven actualmente por su cuenta. Los genes ribosómicos de los plastos son muy parecidos a los de las cianobacterias. A principios de los setenta, cuando se compararon por primera vez las secuencias de nucleótidos del ADN de los plastos de las células algales con las secuencias de las cianobacterias de vida libre, ¡se descubrió que el ADN del cloroplasto era mucho más parecido al ADN de la cianobacteria que al ADN del núcleo de la propia célula algal!
Margulis, Planeta simbiótico.[30]Argumentos en contra
- Las mitocondrias y los plastos contienen intrones, una característica exclusiva del ADN eucariótico. Por tanto debe de haber ocurrido algún tipo de transferencia entre el ADN nuclear y el ADN mitocondrial/cloroplástico.
- Ni las mitocondrias ni los plastos pueden sobrevivir fuera de la célula. Sin embargo, este hecho se puede justificar por el gran número de años que han transcurrido: los genes y los sistemas que ya no eran necesarios fueron suprimidos; parte del ADN de los orgánulos fue transferido al genoma del anfitrión, permitiendo además que la célula hospedadora regule la actividad mitocondrial.
- La célula tampoco puede sobrevivir sin sus orgánulos: esto se debe a que a lo largo de la evolución gracias a la mayor energía y carbono orgánico disponible, las células han desarrollado metabolismos que no podrían sustentarse solamente con las formas anteriores de síntesis y asimilación.
Las membranas internas de las células eucariotas, sobre todo la membrana nuclear y el retículo endoplasmático, se explican mejor como invaginaciones. Como el cromosoma de los procariotas se halla unido a la membrana celular durante su división (Stanier, Doudoroff & Adelberg, 1970), una invaginación de esta parte de la membrana celular situaría automáticamente a los cromosomas en el interior del núcleo. Las conexiones y la semejanza molecular entre los flagelos y centríolos podría explicarse suponiendo que en los primeros eucariotas la membrana nuclear continuaría unida a la membrana celular externa durante la evolución simultánea del aparato mitótico y de los flagelos. Podría explicarse la existencia de DNA en el cuerpo basal del flagelo por adición de un plásmido, tal como han sugerido Raff & Mahler.
Stebbins, Evolución", 1980.[31]Consideración actual
En la actualidad se considera probada la interacción simbiogenética de tres de los cuatro simbiontes propuestos por Margulis. Existiendo discrepancias sobre la incorportación de las espiroquetas, día a día va adquiriendo mayor consenso esa primera incorporación propuesta por Margulis.
En mi opinión, los abrumadores datos de Gupta demuestran por encima de toda duda razonable la parte más radical de la teoría de Margulis. La célula eucariota original, incluso antes de la adición de las mitocondrias y los cloroplastos, no se formó por evolución darwiniana desde una arquea, como siguen sosteniendo Woese, Taylor y muchos otros científicos —y como explican pertinazmente cada vez más libros de texto—, sino por la fusión de una arquea con otra bacteria, una gram-negativa para ser exactos. Los genomas de ambas están todavía ahí, en cada célula de su cuerpo, amable lector, constituyendo su sacrosanto genoma nuclear. A esa fusión simbiótica inicial se añadieron luego las mitocondrias, y después los cloroplastos.
Sampedro, 2002, Deconstruyendo a Darwin[32]Actualmente se consideran posibles alternativas a la propuesta por Margulis para explicar ese primer paso. Alternativas más acorde con el paradigma neodarwinista, mediante procesos graduales de transformación de la célula originaria, y alternativas que contemplan, igualmente, el origen simbiogenético:
La presencia de genes (eu)bacterianos en el núcleo de Giardia sugiere que la ausencia de mitocondrias en este y otros diplomónadas representa una pérdida secundaria. Esta posibilidad se ve apoyada por la presencia de mitocondrias vestigiales en los microsponidios, cuya cercanía filogenética con los hongos es conocida (Williams et al. 2002). Es decir, si las espiroquetas son de verdad los ancestros de la movilidad eucariótica y la mitosis (Margulis 1993), su adquisición como endosimbiontes no necesariamente tiene que haber tenido lugar antes de las mitocondrias. Es decir, me parece que la condición eucarionte, que está ligada en forma directa a la adquisición de endosimbiontes, comenzó cuando el ancestro de las mitocondrias contemporáneas penetró en el interior de una arqueobactenia y comenzó una transferencia masiva de genes eubacterianos al genoma del hospedero. Aunque ésta es una versión ortodoxa que rescata buena parte de los postulados iniciales de Margulis, permite explicar la presencia de genes de mitocondnia y de mitocondnias vestigiales en eucaniontes antiguos cuya anaerobosis es de origen secundario. Esta versión de la teoría endosimbiótica no contradice el papel de las espiroquetas en el origen de la motilidad eucariótica, pero al mismo tiempo deja abierta la posibilidad de que el aparato mitótico y, en general, la motilidad eucariótica, tengan un origen que nada tiene que ver con las espiroquetas —a pesar del enorme atractivo que en lo personal me despierta esta imagen.
