- Archaea
-
Archaea
?Archaea
Rango fósil: Arcaico - Holoceno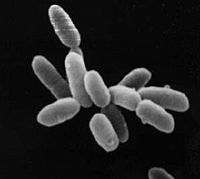
Halobacteria sp. cepa NRC-1, cada célula mide aproximadamente 5 μm de longitud.Clasificación científica Dominio: Archaea
Woese, Kandler & Wheelis, 1990Filos Las arqueas (Archaea) son microorganismos unicelulares. Al igual que las bacterias, las arqueas carecen de núcleo y son por tanto procariontes. Sin embargo, las diferencias a nivel molecular entre arqueas y bacterias son tan fundamentales que se las clasifica en grupos distintos. De hecho, estas diferencias son mayores de las que hay, por ejemplo, entre una planta y un animal. Actualmente se considera que las arqueas están filogenéticamente más próximas a los eucariontes que a las bacterias.
Las arqueas fueron descubiertas originariamente en ambientes extremos, pero desde entonces se las ha hallado en todo tipo de hábitats y podrían contribuir hasta el 20% de la biomasa microbiana oceánica.[1] Son particularmente comunes en los océanos y pueden ser uno de los más abundantes grupos de organismos en el planeta.[2] Algunas especies son hipertermófilas y pueden sobrevivir y prosperar a temperaturas ligeramente superiores a los 100°C y se las encuentra en géiseres, fuentes hidrotermales y pozos de petróleo.[3] Otras extremófilas se encuentran en agua hiper-salina, ácida o alcalina. Sin embargo, otras son mesófilas o psicrófilas y prosperan en ambientes tales como marismas, aguas residuales, agua de mar y el suelo. A las metanógenas se las puede encontrar en el tracto digestivo de animales tales como rumiantes, termitas y seres humanos. No se conocen patógenos para los seres humanos,[4] [5] aunque se ha propuesto alguna relación entre los metanógenos y los trastornos periodontales humanos.[6]
Archaea constituye uno de los dominios en los que se dividen los seres vivos. Antiguamente se clasificaban como perteneciendo al reino Monera en la taxonomía tradicional de los cinco reinos. En 1990 se propuso considerarlos un dominio separado, según el sistema de tres dominios de Carl Woese.[7] [8]
Contenido
Características morfológicas y bioquímicas
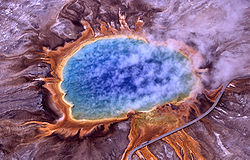 Las arqueas fueron inicialmente detectadas en ambientes extremos, tales como manantiales volcánicos.
Las arqueas fueron inicialmente detectadas en ambientes extremos, tales como manantiales volcánicos.
Las arqueas individuales tienen un diámetro comprendido entre 0,1 y 15 μm, mientras que algunos agregados o filamentos celulares llegan a tener una longitud de hasta 200 μm. Presentan diversas formas: esférica, cilíndrica, espiral, lobular, triangular, rectangular, irregular, pleomórfica, etc. Recientemente, se ha descubierto (en piscinas hipersalinas) una especie de forma cuadrada y plana.[9] Algunas arqueas tienen flagelos y son móviles.
Las arqueas al igual que las bacterias, no tienen membranas internas que delimiten orgánulos. Como todos los organismos presentan ribosomas, pero a diferencia de los encontrados en las bacterias que son sensibles a ciertos agentes químicos inhibidores, los de las arqueas (y también los de Eukarya) no lo son, lo que puede sugerir una relación cercana entre Archaea y Eukarya. La membrama celular tiene una estructura similar a la de las demás células, pero su composición química es única. Casi todas las arqueas poseen una pared celular (algunos Thermoplasma son la excepción) de composición característica, por ejemplo, no contienen peptidoglicano (mureína) como en las bacterias. Pueden clasificarse como Gram positivas (la mayoría tiene una capa homogénea y gruesa de seudopeptidoglicano) o Gram negativas.
En resumen, la célula de las arqueas tiene las mismas estructuras que el resto de los organismos pero se construye con compuestos químicos diferentes. Debido a estas diferencias, las arqueas exhiben una alta resistencia contra los antibióticos y los agentes líticos.
Como en todos los procariontes, las células de las arqueas no tienen núcleo y presentan un sólo cromosoma circular. Elementos extracromosómicos, tales como plásmidos también se encuentran en las arqueas. Sus genomas son de pequeño tamaño, sobre 2-4 millones de pares de bases. También es característica la presencia de ARN-polimerasas de constitución compleja y un gran número de nucleótidos modificados en los ácidos nucleicos ribosomales. Por otra parte, el ADN se empaqueta en forma de nucleosomas, como en los eucariontes gracias a proteínas semejantes a la histona, y algunos genes poseen intrones como en los eucariontes.
Las arqueas pueden ser aerobias, aerobias facultativas o anaerobias estrictas, autótrofas o heterótrofas. Varias especies son productores primarios usando el dióxido de carbono como fuente única de carbono y obteniendo energía por la oxidación de sustancias inorgánicas tales como azufre o hidrógeno, o por la reducción de azufre o nitrato. Aunque no se conocen especies que fijen carbono a través de la fotosíntesis, algunas arqueas usan iones activados por la luz, tales como la bacteriorodopsina y halorodopsina, que generan gradientes iónicos que son convertidos en adenosín trifosfato (ATP).[10]
Pueden reproducirse por fisión binaria o múltiple, fragmentación o gemación.
