- Evolución del sexo
-
Evolución del sexo

La evolución del sexo es un gran rompecabezas de la biología evolutiva moderna. Muchos grupos de organismos, en especial la mayoría de los animales y las plantas, se reproducen sexualmente. La evolución del sexo contiene dos temas relacionados, aunque diferentes: su origen y su mantenimiento. Sin embargo, como las hipótesis para el origen del sexo son difíciles de comprobar experimentalmente, la mayor parte del trabajo actual se ha centrado en el mantenimiento de la reproducción sexual. Los biólogos, incluyendo a W. D. Hamilton, Alexei Kondrashov y George C. Williams, han propuesto varias explicaciones de cómo se mantiene la reproducción sexual en un gran conjunto de seres vivos distintos.
Parece que el ciclo sexual se mantiene porque mejora la calidad de la progenie (aptitud), a pesar de reducir el número total de la descendencia (el doble coste del sexo). Para que sea evolutivamente ventajoso debe estar asociado con un aumento significativo de la aptitud de la descendencia. Una de las explicaciones más aceptadas para la ventaja del sexo recae en la creación de variabilidad genética. Hay tres razones posibles que pueden dar cuenta de esto. Primero, la reproducción sexual puede juntar mutaciones que son beneficiosas en el mismo individuo (el sexo ayuda en la difusión de caracteres ventajosos). Segundo, el sexo actúa juntando mutaciones perjudiciales para crear individuos con una aptitud muy baja que son eliminados de la población (el sexo ayuda a eliminar los genes perjudiciales). Por último, el sexo crea nuevas combinaciones de genes que pueden ser más aptas que las existentes anteriormente, o que simplemente producen una menor competitividad entre los parientes.
Estas clases de hipótesis se explican con mayor detalle más adelante. Es importante tener en cuenta que cualquier número de estas hipótesis puede ser cierta en una especie concreta (no son eventos mutuamente exclusivos), y que en otras especies pueden aplicarse otras hipótesis. Sin embargo, todavía tiene que encontrarse un marco de investigación que permita determinar si la razón para el sexo es universal para todas las especies y, si no lo es, qué mecanismo está actuando en cada especie.
Historia inicial del estudio
El pensamiento filosófico científico sobre el problema se puede remontar hasta Erasmus Darwin en el siglo XVIII; también aparece en los escritos de Aristóteles. El tema lo retomó en el siglo XIX August Weismann, que comprendió que el propósito del sexo era generar variabilidad genética, tal y como se detalla abajo.
El doble coste del sexo
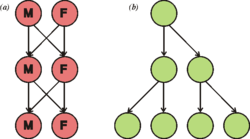 Este diagrama ilustra el doble coste del sexo. Si cada individuo contribuyera con el mismo número de descendientes (dos), la población sexual (a) mantiene su tamaño tras cada generación, mientras que la población asexual (b) dobla su tamaño en cada generación.
Este diagrama ilustra el doble coste del sexo. Si cada individuo contribuyera con el mismo número de descendientes (dos), la población sexual (a) mantiene su tamaño tras cada generación, mientras que la población asexual (b) dobla su tamaño en cada generación.
En todas las especies sexuales, la población consiste en dos sexos, de los cuales solo uno de ellos es capaz de engendrar la prole (con la excepción de los hermafroditas simultáneos). En una especie asexual, todos los miembros de la población son capaces de engendrar descendencia. Esto implica, de manera intrínseca, que, en cada generación, una población asexual puede crecer más rápidamente. Este coste fue definido por primera vez en términos matemáticos por John Maynard Smith.