Antonio Lazcano Araujo.[33]En todo caso:
Hoy en día existen pruebas concluyentes a favor de la teoría de que la célula eucariota moderna evolucionó en etapas mediante la incorporación estable de las bacterias. Diferentes aportaciones justifican el origen de los cloroplastos y las mitocondrias a partir de éstas.
Isabel Esteve, UAB.[34]Lynn Margulis en los próximos meses (a principios del año 2010) publicó un artículo científico en Biological Bulletin con sus últimos descubrimentos sobre los cirios de las células eucariotas que probarían su orígen simbiotico y el origen de la mitosis: «Existen formas intermedias en las que no se puede ver si son cilios o espiroquetas (bacterias helicoidales). Ahora hemos obtenido cada paso, y eso es noticia.»
Ahora tenemos cada paso y no hay eslabones perdidos en este tipo de simbiogénesis en la formación de cilios. Formamos relaciones con las espiroquetas pero cada paso está analizado. Para comprender este esquema hay que elegir cada elemento y ponerlo en orden porque en la naturaleza este orden no existe. Empezamos con un esquema teórico y en la vida tenemos ya exactamente lo que hemos predicho y todo va en la misma dirección.
Referencias
- ↑ L. Margulis (1967), "On origen of mitosing cells", Journal of theoretical biology , 14 (3): 225. L. Margulis (1975), Origins of Eukaryotic Cells. Yale University Press, New Haven. L. Margulis (1981). Symbiosis in Cell evolution. Freeman, New York. L. Margulis (1992), Symbiosis in Cell Evolution: Microbial Communities in the Archean and Proterozoic Eons, Freeman, New York.
- ↑
Hoy en día existen pruebas concluyentes a favor de la teoría de que la célula eucariota moderna evolucionó en etapas mediante la incorporación estable de las bacterias. Diferentes aportaciones justifican el origen de los cloroplastos y las mitocondrias a partir de éstas.
Isabel Esteve, Discurso de presentación de Lynn Margulis en el acto de investidura doctora honoris causa UAB - ↑ L. Margulis and D. Sagan (2001). «Marvellous microbes». Resurgence 206: pp. 10–12.
- ↑ Entrevista con Lynn Márgulis, Muchas de las cosas que nadie sabe de Darwin han pasado en Chile, SINC, 27/11/2009
- ↑ Mujeres en Biología: Lynn Margulis. Universidad de Zaragoza (UNIZAR)
- ↑ Margulis, 2002, p. 35.
- ↑ p. 36-37.
- ↑ "No estoy de acuerdo con Hartman o Merezhkovsky en que el núcleo tenga orígenes simbióticos. Que yo sepa, en el mundo microbiano no hay bacterias nucleiformes de vida libre". Margulis, 2002, p. 57.
- ↑ Sampedro, 2002, p. 37.
- ↑ Margulis, Una revolución en la Evolución, 2003, p. 167.
- ↑ Borges dijo que todo genio crea a sus antecesores, y Margulís no es una excepción. Ella misma ha reconocido: «Prácticamente todo aquéllo en lo que trabajo ahora fue anticipado por estudiosos y naturalistas desconocidos». Sampedro 2002, Pg.: 35
- ↑ T.M. Embley and W. Martin (2006), Eukaryotic evolution, changes and challenges, Nature Reviews, Vol 440:30, March 2006, doi:10.1038/nature04546
- ↑ Poole A, Penny D (2007). «Evaluating hypotheses for the origin of eukaryotes». Bioessays 29 (1): pp. 74-84. PMID 17187354.
- ↑ P. 39.
- ↑
En las plantas y en los animales el núcleo delimitado por una membrana desaparece cada vez que la célula se multiplica por división; la membrana se disuelve y los cromosomas se hacen visibles. La cromatina, el material que se tiñe de rojo y del que están hechos los cromosomas, se enrolla formando estructuras fácilmente observables. La explicación de libro de texto de este proceso es que la cromatina se condensa formando cromosomas visibles y contables cuyo número es representativo de la especie en cuestión. La danza de los cromosomas, que más tarde desaparecen pasando a ser cromatina débilmente enrollada a medida que reaparece la membrana nuclear, constituye los pasos del proceso de división celular de la mitosis. La mitosis, con sus numerosas variaciones en las células nucleadas de los protoctistas y los hongos, evolucionó en los más tempranos organismos con núcleo.