Hábitats
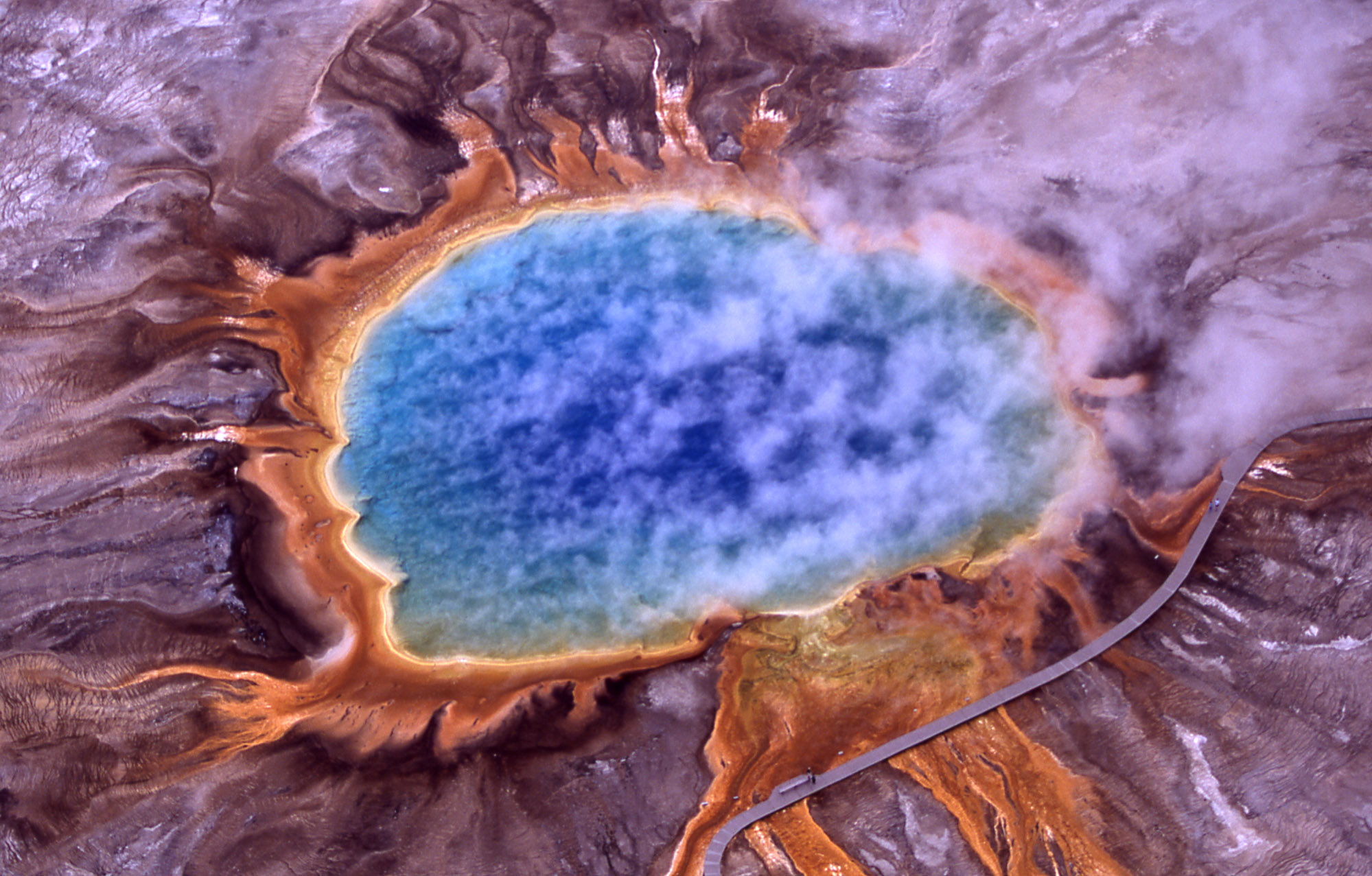
 En los géiseres del parque Yellowstone proliferan las arqueas.
En los géiseres del parque Yellowstone proliferan las arqueas.Según el hábitat preferido, podemos dividir a las arqueas en tres grupos. Estos grupos no coinciden necesariamente con filogénesis moleculares, ni son necesariamente completos, ni mutuamente exclusivos. No obstante, son un punto de partida útil para estudios más detallados.
- Halófilos. Viven en ambientes extremadamente salinos. Halococcus y Halobacterium solo viven en medios con más del 12% de sal y pueden sobrevivir a concentraciones del 32% de sal.[3] [11]
- Termo-acidófilos. Necesitan temperaturas de más de 60-80 °C, y algunas especies también un pH bajo, de 1-3. Sulfolobus acidocaldarius oxida el azufre y vive en las fuentes termales del parque Yellowstone. Thermoplasma se encuentra en escombreras de carbón encendidas. Pyrolobus fumarii es el organismo más termófilo de todos los conocidos con una temperatura máxima de crecimiento de 113 °C. El organismo más acidófilo, Picrophilus, puede crecer a un pH de -0,06.[3]
- Metanógenos. Viven en ambientes anaeróbicos y producen metano. Se pueden encontrar en sedimentos o en los intestinos de animales.[11] Se han encontrado metanógenos vivos en muestras de hielo glaciar de Groenlandia tomadas a 3 km de profundidad.[12]
Recientemente, varios estudios han demostrado que las arqueas proliferan, no sólo en ambientes mesofílicos y termofílicos, sino también, a veces en gran número, en ambientes psicrofílicos. Los estudios filogenéticos sugieren que estas arqueas derivan de antepasados termófilos que invadieron diversos hábitats de baja temperatura. Quizás lo más significativo sea la gran cantidad de arqueas encontradas en la mayor parte de de los océanos del mundo, un ambiente predominante frío. Estas arqueas pueden estar presentes en cantidades extremadamente altas (hasta 40% de la biomasa microbiana) aunque hasta el momento ninguno ha podido ser cultivado.[13] No tenemos actualmente casi ninguna información con respecto a la fisiología de estos organismos, lo que significa que sus efectos sobre los ciclos biogeoquímicos globales siguen siendo desconocidos. Un estudio reciente ha demostrado, sin embargo, que un grupo de crenarchaeota marina es capaz de la nitrificación, un rasgo previamente desconocido en las arqueas.[14]
Comparación de Archaea, Bacteria y Eukarya
Archaea es similar a Bacteria en la mayoría de los detalles de la estructura y del metabolismo de la célula, pero difiere sobre todo en la composición de los lípidos de la membrana celular y en los aspectos genéticos. Así, la transcripción y traducción genéticas, los dos procesos centrales de la biología molecular, no presentan las características bacterianas típicas sino que son más similares en muchos aspectos a los encontrados en Eukarya. Por ejemplo, la traducción usa factores de iniciación y de alargamiento del tipo eucarionte y la transcripción implica proteínas de unión TATA y TFIIB como en los eucariontes. Muchos genes ARNr y ARNt de las arqueas albergan intrones únicos que no se encuentran ni en eucariontes ni en bacterias.
Varias otras características sitúan a Archaea en un grupo aparte. Como bacterias y eucariontes, las arqueas poseen fosfolípidos basados en glicerol,[15] pero con tres características que los hacen inusuales:[16]
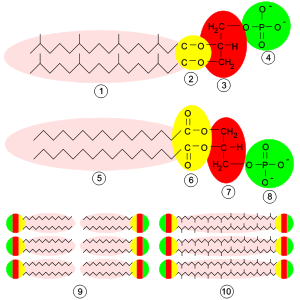 Comparación de las membranas celulares. Arriba, estructura molecular típica de los lípidos de las membranas de Archaea: 1-cadena isoprenoide, 2-enlace éter, 3-glicerol L, 4-grupo fosfato. En medio, estructura típica de los lípidos de las membranas de Bacteria y Eukarya: 5-ácido graso, 6-enlace éster, 7-glicerol D, 8-grupo fosfato. Abajo, 9-bicapa lípida característica de bacterias y eucariontes y similar a la de la mayoría de las arqueas. 10-monocapa lipídica propia de algunas arqueas.