Un coste adicional es que los machos y las hembras deben buscarse entre ellos para aparearse, y la selección sexual suele favorecer caracteres que reducen la aptitud de los individuos.[1]
Sin embargo, George C. Williams aportó evidencia de que este coste no es insuperable, apuntando la existencia de especies que son capaces de reproducirse tanto asexual como sexualmente. Estas especies programan su reproducción sexual para los periodos de incertidumbre ambiental, y se reproducen de manera asexual cuando las condiciones son más favorables. El punto importante es que se observa que estas especies se reproducen sexualmente cuando podrían elegir no hacerlo, lo que implica que existe una ventaja selectiva en la reproducción sexual.[2]
El sexo crea variabilidad genética entre los hermanos
August Weismann propuso en 1889[3] una explicación para la evolución del sexo en la que la ventaja de este es la creación de variabilidad entre hermanos. Luego fue explicada en términos genéticos por Fisher[4] y Muller[5] y recientemente ha sido resumida por Burt, en 2000.[6]
George C. Williams proporcionó un ejemplo basado en el olmo. En el bosque de este ejemplo, las zonas vacías que había entre los árboles se consideraba que podían sustentar a un individuo cada una. Cuando una zona queda disponible por la muerte de un árbol, habrá una competencia para rellenar esa zona. Como la probabilidad de que una semilla consiga ocupar esa zona depende de su genotipo, y un progenitor no es capaz de anticipar qué genotipo será el más exitoso, los progenitores lanzarán muchas semillas, creando una competencia entre hermanos. La selección natural, por tanto, favorece a los progenitores que pueden producir una gran variedad de descendencia. Una hipótesis similar se conoce como hipótesis del ribazo enmarañado por un pasaje de El origen de las especies, de Charles Darwin:
"Es interesante contemplar un enmarañado ribazo cubierto por muchas plantas de varias clases, con aves que cantan en los arbustos, con diferentes insectos que revolotean y con gusanos que se arrastran entre la tierra húmeda y reflexionar que estas formas, primorosamente construidas, tan diferentes entre sí, y que dependen mutuamente de modos tan complejos, han sido producidas por leyes que obran a nuestro alrededor."
Esta hipótesis, propuesta por Michael Ghiselin en su libro The Economy of Nature and the Evolution of Sex, publicado en 1974, sugiere que un grupo diverso de hermanos puede ser capaz de extraer más comida de su entorno que un clon, porque cada hermano utiliza un nicho ligeramente distinto. Uno de los principales defensores de esta hipótesis es Graham Bell, de la Universidad McGill. La hipótesis ha sido criticada porque no consigue explicar cómo desarrollaron el sexo las especies asexuales. En su libro Evolution and Human Behavior (MIT Press, 2000), John Cartwright comenta:
"Aunque fue popular, ahora la hipótesis del ribazo enmarañado parece afrontar muchos problemas, y sus anteriores defensores están disminuyendo. La teoría predice un mayor interés en el sexo entre los animales que producen muchas proles pequeñas que compiten entre sí. En realidad, el sexo está asociado invariablente con organismos que producen pocas proles de gran tamaño, mientras que los organismos que producen pequeñas proles con frecuencia realizan la partenogénesis [reproducción asexual]. Además, la evidencia de los fósiles sugiere que las especies pasan por vastos preiodos de tiempo [geológico] sin cambiar mucho."
El sexo ayuda a la difusión de los caracteres ventajosos
El sexo crea genotipos novedosos más rápidamente
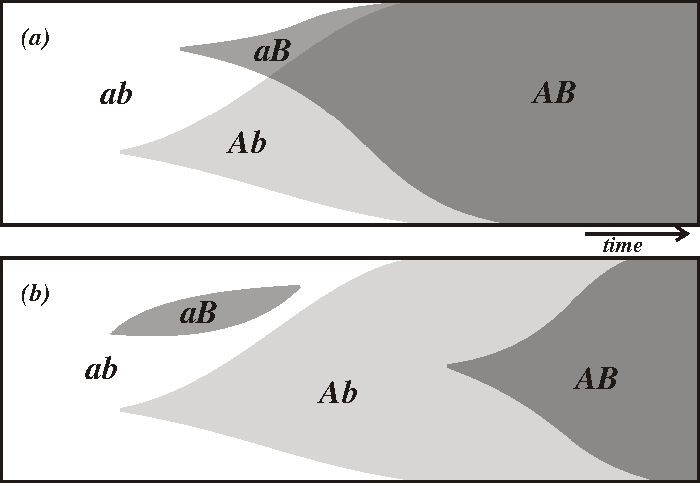
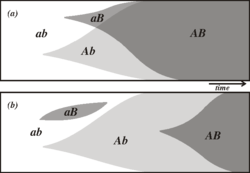 Este diagrama ilustra cómo puede el sexo crear genotipos novedosos de manera más rápida. Dos alelos ventajosos A y B aparecen por azar. Los dos alelos se recombinan rápidamente en (a), una población sexual, pero en (b), una población asexual, los dos alelos deben prosperar independientemente.