Margulis, Una revolución en la Evolución, p. 98. - ↑ Bermudes, D., Fracek, S. P. Jr., Laursen, R. A., Margulis, L., Obar, R. and Tzertzinis, G. (1987). "Tubulinlike protein from Spirochaeta bajacaliforniensis", in Annals of the New York Academy of Sciences: Endocytobiology III. New York, The New York Academy of Sciences. 503: 515-527.
- ↑ Barth, A. L., Stricker, J. A. and Margulis, L. (1991). "Search for Eukaryotic Motility Proteins in Spirochetes - Immunological Detection of a Tektin-Like Protein in Spirochaeta-Halophila". BioSystems, V24(N4): 313-319.
- ↑ Bermudes, D., Hinkle, G. and Margulis, L. (1994). "Do Prokaryotes Contain Microtubules". Microbiological Reviews, V58(N3): 387-400.
- ↑ Munson, D., Obar, R., Tzertzinis, G. and Margulis, L. (1993). "The Tubulin-Like S1 Protein of Spirochaeta Is a Member of the Hsp65 Stress Protein Family". BioSystems, V31(N2-3): 161-167.
- ↑ a b Lazcano, 2002, p. 197.
- ↑
Las membranas internas de las células eucariotas, sobre todo la membrana nuclear y el retículo endoplasmático, se explican mejor como invaginaciones. Como el cromosoma de los procariotas se halla unido a la membrana celular al menos durante su división (Stanier, Doudoroff & Adelberg, 1970), una invaginación de esta parte de la membrana celular situaría automáticamente a los cromosomas en el interior del núcleo. Las conexiones y la semejanza molecular entre los flagelos y centríolos podrían explicarse suponiendo que en los primeros eucariotas, la membrana nuclear continuaría unida a la membrana celular externa durante la evolución simultánea del aparato mitótico y de los flagelos. Podría explicarse la existencia de DNA en el cuerpo basal del flagelo por adición de un plásmido, tal como han sugerido Raff & Mahler.
Stebbins, 1980. - ↑
La idea de que el núcleo mismo sea el resultado de una simbiosis es una hipótesis recurrente, que se propuso, en forma vaga y poco precisa, en 1909 por el mismo Merezhkovsky (Margulis 1993). Aunque la continuidad funcional y anatómica entre el medio intranuclear y el citoplasma eucarionte está perfectamente demostrada, de cuando en cuando se revive en la bibliografía la posibilidad de que el núcleo provenga, al igual que las mitocondrias y los cloroplastos, de un simbionte microbiano (Pickett-Heaps 1974, Hartman 1975, 1984, Hart¬man y Fedorov 2002, Sogin 1991, Lake y Rivera 1994, Gupta y Golding 1996, Vellai et al. 1998, Horiike et al. 2001). Lazcano, 2002, p. 191.
- ↑ Sampedro 2002, Pg.: 39.
- ↑ Sampedro 2002, Pg.: 41
- ↑ Sánchez Acedo, 2000, p. 43.
- ↑
Radney Gupta, de la Universidad McMaster, continúa presentando pruebas detalladas, aunque quizá arcanas, de la naturaleza «quimérica» de las más tempranas células nucleadas. Los argumentos de Gupta están basados exclusivamente en el análisis de la secuencia de aminoácidos de muchas proteínas indispensables. Utiliza términos diferentes y criterios distintos pero nuestras ideas básicas son las mismas: la fusión arqueo-eubacterial generó los primeros ancestros celulares con núcleo limitado por una membrana. […] Quizá bacterias distintas de las espiroquetas, tales como un tipo de eubacteria verde no sulfúrica según conjetura Gupta, se infiltraron hace mucho tiempo en otras células.
Margulis, 2002, p. 57, 63. - ↑
La solución propuesta por Hartman y Fedorov es acudir a un tercer hombre. La célula eucariota (esto es, su cuerpo central) no se formó por la fusión de dos microbios, sino de tres: una bacteria, una arquea y lo que ellos llaman un cronocito: un tercer microorganismo que aportó los 347 genes responsables de la eucariotez.