Comparación de las membranas celulares. Arriba, estructura molecular típica de los lípidos de las membranas de Archaea: 1-cadena isoprenoide, 2-enlace éter, 3-glicerol L, 4-grupo fosfato. En medio, estructura típica de los lípidos de las membranas de Bacteria y Eukarya: 5-ácido graso, 6-enlace éster, 7-glicerol D, 8-grupo fosfato. Abajo, 9-bicapa lípida característica de bacterias y eucariontes y similar a la de la mayoría de las arqueas. 10-monocapa lipídica propia de algunas arqueas.- Estereoquímica del glicerol. Los lípidos de las arqueas son únicos porque la estereoquímica del glicerol es opuesta a la encontrada en bacterias y eucariontes. Esto evidencia fuertemente un camino biosintético independiente.
- Enlaces éter. La mayoría de bacterias y eucariontes tienen membranas compuestas principalmente por glicerol que se une a las cadenas laterales mediante enlaces éster, mientras que en las arqueas la unión es por enlaces éter. Aun cuando alguna bacteria tiene lípidos ligados por éter, la estereoquímica del glicerol sigue siendo de tipo bacteriano. Estas diferencias pueden ser debidos por una adaptación de las arqueas a los ambientes hipertermófilos. Sin embargo, hay que hacer notar que incluso las arqueas mesófilas tienen lípidos ligados por éter.
- Cadenas isoprenoides. Las cadenas laterales de las membranas celulares también tiene una composición distintiva en Archaea, pues son cadenas isoprenoides (compuestas de 20 o incluso 40 átomos de carbono), mientras que en Bacteria y Eukarya están compuestas por ácidos grasos (usualmente de 16 a 18 átomos de carbono). Las cadenas isoprenoides son comunes en la industria del caucho y como componente de algunas vitaminas comunes en bacterias y eucariontes, sin embargo, solo las arqueas incorporan estos compuestos a sus lípidos celulares. En algunas arqueas la cadena de isoprenoide C-40 es lo suficientemente larga para atravesar el espesor de la membrana, uniendo el fosfato de glicerol de ambos extremos y formando una monocapa (en lugar de la bicapa habitual). Esta adaptación hace más estable y resistente la membrana, siendo habitual, por lo tanto, en las arqueas hipertemófilas.
Aunque no son únicas, las paredes celulares de las arqueas son también inusuales. Las arqueas no tienen paredes de peptidoglicano como en las bacterias, aunque en un grupo de metanógenos contiene seudopeptidoglicano (se diferencia en la composición y en los enlaces).[17] Tampoco tienen paredes de celulosa como las plantas ni de quitina como los hongos. Las paredes celulares de otras arqueas carecen tanto de peptidoglicano como de seudopeptidoglicano y se componen de polisacáridos, glicoproteínas o proteínas. El tipo más común de pared es la capa superficial paracristalina (capa S) que está formada por proteína o glicoproteína, generalmente de simetría hexagonal.[18] Las capas S son comunes en las bacterias, donde constituyen el único componente de la pared celular en algunos organismos (como en Planctomyces) o la capa externa de muchos otros organismos con peptidoglicano. La arquea Thermoplasma carece de pared celular.
Las arqueas también tienen flagelos que son notablemente diferentes en composición y desarrollo de los superficialmente similares flagelos de las bacterias.[19] El flagelo bacteriano es un sistema de secreción de tipo III modificado, mientras que los flagelos de las arqueas se asemejan a los pilis de tipo IV, los cuales utilizan un sistema de secreción algo similar pero no idéntico al sistema de secreción de tipo II.[20]
Morfología
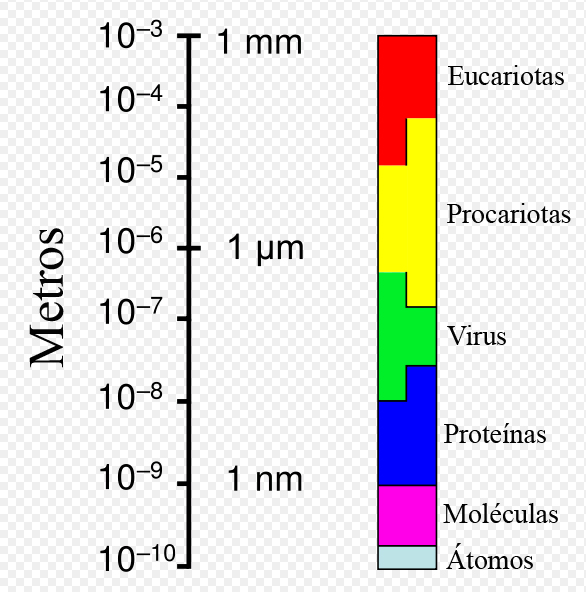
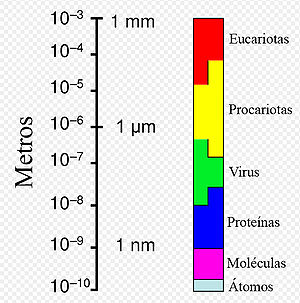 Rango de tamaños que presentan las células procariotas en relación a otros organismos y biomoléculas.