Este diagrama ilustra cómo puede el sexo crear genotipos novedosos de manera más rápida. Dos alelos ventajosos A y B aparecen por azar. Los dos alelos se recombinan rápidamente en (a), una población sexual, pero en (b), una población asexual, los dos alelos deben prosperar independientemente.El sexo podría ser un método por el cual se crean genotipos novedosos. Como el sexo combina los genes de dos individuos, las poblaciones con reproducción sexual pueden combinar más fácilmente los genes ventajosos que las poblaciones asexuales. Si en una población sexual aparecen dos alelos ventajosos distintos en diferentes locus de un cromosoma y en distintos miembros de una población, se puede producir un cromosoma que contenga los dos alelos ventajosos en pocas generaciones, por recombinación. Sin embargo, si aparecen esos mismos alelos en distintos miembros de una población asexual, la única manera de que un cromosoma pueda desarrollar el otro alelo es que sufra la misma mutación de manera independiente, algo que podría tardar mucho más.
Ronald Fisher también sugirió que el sexo puede facilitar la dispersión de los genes ventajosos permitiéndoles escapar de sus alrededores genéticos, en el caso de que aparecieran en un cromosoma con genes perjudiciales.
Pero estas explicaciones dependen de la tasa de mutación. Si las mutaciones favorables son tan raras que todas acabarán fijadas en la población antes de que aparezca la siguiente (teniendo en mente que la mutación es un proceso de Poisson, entonces las poblaciones sexuales y asexuales evolucionarían a la misma velocidad.
Además, estas explicaciones dependen de la selección de grupo, que es una fuerza selectiva débil relacionada con la selección natural –el sexo sigue siendo desventajoso para el individuo debido al doble coste del sexo. Por tanto, estas explicaciones no dan cuenta de por qué las especies "heterogónicas" eligen adoptar la reproducción sexual, como indicó George C. Williams en su argumento del equilibrio, y por tanto son insuficientes para explicar la evolución del sexo.
Los defensores de estas teorías responden al argumento del equilibrio que los individuos producidos por reproducción sexual y asexual pueden diferir en otros aspectos también –lo que puede influir en la persistencia de la sexualidad. Por ejemplo, en las pulgas de agua (cladóceros), la descendencia sexual forma huevos que son más capaces de sobrevivir al invierno.
El sexo aumenta la resistencia a los parásitos
Una de las teorías más aceptadas para explicar la evolución del sexo afirma que evolucionó como una adaptación para ayudar a los individuos sexuales en su resistencia a los parásitos, conocida como hipótesis de la "Reina Roja".[7] [8] [9]
Cuando el entorno cambia, alelos que anteriormente eran neutrales o perjudiciales pueden hacerse beneficiosos. Si el entorno cambia lo suficientemente rápido (es decir, entre generaciones), estos cambios en el entorno pueden hacer que el sexo sea ventajoso para el individuo. Estos cambios rápidos en el entorno están causados por la coevolución entre los huéspedes y los parásitos.