Sampedro, 2002, p. 59. - ↑ p. 44
- ↑
Tras el origen de la vida celular, el primer avance en grado fue el de la célula eucaniota. Su origen puede ser considerado como el avance más significativo de la evolución orgánica, ya que hasta que las células no adquirieron la complejidad y el grado de división del trabajo entre membranas y orgánulos que se halla asociado al estado eucariota, no pudieron evolucionar los organismos pluricelulares con células y tejidos diferenciados
Stebbins, 1980, Evolución, p. 381. - ↑ Margulis, 2002, p. 53.
- ↑ Stebbins, 1980, p. 386.
- ↑ Sampedro, 2002, p. 47.
- ↑ Lazcano, 2003, p. 197.
- ↑ Isabel Esteve, Discurso de presentación de Lynn Margulis en el acto de investidura doctora honoris causa UAB
Bibliografía
- Margulis, Lynn (2002). Planeta Simbiótico. Un nuevo punto de vista sobre la evolución.. Victoria Laporta Gonzalo (trad.). Madrid: Editorial Debate.
- Margulis, Lynn; Dorion Sagan (2003). Captando Genomas. Una teoría sobre el origen de las especies.. Ernst Mayr (prólogo). David Sempau (trad.) (1ª edición). Barcelona: Editorial Kairós. ISBN 84-7245-551-3.
- Margulis, Lynn; Dorion Sagan (1995). Microcosmos: Cuatro mil millones de años de evolución desde nuestros ancestros microbianos. Lewis Thomas, Ricard Guerrero (trad.) (2ª ed. edición). Tusquets Editores. pp. 317 páginas. ISBN 9788472238428.
- Margulis, Lynn (2003). Una revolución en la evolución: Escritos seleccionados (1ª ed. edición). Universitat de València. pp. 374 págs.. ISBN 9788437054940.
- Dobzhansky, Theodosius; Francisco J. Ayala, G. Ledyard Stebbins, James W. Valentine; trad.: Montserrat Aguadé (1980). Evolución (Ediciones Omega edición).
- Mujeres en Biología: Lynn Margulis. Universidad de Zaragoza (UNIZAR)
- Lazcano Araujo, Antonio, El origen del nucleocitoplasma. Breve historia de una hipótesis cambiante, incluido en Una revolución en la Evolución, Universitat València, 2003, ISBN 9788437054940
- Bruce Alberts, Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts y Peter Walter, Molecular Biology of the Cell, Garland Science, Nueva York, 2002. ISBN 0-8153-3218-1.
- Jeffrey L. Blanchard and Michael Lynch (2000), "Organellar genes: why do they end up in the nucleus?", Trends in Genetics, '16 (7), pp. 315-320.[1]
- Paul Jarvis (2001), "Intracellular signalling: The chloroplast talks!", Current Biology, 11 (8), pp. R307-R310. [2]
- Fiona S.L. Brinkman, Jeffrey L. Blanchard, Artem Cherkasov, Yossef Av-Gay, Robert C. Brunham, Rachel C. Fernandez, B. Brett Finlay, Sarah P. Otto, B.F. Francis Ouellette, Patrick J. Keeling, Ann M. Rose, Robert E.W. Hancock y Steven J.M. Jones (2002,) Evidence That Plant-Like Genes in Chlamydia Species Reflect an Ancestral Relationship between Chlamydiaceae, Cyanobacteria, and the Chloroplast Genome Res., 12: pp 1159 – 1167. [3]
- Okamoto, N. y Inouye, I. (2005), "A Secondary Symbiosis in Progress?", Science, 310, p. 287
- Gabaldón T. et al (2006), "Origin and evolution of the peroxisomal proteome", Biology Direct, 1 (8),.
- Sampedro, Javier (2002). Desconstruyendo a Darwin. Ginés Morata (prólogo). (1ª edición). Barcelona: Editorial Crítica.
- Caridad Sánchez Acedo, Discurso de ingreso en la Academia de Ciencias Exactas, Físicas, Químicas y Naturales de Zaragoza, 2000.
- Sandín, Máximo, (2006), Pensando la evolución, pensando la vida, Ediciones Crimentales S. L. Murcia, ISBN 84-935141-0-1
- Youtube. Acto de investidura doctora honoris causa por la Universidad Autònoma de Barcelona UAB. 1/02/2006
- Acto de investidura doctora honoris causa por la Universidad Autònoma de Barcelona UAB. 1/02/2006
Véase también
- Lynn Margulis
- Hipótesis del hidrógeno
- Simbiogénesis
- Teoría simbiogenética
- Eucariogénesis viral (hipótesis del origen endosimbiótico del núcleo celular).
- Genoma mitocondrial
- Teoría autógena
- Hipótesis fagotrófica

Wikimedia foundation. 2010.