Rango de tamaños que presentan las células procariotas en relación a otros organismos y biomoléculas.Las arqueas individuales tienen diámetros comprendidos entre 0,1 μm y más de 15 μm y se presentan en diversas formas, siendo comunes esferas, barras, espirales y placas.[10] El grupo Crenarchaeota incluye otras morfologías tales como células lobuladas irregularmente en Sulfolobus, finos filamentos de menos de medio μm de diámetro en Thermofilum y barras casi perfectamente rectangulares en Thermoproteus y Pyrobaculum.[21] Recientemente, se ha descubierto en piscinas hipersalinas una especie de forma cuadrada y plana (como un sello de correos) denominada Haloquadra walsbyi.[22] Estas formas inusuales probablemente se mantienen tanto por la pared celular como por un citoesqueleto procariótico, pero estas estructuras celulares, en contraste con las bacterianas, son poco conocidas.[23] En las células de las arqueas se han identificado, sin embargo, proteínas relacionadas con los componentes del citoesqueleto,[24] así como filamentos.[25]
Algunas especies forman agregados o filamentos celulares de hasta 200 μm de longitud[10] y pueden ser miembros prominentes de las comunidades de microbios que conforman biopelículas.[26] Un ejemplo particularmente elaborado de colonias multicelulares lo constituyen las arqueas del género Pyrodictium. En este caso, las células producen arreglos de tubos largos, delgados y huecos, denominados cánulas, que se conectan formando densas colonias arbustivas.[27] La función de estas cánulas se desconoce, pero pueden permitir que las células se comuniquen o intercambien nutrientes con sus vecinos.[28]
Genética
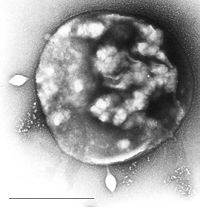
Las arqueas por lo general tienen un único cromosoma circular al igual que las bacterias.[30] Estos varían en tamaño desde 5.751.492 pares de bases en Methanosarcina acetivorans,[31] el mayor genoma secuenciado hasta la fecha, hasta 490.885 pares de bases en Nanoarchaeum equitans, el genoma más pequeño conocido que puede contener sólo 537 genes codificadores de proteínas. Las arqueas también presentan plásmidos[32] que se pueden propagar por contacto físico entre células, en un proceso que puede ser similar a la conjugación bacteriana.[33] [34] Los plásmidos son cada vez más importantes como herramientas genéticas pues permiten la realización de estudios genéticos en Archaea.[35]
Al igual que los bacteriófagos que infectan bacterias, existen virus que se replican en las arqueas. Son virus ADN bicatenarios que parecen no estar relacionados con otros virus y que pueden tener una gran variedad de formas inusuales, como botellas o ganchos.[36] Estos virus han sido estudiados en más detalle en los termófilos, en particular en los órdenes Sulfolobales y Thermoproteales.[37] La defensa contra estos virus pueden implicar la interferencia de ARN por secuencias de ADN repetitivas en el genoma de las arqueas.[38] [39]
Las arqueas son genéticamente distintas a otros organismos, con hasta un 15% de proteínas exclusivas codificadas por el genoma de cualquiera arquea.[40] Los genes que son compartidos entre Archaea, Bacteria y Eukarya forman un núcleo de funciones de la célula, relacionados principalmente con la transcripción, traducción y metabolismo de nucleótidos.[41] La mayoría de los genes exclusivos de las arqueas no tienen una función conocida, pero de los que tienen una función identificada, la mayoría participan en la metanogénesis. Otros elementos característicos de los genomas de las arqueas son la organización de genes de función relacionada, tales como las enzimas que catalizan las etapas de la misma ruta metabólica, nuevos operones y grandes diferencias en genes ARNt y sus aminoacil ARNt sintetasas.[41]
La transcripción y traducción en Archaea son más similares a Eukarya que a Bacteria, por ejemplo en las subunidades y secuencias de la ARN polimerasa II y en los ribosomas.[30] Las funciones e interacciones de la ARN polimerasa en la transcripción en Archaea también parece estar relacionada con la de Eukarya, con un similar ensamblado de proteínas (factores de transcripción genéricos) dirigiendo la unión de la ARN polimerasa a un promotor de gen. Sin embargo, muchos otros factores de transcripción en las arqueas son similares a los de bacterias.[42]
Reproducción
Las arqueas se reproducen asexualmente por fisión binaria o múltiple, fragmentación o gemación. No se produce meiosis, de manera que si una especie de arquea existe en más de una forma, todas tienen el mismo número de cromosomas (tienen el mismo cariotipo).[10] La división celular está controlada como parte de un complejo ciclo celular, donde el cromosoma se replica, las copias se separan y luego la célula se divide.[43] Los detalles del ciclo celular sólo han sido investigados en el género Sulfolobus, siendo similares a los de bacterias y eucariontes: los cromosomas se replican desde múltiples puntos de partida (origen de replicación) usando ADN polimerasas que son similares a las enzimas equivalentes eucarióticas.[44] Sin embargo, las proteínas que dirigen la división celular, como la proteína FtsZ que forma un anillo contráctil alrededor de la célula, parecen estar más relacionadas con sus equivalentes bacterianos.[43]
No se forman endosporas en ninguna especie de arquea,[45] aunque algunas especies de Halobacteria pueden alternar entre fenotipos y crecer como diferentes tipos de células, incluidas estructuras de paredes gruesas que son resistentes al choque osmótico y que les permiten sobrevivir a bajas concentraciones de sal.[46] No se trata de estructuras reproductivas, pero es posible que ayuden a estas especies a dispersarse en nuevos hábitats.
Origen y evolución
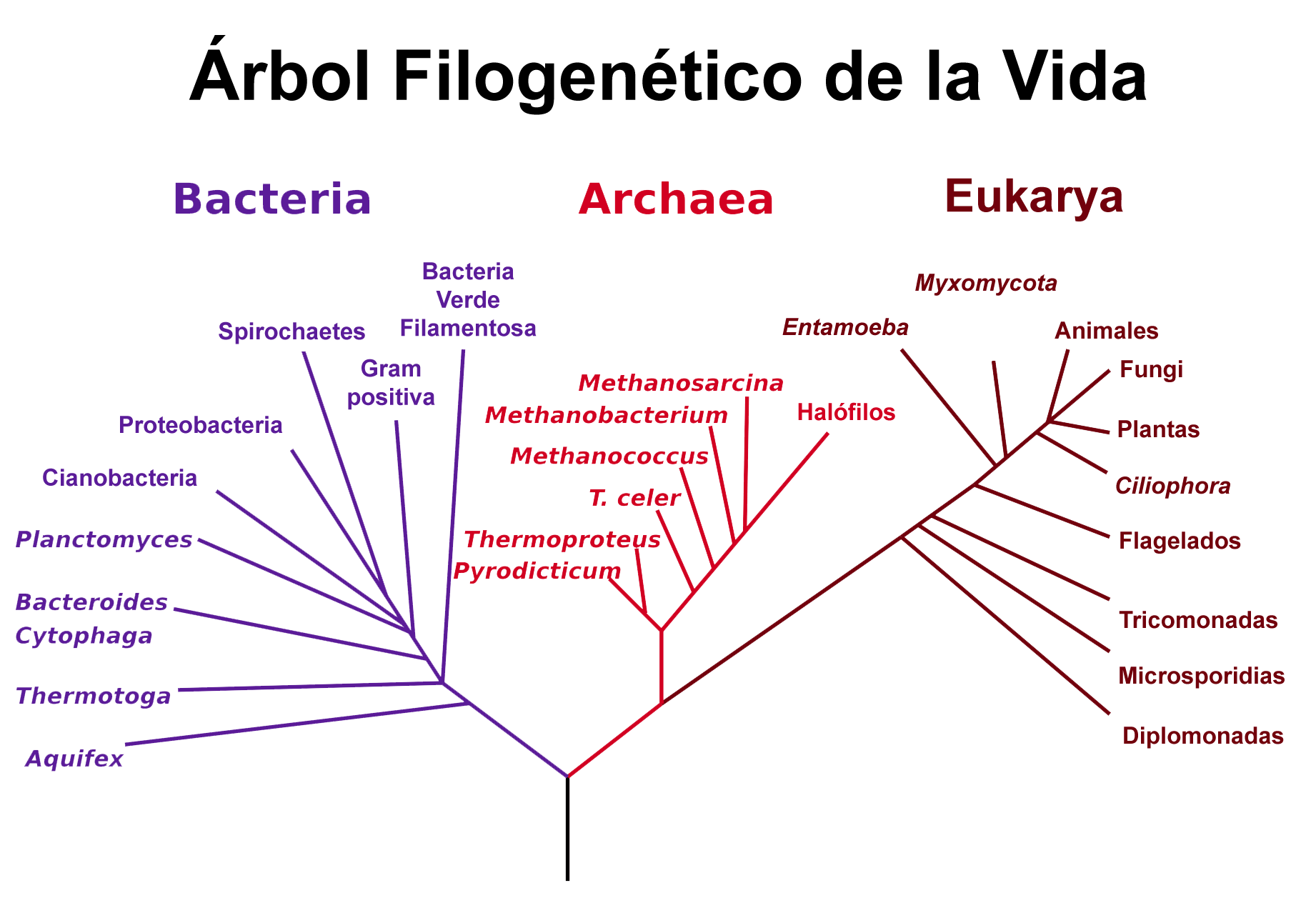
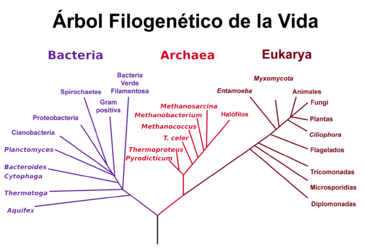 Situación de las arqueas en el árbol filogénético de Carl Woese et al basado en datos de secuencias genéticas de ARNr 16S.