Imagine, por ejemplo, que en los parásitos hay un gen con dos alelos p y P que les confiere dos tipos de habilidad parasitaria, y un gen en los huéspedes con dos alelos h y H que les confiere dos tipos de resistencia a los parásitos, de manera que los parásitos con el alelo p pueden parasitar a los huéspedes con el alelo h, y los que tienen el P pueden parasitar a los que tienen el H. Esta situación conducirá a una serie de cambios cíclicos en las frecuencias alélicas –mientras p aumenta de frecuencia, h se hará desfavorable.
En la realidad habrá varios genes implicados en la relación entre los huéspedes y los parástos. En las poblaciones asexuales de huéspedes, la descendencia solo tendrá la resistencia a los parásitos distinta si surge por mutación. Sin embargo, en una población sexual de huéspedes, la descendencia tendrá una nueva combinación de alelos de resistencia a los parásitos.
En otras palabras, al igual que la reina roja de Lewis Carroll, los huéspedes sexuales se adaptan continuamente para adelantarse a sus parásitos.
La comparación entre el ritmo de evolución molecular de los genes de las quinasas y las inmunoglobulinas del sistema inmunitario y los genes que codifican a otras proteínas proporciona una evidencia para esta explicación de la evolución del sexo. Los genes que codifican las proteínas del sistema inmunitario evolucionan a una velocidad considerablemente mayor.[10] [11]
Los críticos de la hipótesis de la Reina Roja ponen en cuestión que este entorno que cambia constantemente provocado por los huéspedes y los parásitos sea lo bastante común para explicar la evolución del sexo.
El sexo ayuda a eliminar los genes perjudiciales
Las mutaciones pueden tener muchos efectos distintos sobre un organismo. En general se piensa que la mayoría de las mutaciones no neutrales son perjudiciales, lo que significa que provocan una disminución de la aptitud total del organismo.[12] Si una mutación tiene un efecto perjudicial, normalmente será eliminada de la población mediante la selección natural. Se cree que la reproducción sexual es más eficiente que la reproducción asexual a la hora de eliminar del genoma esas mutaciones.[13]
Hay dos hipótesis principales que eplican cómo puede actuar el sexo para eliminar del genoma los genes perjudiciales.
El sexo permite la reconstrucción de individuos libres de mutaciones
En una población asexual finita bajo la presión de mutaciones perjudiciales, la pérdida aleatoria de individuos sin esas mutaciones es inevitable. Esto se conoce como trinquete de Muller. Sin embargo, en una población sexual, se pueden recuperar individuos libres de mutaciones por recombinación de genotipos que contienen mutaciones perjudiciales.
Esta comparación solo funcionará para poblaciones pequeñas; en una población grande, la pérdida aleatoria del genotipo más adaptado se hace poco probable incluso para las poblaciones asexuales.
El sexo actúa fomentando la eliminación de los genes perjudiciales
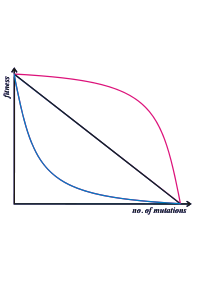 Este diagrama ilustra las diferentes relaciones entre el número de mutaciones y la aptitud. El modelo de Kondrashov requiere de epistasis sinérgico, que está representado por la línea roja — cada mutación tiene un efecto desproporcionadamente grande en la aptitud del organismo.
Este diagrama ilustra las diferentes relaciones entre el número de mutaciones y la aptitud. El modelo de Kondrashov requiere de epistasis sinérgico, que está representado por la línea roja — cada mutación tiene un efecto desproporcionadamente grande en la aptitud del organismo.Esta hipótesis fue propuesta por Alexei Kondrashov, y a veces se conoce como hipótesis de la mutación determinista.[13] Asume que la mayoría de las mutaciones perjudiciales son solo ligeramente perjudiciales, y afecta al individuo de manera que cada mutación adicional tiene un efecto cada vez mayor sobre la aptitud del organismo. Esta relación entre el número de mutaciones y la aptitud se conoce como epistasis sinérgico.