Situación de las arqueas en el árbol filogénético de Carl Woese et al basado en datos de secuencias genéticas de ARNr 16S.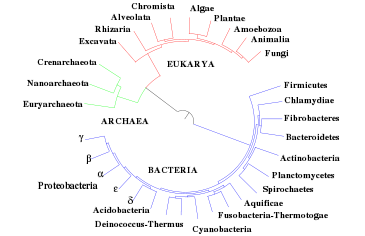 Árbol filogenético de los seres vivos obtenido a partir de genomas completamente secuenciados.[47] Los árboles moleculares colocan a Archaea más próximo a Eukarya que a Bacteria.
Árbol filogenético de los seres vivos obtenido a partir de genomas completamente secuenciados.[47] Los árboles moleculares colocan a Archaea más próximo a Eukarya que a Bacteria.No hay acuerdo sobre cuándo se han originado las arqueas puesto que los fósiles más tempranos son difíciles de interpretar. Los sedimentos más antiguos que se conocen, los de Groenlandia occidental de hace 3.800 millones de años, podrían contener restos de arqueas.[48] [49] Fósiles probables datan de hace casi 3.500 millones de años[50] y restos de lípidos, que podrían ser tanto de arqueas como de eucariontes, se han detectado en pizarras de hace 2.700 millones de años.[51] Sin embargo, no todos los investigadores están de acuerdo con la asignación de estos fósiles a Archaea y sitúan su origen hace sólo 900 millones de años.[52] [53]
Las arqueas fueron identificadas como una línea evolutiva separada en 1977 (y finalmente promovidas a dominio en 1990) por Carl Woese y George E. Fox, debido a su posición en los árboles filogenéticos basados en secuencias de ARNr.[7] Aunque hay cierta incertidumbre en la posición filogenética de los dominios Archaea, Bacteria y Eukarya se considera que, a pesar de ser procariontes, las arqueas están más relacionadas con los eucariontes que con las bacterias.
Hay dos puntos de vista fundamentalmente diferentes sobre el origen de Archaea. Una de las posibilidades es que la especificidad de Archaea refleje una temprana divergencia en el árbol de la vida, antes de que muchas de las características celulares se estabilizaran. En este sentido, algunos estudios sugieren que puede ser el linaje más antiguo, con Bacteria y Eukarya divergiendo de este grupo.[54] Puesto que un estudio de árboles moleculares establece que el último ancestro común de las arqueas existentes en la actualidad pudiera ser un termófilo no metanógeno,[55] se plantea la posibilidad de que los organismos que prosperan a temperaturas templadas aparecieran más tarde que los adaptados a temperaturas altas. Por su parte, Woese sugiere que los tres dominios podrían ser igual de antiguos pues derivarían de una colonia ancestral en la que la transferencia horizontal de genes fuese el principal motor evolutivo hasta que la transferencia vertical se hizo dominante y los dominios se separaran.[56]
El otro punto de vista, mayoritario en la actualidad, es que Archaea es relativamente reciente y que surgió de bacterias especializadas.[57] En esta línea, Cavalier-Smith sostiene que las innovaciones que dieron lugar a las arqueas tuvieron como partida un antecesor Neomura, del cual se derivaron tanto Archaea como Eukarya, que serían, por tanto, grupos hermanos.[58] [59] [60] Neomura, a su vez, puede haberse originado a partir de actinobacterias (bacterias gram positivas) que perdieron su pared externa de mureína. Luego una posterior adaptación a condiciones hipertermófilas da origen a Archaea, que sería tan reciente como Eukarya y de hace sólo unos 900 millones de años.[52] [53]
La relación entre Archaea, Bacteria y Eukarya sigue siendo todavía un problema importante.
Clasificación
La sistemática de las arqueas no está completa y las diferencias entre algunos grupos son muy grandes.
Se conocen unos 100 géneros y unas 300 especies de arqueas. Se proponen cuatro filos (cada uno de los cuales se puede incluir en un reino del mismo nombre), comprendiendo los dos primeros la mayor parte de las especies conocidas:
- Crenarchaeota. Comprende a la mayoría de las especies hipertermófilas. Muchas son autótrofas quimiosintéticas, constituyendo los únicos productores primarios en ciertos hábitats inhóspitos. Recientes análisis basados en secuencias de ARNr indican que Crenarchaeota también se puede distribuir extensamente en ambientes de baja temperatura tales como mares, sedimentos y suelos.
- Euryarchaeota. Comprende a las que se encuentran en concentraciones salinas altas y que pueden obtener su energía de la luz, sin ningún pigmento clorofílico. También se clasifican dentro de este clado los metanógenos que habitan los sedimentos acuosos y en el tracto digestivo de diversos animales. Análisis de secuencias de ARNr sugieren que miembros de este grupo también se encuentran en gran número en las aguas oceánicas.
- Korarchaeota. Es un pequeño grupo de hipertermófilas (encontradas en el parque Yellowstone), filogenéticamente diferentes a las Crenarchaeotas y que se consideran las arqueas más primitivas.
- Nanoarchaeota. Es un taxón especialmente creado para la especie Nanoarchaeum equitans, descubierta en 2002 y cuyas relaciones de parentesco son inciertas.[61]
Se debe tomar en cuenta que, aunque los dos primeros filos están firmemente establecidos, los dos restantes comprenden pocas especies mal conocidas y son dudosos. Dado el escaso conocimiento actual sobre las especies de este dominio, el número de filos puede aumentar con rapidez. Muchos de estos hipotéticos grupos son conocidos por una única secuencia de ARNr, lo que indica que la gran mayoría de la diversidad de estos organismos sigue siendo completamente desconocida.[62] [63] Este problema de como abordar el estudio y clasificación de microbios no cultivados es común en todos los procariontes.[64] Recientemente (en 2006) se han descubierto tres nuevos grupos de Euryarchaeota en el ambiente extremadamente ácido en una mina del Norte de California, denominados provisionalmente ARMAN (Archaeal Richmond Mine Acidophilic Nanoorganisms).[65]
Uso de Archaea en tecnología e industria
Las arqueas extremófilas, en particular las resistentes a las altas temperaturas o a los extremos de acidez y alcalinidad, son una importante fuente de enzimas que puede funcionar bajo estas duras condiciones.[66] [67] Estas enzimas tienen una amplia gama de usos. Por ejemplo, las ADN polimerasas termoestables, como la ADN polimerasa Pfu de Pyrococcus furiosus, han revolucionado la biología molecular, al permitir el uso de la reacción en cadena de la polimerasa como método simple y rápido para la clonación del ADN.
En la industria, las amilasas, galactosidasas y pululanasas de otras especies de Pyrococcus realizan su función a más de 100 °C, lo que permite la elaboración de alimentos a altas temperaturas, tales como leche baja en lactosa y suero de leche.[68] Las enzimas de estas arqueas termófilas también tienden a ser muy estables en solventes orgánicos, por lo que pueden utilizarse en una amplia gama de procesos respetuosos con el medio ambiente para la síntesis de compuestos orgánicos.[67]
En contraste con la amplia gama de aplicaciones de las enzimas, la utilización en biotecnología de los organismos en sí mismos es más reducida. Sin embargo, las arqueas metanógeneas son una parte vital del tratamiento de aguas residuales, realizando la digestión anaeróbica de los residuos y produciendo biogás.[69] Las arqueas acidófilas son también prometedores en minería para la extracción de metales tales como oro, cobalto y cobre.[70]
Una nueva clase de antibióticos potencialmente útiles se derivan de este grupo de organismos. Ocho de esas sustancias ya han sido caracterizadas, pero podría haber muchas más, especialmente en Halobacteria. El descubrimiento de nuevas sustancias depende de la recuperación de estos organismos del medio ambiente y de su cultivo.[71]
Referencias
- ↑ DeLong EF, Pace NR (2001). «Environmental diversity of bacteria and archaea» Syst. Biol.. Vol. 50. n.º 4. pp. 470-8. PMID 12116647.
- ↑ Karner MB, DeLong EF, Karl DM (2001). «Archaeal dominance in the mesopelagic zone of the Pacific Ocean» Nature. Vol. 409. n.º 6819. pp. 507-10. PMID 11206545.
- ↑ a b c Valentine DL (2007). «Adaptations to energy stress dictate the ecology and evolution of the Archaea» Nat. Rev. Microbiol.. Vol. 5. n.º 4. pp. 316–23. PMID 17334387.
- ↑ Eckburg P, Lepp P, Relman D (2003). «Archaea and their potential role in human disease» Infect Immun. Vol. 71. n.º 2. pp. 591-6. PMID 12540534.
- ↑ Cavicchioli R, Curmi P, Saunders N, Thomas T (2003). «Pathogenic archaea: do they exist?» Bioessays. Vol. 25. n.º 11. pp. 1119-28. PMID 14579252.
- ↑ Lepp P, Brinig M, Ouverney C, Palm K, Armitage G, Relman D (2004). «Methanogenic Archaea and human periodontal disease» Proc Natl Acad Sci U S A. Vol. 101. n.º 16. pp. 6176-81. PMID 15067114.
- ↑ a b Woese C, Fox G (1977). «Phylogenetic structure of the prokaryotic domain: the primary kingdoms» Proc Natl Acad Sci U S A. Vol. 74. n.º 11. pp. 5088 – 90. PMID 270744.
- ↑ Woese, Carl R., Kandler, Otto, Wheelis, Mark L (1990). «Towards a natural system of organisms: Proposal for the domains Archaea, Bacteria, and Eucarya» Proceedings of the National Academy of Sciences. Vol. 87. n.º 12. pp. 4576 – 4579.
- ↑ Burns DG, Camakaris HM, Janssen PH, Dyall-Smith ML. (2004). «Cultivation of Walsby's square haloarchaeon.» FEMS Microbiol Lett.. Vol. 238. n.º 2. pp. 469-73. PMID 15358434.
- ↑ a b c d Krieg, Noel (2005). Bergey’s Manual® of Systematic Bacteriology. USA: Springer, pp. 21-26. ISBN 978-0-387-24143-2.
- ↑ a b Eckburg PB, Bik EM, Bernstein CN, et al (2005). «Diversity of the human intestinal microbial flora» Science. Vol. 308. n.º 5728. pp. 1635–8. PMID 15831718.
- ↑ H.C. Tung, N.E. Bramall, and P.B. Price (2005), Microbial origin of excess methane in glacial ice and implications for life on Mars, Proceedings of the National Academy of Sciences (DOI: 10.1073/pnas.0507601102)
- ↑ Giovannoni SJ, Stingl U. (2005). «Molecular diversity and ecology of microbial plankton» Nature. Vol. 427. n.º 7057. pp. 343-8. PMID 16163344.
- ↑ Konneke M, Bernhard AE, de la Torre JR, Walker CB, Waterbury JB, Stahl DA. (2005). «Isolation of an autotrophic ammonia-oxidizing marine archaeon» Nature. Vol. 437. n.º 7057. pp. 543-6. PMID 16177789.
- ↑ De Rosa M, Gambacorta A, Gliozzi A (1986). «Structure, biosynthesis, and physicochemical properties of archaebacterial lipids» Microbiol. Rev.. Vol. 50. n.º 1. pp. 70–80. PMID 3083222.
- ↑ Koga Y, Morii H (2007). «Biosynthesis of ether-type polar lipids in archaea and evolutionary considerations» Microbiol. Mol. Biol. Rev.. Vol. 71. n.º 1. pp. 97–120. DOI 10.1128/MMBR.00033-06. PMID 17347520.
- ↑ Howland, John L. (2000) The Surprising Archaea: Discovering Another domain of Life, pages 69-71. (Oxford: Oxford University Press). ISBN 0-19-511183-4.
- ↑ Sára M, Sleytr UB (2000). «S-Layer proteins» J. Bacteriol.. Vol. 182. n.º 4. pp. 859–68. PMID 10648507.