Piense por analogía en un coche con varias averías menores. Ninguna es suficiente para evitar que el coche ande pero, en combinación, las averías hacen que el coche no funcione.
De manera similar, un organismo puede que sea capaz de soportar unos pocos defectos, pero la presencia de muchas mutaciones puede superar sus mecanismos de respaldo.
Kondrashov afirma que la naturaleza ligeramente perjudicial de las mutaciones implica que la población tenderá a estar compuesta por individuos con un pequeño número de mutaciones. El sexo actuará recombinando estos genotipos, creando individuos con menos mutaciones e individuos con más mutaciones, y como existe una gran desventaja selectiva contra los individuos con muchas mutaciones, estos individuos se eliminan de la población. En esencia, el sexo compartimenta a las mutaciones perjudiciales.
La teoría de Kondrashov ha recibido mucha crítica. Primero, necesita una tasa de mutación muy alta – una mutación por generación, algo para lo que existe cierta evidencia empírica (por ejemplo en la Drosophila[14] y la E. coli[15] ). Segundo, también necesita que las mutaciones perjudiciales actúen de una forma sinérgica. Aunque existe cierta evidencia para este tipo de mutaciones, también existe la misma cantidad de evidencia de que las mutaciones actúan sinérgicamente. Puede que no exista epistasis (una mutación no tiene influencia sobre otra) o interacción antagónica (toda mutación adicional tiene un efecto desproporcionadamente pequeño).
Otras explicaciones
El sexo y la velocidad de la evolución
Ilan Eshel ha sugerido que el sexo evita que la evolución vaya demasiado rápido. Sugiere que la recombinación rompe las combinaciones favorables de genes más a menudo de lo que las crea, y el sexo se mantiene porque asegura la selección a más largo plazo que en las poblaciones asexuales — por tanto los cambios a corto plazo afectan menos a la población. Esta explicación no tiene mucha aceptación, ya que sus supuestos son muy restrictivos.
Recientemente se ha demostrado, en experimentos con algas Chlamydomonas, que el sexo puede eliminar el límite de velocidad de la evolución.[16]
Origen de la reproducción sexual
La reproducción sexual evolucionó hasta el proceso que es hoy, esto es, meiosis seguida de fertilización. Se producen gametos (el producto de la meiosis) en las eucariotas más primitivas que existen en la actualidad: los protistas.
Los organismos necesitan replicar su material genético de manera eficiente y fiable. La necesidad de reparar los daños genéticos es una de las teorías principales que explican el origen de la reproducción sexual. Los individuos diploides pueden reparar una sección mutada de su ADN mediante la recombinación genética, ya que hay dos copias del gen en la célula, y se supone que una de ellas permanece sin daños. Por otro lado, una mutación en un individuo haploide tiene más probabilidad de persistir, ya que la maquinaria de reparación del ADN no tiene manera de saber cuál era la secuencia original sin daños.[17] La forma más primitiva de sexo podría haber sido un organismo con ADN dañado replicando una hebra sin dañar de un organismo similar para repararse a sí mismo.[18]
Otra teoría es que la reproducción sexual se originó a partir de elementos genéticos parasitarios egoístas que intercambian material genético (esto es: copias de su propio genoma) para transmitirse y propagarse. En algunos organismos, se ha demostrado que la reproducción sexual mejora la difusión de los elementos genéticos parasitarios (por ejemplo en la levadura, un hongo filamentoso).[19] La conjugación bacteriana, una forma de intercambio genético que algunas fuentes describen como sexo, no es una forma de reproducción. Sin embargo, resplada la teoría del elemento genético egoísta, ya que se propaga mediante uno de esos "genes egoístas", el plásmido F.[18]
Una tercera teoría dice que el sexo evolucionó como una forma de canibalismo. Un organismo primitivo se comió a otro pero, en lugar de digerirlo completamente, parte del ADN del organismo 'comido' se incorporó al organismo 'comedor'.[18]
Notas
- ↑ John Maynard Smith The Evolution of Sex 1978.