- ↑ Thomas NA, Bardy SL, Jarrell KF (2001). «The archaeal flagellum: a different kind of prokaryotic motility structure» FEMS Microbiol. Rev.. Vol. 25. n.º 2. pp. 147–74. PMID 11250034.
- ↑ Ng SY, Chaban B, Jarrell KF (2006). «Archaeal flagella, bacterial flagella and type IV pili: a comparison of genes and posttranslational modifications» J. Mol. Microbiol. Biotechnol.. Vol. 11. n.º 3-5. pp. 167–91. PMID 16983194.
- ↑ Barns, Sue and Burggraf, Siegfried. (1997) Crenarchaeota. Version 01 January 1997. in The Tree of Life Web Project
- ↑ Walsby AE (2005). «Archaea with square cells» Trends Microbiol.. Vol. 13. n.º 5. pp. 193–5. PMID 15866034.
- ↑ Hixon WG, Searcy DG (1993). «Cytoskeleton in the archaebacterium Thermoplasma acidophilum? Viscosity increase in soluble extracts» BioSystems. Vol. 29. n.º 2-3. pp. 151–60. PMID 8374067.
- ↑ Hara F, Yamashiro K, Nemoto N, et al (2007). «An actin homolog of the archaeon Thermoplasma acidophilum that retains the ancient characteristics of eukaryotic actin» J. Bacteriol.. Vol. 189. n.º 5. pp. 2039–45. DOI 10.1128/JB.01454-06. PMID 17189356.
- ↑ Trent JD, Kagawa HK, Yaoi T, Olle E, Zaluzec NJ (1997). «Chaperonin filaments: the archaeal cytoskeleton?» Proc. Natl. Acad. Sci. U.S.A.. Vol. 94. n.º 10. pp. 5383–8. PMID 9144246.
- ↑ Hall-Stoodley L, Costerton JW, Stoodley P (2004). «Bacterial biofilms: from the natural environment to infectious diseases» Nat. Rev. Microbiol.. Vol. 2. n.º 2. pp. 95–108. PMID 15040259.
- ↑ Nickell S, Hegerl R, Baumeister W, Rachel R (2003). «Pyrodictium cannulae enter the periplasmic space but do not enter the cytoplasm, as revealed by cryo-electron tomography» J. Struct. Biol.. Vol. 141. n.º 1. pp. 34–42. PMID 12576018.
- ↑ Horn C, Paulmann B, Kerlen G, Junker N, Huber H (1999). «In vivo observation of cell division of anaerobic hyperthermophiles by using a high-intensity dark-field microscope» J. Bacteriol.. Vol. 181. n.º 16. pp. 5114–8. DOI 10.1073/pnas.241636498v1. PMID 10438790.
- ↑ Xiang X, Chen L, Huang X, Luo Y, She Q, Huang L (2005). «Sulfolobus tengchongensis spindle-shaped virus STSV1: virus-host interactions and genomic features» J. Virol.. Vol. 79. n.º 14. pp. 8677–86. DOI 10.1128/JVI.79.14.8677-8686.2005. PMID 15994761.
- ↑ a b Allers T, Mevarech M (2005). «Archaeal genetics - the third way» Nat. Rev. Genet.. Vol. 6. n.º 1. pp. 58–73. PMID 15630422.
- ↑ Baliga NS, Bonneau R, Facciotti MT, et al (2004). «Genome sequence of Haloarcula marismortui: a halophilic archaeon from the Dead Sea» Genome Res.. Vol. 14. n.º 11. pp. 2221–34. DOI 10.1101/gr.2700304. PMID 15520287.
- ↑ Waters E, Hohn MJ, Ahel I, et al (2003). «The genome of Nanoarchaeum equitans: insights into early archaeal evolution and derived parasitism» Proc. Natl. Acad. Sci. U.S.A.. Vol. 100. n.º 22. pp. 12984–8. DOI 10.1073/pnas.1735403100. PMID 14566062.
- ↑ Schleper C, Holz I, Janekovic D, Murphy J, Zillig W (1995). «A multicopy plasmid of the extremely thermophilic archaeon Sulfolobus effects its transfer to recipients by mating» J. Bacteriol.. Vol. 177. n.º 15. pp. 4417–26. PMID 7635827.
- ↑ Sota M; Top EM (2008). «Horizontal Gene Transfer Mediated by Plasmids», Plasmids: Current Research and Future Trends. Caister Academic Press. ISBN 978-1-904455-35-6.
- ↑ Lipps G (2008). «Archaeal Plasmids», Plasmids: Current Research and Future Trends. Caister Academic Press. ISBN 978-1-904455-35-6.
- ↑ Prangishvili D, Forterre P, Garrett RA (2006). «Viruses of the Archaea: a unifying view» Nat. Rev. Microbiol.. Vol. 4. n.º 11. pp. 837–48. PMID 17041631.
- ↑ Prangishvili D, Garrett RA (2004). «Exceptionally diverse morphotypes and genomes of crenarchaeal hyperthermophilic viruses» Biochem. Soc. Trans.. Vol. 32. n.º Pt 2. pp. 204–8. PMID 15046572.
- ↑ Mojica FJ, Díez-Villaseñor C, García-Martínez J, Soria E (2005). «Intervening sequences of regularly spaced prokaryotic repeats derive from foreign genetic elements» J. Mol. Evol.. Vol. 60. n.º 2. pp. 174–82. PMID 15791728.
- ↑ Makarova KS, Grishin NV, Shabalina SA, Wolf YI, Koonin EV (2006). «A putative RNA-interference-based immune system in prokaryotes: computational analysis of the predicted enzymatic machinery, functional analogies with eukaryotic RNAi, and hypothetical mechanisms of action» Biol. Direct. Vol. 1. pp. 7. PMID 16545108.
- ↑ Graham DE, Overbeek R, Olsen GJ, Woese CR (2000). «An archaeal genomic signature» Proc. Natl. Acad. Sci. U.S.A.. Vol. 97. n.º 7. pp. 3304–8. DOI 10.1073/pnas.050564797. PMID 10716711.
- ↑ a b Gaasterland T (1999). «Archaeal genomics» Curr. Opin. Microbiol.. Vol. 2. n.º 5. pp. 542–7. PMID 10508726.
- ↑ Aravind L, Koonin EV (1999). «DNA-binding proteins and evolution of transcription regulation in the archaea» Nucleic Acids Res.. Vol. 27. n.º 23. pp. 4658–70. PMID 10556324.
- ↑ a b Bernander R (1998). «Archaea and the cell cycle» Mol. Microbiol.. Vol. 29. n.º 4. pp. 955–61. PMID 9767564.
- ↑ Kelman LM, Kelman Z (2004). «Multiple origins of replication in archaea» Trends Microbiol.. Vol. 12. n.º 9. pp. 399–401. PMID 153371581.