- ↑ George C. Williams Sex and Evolution 1975, Princeton University Press, ISBN 0-691-08152-2
- ↑ Weismann, A. 1889. Essays on heredity and kindred biological subjects. Oxford Univ. Press, Oxford, UK
- ↑ Fisher, R. A. 1930. The genetical theory of natural selection. Clarendon Press, Oxford, UK
- ↑ Muller, H. J. 1932. Some genetic aspects of sex. Am. Nat. 8: 118–138.
- ↑ Burt, A. 2000. Perspective: sex, recombination, and the efficacy of selection—was Weismann right? Evolution 54: 337–351.
- ↑ Van Valen, L. (1973) "A New Evolutionary Law." Evolutionary Theory 1, 1-30.
- ↑ Hamilton, W. D. et al. "Sexual reproduction as an adaptation to resist parasites" 1990. Proceedings of the National Academy of Sciences (PNAS) 87:3566-3573.
- ↑ Matt Ridley 1995 The Red Queen: Sex and the Evolution of Human Nature 1995 Penguin.
- ↑ Kuma, K., Iwabe, N. & Miyata, T. 1995. Functional constraints against variations on molecules from the tissue-level - slowly evolving brain-specific genes demonstrated by protein-kinase and immunoglobulin supergene families. Molecular Biology and Evolution 12 (1): 123-130.
- ↑ Wolfe KH & Sharp PM. 1993. Mammalian gene evolution - nucleotide-sequence divergence between mouse and rat. Journal of molecular evolution 37 (4): 441-456 OCT 1993
- ↑ Griffiths et al. 1999. Gene mutations, p197-234, in Modern Genetic Analysis, New York, W.H. Freeman and Company.
- ↑ a b Kondrashov, A. S. "Deleterious mutations and the evolution of sexual reproduction" 1988. Nature 336:435-440.
- ↑ Whitlock, M. C., & Bourget, D. 2000. Factors affecting the genetic load in Drosophila: synergistic epistasis and correlations among fitness components. Evolution 54:1654–1660.
- ↑ Elena, S. F., & Lenski, R. E. 1997. Test of synergistic interactions among deleterious mutations in bacteria. Nature 390:395–398.
- ↑ Colegrave, N. 2002. Sex releases the speed limit on evolution. Nature 420: 664-666.
- ↑ Bernstein H, Byerly H, Hopf F, Michod R (1984). «Origin of sex.» J Theor Biol. Vol. 110. n.º 3. pp. 323-51. PMID 6209512.
- ↑ a b c Olivia Judson (2002). Dr. Tatiana's sex advice to all creation. New York: Metropolitan Books, pp. 233-4. ISBN 0-8050-6331-5.
- ↑ Hickey D (1982). «Selfish DNA: a sexually-transmitted nuclear parasite» Genetics. Vol. 101. n.º 3-4. pp. 519-31. PMID 6293914.
Referencias
- Graham Bell 1982. The masterpiece of nature. Croom Helm, London.
- George C. Williams Sex and Evolution 1975, Princeton University Press, ISBN 0-691-08152-2
- Maynard Smith, J. (1978) The Evolution of Sex. Cambridge University Press. ISBN 0-521-29302-2.
- Mark Ridley Evolution 1993, ISBN 0-632-03481-5
- Matt Ridley The Red Queen: Sex and the Evolution of Human Nature 1995, Penguin Reprint, ISBN 0-14-024548-0
- John Maynard Smith & Szathmary, E. The Major Transitions in Evolution 1995
- Mark Ridley (2004) Mendel's Daemon: Evolution and the Complexity of Life
- Hurst, L. D. and Peck, J. R. "Recent advances in the understanding of the evolution and maintenance of sex" 1996. Trends in Ecology and Evolution 11:46-52.
Véase también
Enlaces externos
Categoría: Evolución

Wikimedia foundation. 2010.