- ↑ Onyenwoke RU, Brill JA, Farahi K, Wiegel J (2004). «Sporulation genes in members of the low G+C Gram-type-positive phylogenetic branch ( Firmicutes)» Arch. Microbiol.. Vol. 182. n.º 2-3. pp. 182–92. PMID 15340788.
- ↑ Kostrikina NA, Zvyagintseva IS, Duda VI. (1991). «Cytological peculiarities of some extremely halophilic soil archaeobacteria» Arch. Microbiol.. Vol. 156. n.º 5. pp. 344-49. DOI 10.1007/BF00248708.
- ↑ Ciccarelli FD, Doerks T, von Mering C, Creevey CJ, Snel B, Bork P (2006). «Toward automatic reconstruction of a highly resolved tree of life» Science. Vol. 311. n.º 5765. pp. 1283-7. PMID 165139821.
- ↑ Hahn J, Haug P (1986). «Traces of Archaebacteria in ancient sediments» System Appl Microbiol. Vol. 7. pp. 178-183..
- ↑ Chappe, B. and Albrecht, P. and Michaelis, W. ({1982). «Polar Lipids of Archaebacteria in Sediments and Petroleums» Science. Vol. 217. n.º 4554. pp. 65-66. DOI 10.1126/science.217.4554.65.
- ↑ Schopf J (2006). «Fossil evidence of Archaean life» Philos Trans R Soc Lond B Biol Sci. Vol. 361. n.º 1470. pp. 869-85. PMID 16754604.
- ↑ Brocks JJ, Logan GA, Buick R, Summons RE (1999). «Archean molecular fossils and the early rise of eukaryotes» Science. Vol. 285. n.º 5430. pp. 1033-6. PMID 10446042.
- ↑ a b Thomas Cavalier-Smith (2006), Rooting the tree of life by transition analyses, Biol Direct. 1: 19.
- ↑ a b Cavalier-Smith T (2006). «Cell evolution and Earth history: stasis and revolution» Philos Trans R Soc Lond B Biol Sci. Vol. 361. n.º 1470. pp. 969–1006. PMID 16754610.
- ↑ Wang M, Yafremava LS, Caetano-Anollés D, Mittenthal JE, Caetano-Anollés G (2007). «Reductive evolution of architectural repertoires in proteomes and the birth of the tripartite world» Genome Res.. Vol. 17. n.º 11. pp. 1572–85. DOI 10.1101/gr.6454307. PMID 17908824.
- ↑ Gribaldo S, Brochier-Armanet C (2006). «The origin and evolution of Archaea: a state of the art» Philos. Trans. R. Soc. Lond., B, Biol. Sci.. Vol. 361. n.º 1470. pp. 1007-22. DOI 10.1098/rstb.2006.1841. PMID 16754611.
- ↑ Woese C (1998). «[http://www.pnas.org/cgi/content/full/95/12/6854 The universal ancestor]» Proc. Natl. Acad. Sci. U.S.A.. Vol. 95. n.º 12. pp. 6854-9. PMID 9618502.
- ↑ Gupta RS (2000). «The natural evolutionary relationships among prokaryotes» Crit. Rev. Microbiol.. Vol. 26. n.º 2. pp. 111–31. PMID 10890353.
- ↑ Thomas Cavalier-Smith (1987), The origin of eukaryote and archaebacterial cells, Ann NY Acad Sci 503: 17–54
- ↑ Thomas Cavalier-Smith (2002), The neomuran origin of archaebacteria, the negibacterial root of the universal tree and bacterial megaclassification, International Journal of Systematic and Evolutionary Microbiology 52: 7–76
- ↑ T. Cavalier-Smith (2002). «The phagotrophic origin of eukaryotes and phylogenetic classification of Protozoa» International Journal of Systematic and Evolutionary Microbiology. Vol. 52. pp. 297-354.
- ↑ Huber H, Hohn MJ, Rachel R, Fuchs T, Wimmer VC, Stetter KO. (2002). «A new phylum of Archaea represented by a nanosized hyperthermophilic symbiont.» Nature. Vol. 417. n.º 6884. pp. 27 – 8. PMID 11986665.
- ↑ Robertson CE, Harris JK, Spear JR, Pace NR (2005). «Phylogenetic diversity and ecology of environmental Archaea» Curr. Opin. Microbiol.. Vol. 8. n.º 6. pp. 638–42. PMID 16236543.
- ↑ Hugenholtz P (2002). «Exploring prokaryotic diversity in the genomic era» Genome Biol.. Vol. 3. n.º 2. pp. REVIEWS0003. DOI 10.1186/gb-2002-3-2-reviews0003. PMID 11864374.
- ↑ Rappé MS, Giovannoni SJ (2003). «The uncultured microbial majority» Annu. Rev. Microbiol.. Vol. 57. pp. 369–94. PMID 14527284.
- ↑ Baker, B.J., Tyson, G.W., Webb, R.I., Flanagan, J., Hugenholtz, P. and Banfield, J.F. (2006). «Lineages of acidophilic Archaea revealed by community genomic analysis. Science» Science. Vol. 314. n.º 6884. pp. 1933 – 1935.
- ↑ Breithaupt H (2001). «The hunt for living gold. The search for organisms in extreme environments yields useful enzymes for industry» EMBO Rep.. Vol. 2. n.º 11. pp. 968–71. DOI 10.1093/embo-reports/kve238. PMID 11713183.
- ↑ a b Egorova K, Antranikian G (2005). «Industrial relevance of thermophilic Archaea» Curr. Opin. Microbiol.. Vol. 8. n.º 6. pp. 649–55. PMID 16257257.
- ↑ Synowiecki J, Grzybowska B, Zdziebło A (2006). «Sources, properties and suitability of new thermostable enzymes in food processing» Crit Rev Food Sci Nutr. Vol. 46. n.º 3. pp. 197–205. PMID 16527752.
- ↑ Schiraldi C, Giuliano M, De Rosa M (2002). «Perspectives on biotechnological applications of archaea» Archaea. Vol. 1. n.º 2. pp. 75–86. PMID 15803645.
- ↑ Norris PR, Burton NP, Foulis NA (2000). «Acidophiles in bioreactor mineral processing» Extremophiles. Vol. 4. n.º 2. pp. 71–6. PMID 10805560.
- ↑ Shand RF; Leyva KJ (2008). «Archaeal Antimicrobials: An Undiscovered Country», Archaea: New Models for Prokaryotic Biology. Caister Academic Press. ISBN 978-1-904455-27-1.
Enlaces externos
 Wikiespecies tiene un artículo sobre Archaea.
Wikiespecies tiene un artículo sobre Archaea.- Molecular phylogeny of Archaea in boreal forest soil, freshwater and temperate estuarine sediment
- Oceans of Archaea
Categorías: Archaea | Extremófilos




Wikimedia foundation. 2010.