- Memoria icónica
-
La memoria icónica es el registro de la memoria sensorial relacionado con el dominio visual. Es un componente del sistema de memoria visual, que incluye también la memoria visual a corto plazo (MVCP) y la memoria a largo plazo (MLP). La memoria icónica se ha descrito como un almacén memorístico de muy breve duración (menos de 1000 ms), pre-categórico y de alta capacidad.[1] [2] Contribuye a la MVCP proporcionando una representación coherente de la percepción visual global durante un breve periodo de tiempo. La memoria icónica contribuye a explicar fenómenos como la ceguera al cambio y la continuidad de la experiencia durante los movimientos sacádicos. Actualmente no se considera que la memoria icónica sea una entidad individual, sino que está compuesta por al menos dos componentes diferenciados.[3] Los experimentos clásicos basados en el paradigma de Sperling del informe parcial, así como las técnicas actuales, continúan arrojando luz sobre la naturaleza de este almacén de memoria sensorial.
Contenido
Consideraciones generales
La existencia de la imagen fisiológica persistente de un objeto aún después de su desaparición física ha sido un fenómeno observado por mucha gente a través de la historia. Una de las primeras explicaciones documentadas del fenómeno fue realizada por Aristóteles, que propuso que las post-imágenes estaban relacionadas con los sueños.[4] La observación natural del haz de luz provocado por el movimiento rápido de un ascua ardiente suscitó el interés de los investigadores durante los años 1700 y 1800. Este tipo de estudios supuso el comienzo de la investigación empírica de este fenómeno,[4] que posteriormente se denominó persistencia visual.[3] En los años 1900, el papel de la persistencia visual en la memoria adquirió una importancia considerable, gracias a su hipotético papel como representación pre-categórica de la información visual en la MVCP. En 1960, George Sperling comenzó sus experimentos clásicos de informe parcial para confirmar la existencia de la memoria sensorial visual y determinar algunas de sus características, como su capacidad y duración.[1] No sería hasta el año 1967 cuando Ulric Neisser acuñara el término memoria icónica para este almacén memorístico de breve duración.[5]
Aproximadamente 20 años después de los experimentos originales de Sperling, comenzaron a definirse dos componentes diferenciados de la memoria sensorial visual: persistencia de la visión, y persistencia de la información. Los experimentos de Sperling se basaban principalmente en medir la información proporcionada por los estímulos, mientras que otros estudios, como los de Coltheart, se basaban en la ejecución de pruebas de medidas directas de la persistencia visual.[3] En 1978, Di Lollo propuso un modelo de dos estados de la memoria sensorial visual.[6] A pesar de que se ha debatido durante muchos años, la actual concepción de la memoria icónica establece una clara distinción entre la persistencia visual y la persistencia informativa, midiéndose de forma separada, y presentando diferentes propiedades. Se cree que la persistencia de la información, que se considera la base de la memoria icónica, es el factor clave para la memoria visual a corto plazo como un almacén sensorial pre-categórico.[7] [3]
Componentes de la memoria icónica
Los dos principales componentes de la memoria icónica son la persistencia visual y la persistencia informativa. La persistencia visual es una representación visual pre-categórica relativamente breve (150 ms) de la imagen física creada por el sistema sensorial. Sería como una "fotografía" de lo que el individuo está mirando y percibiendo. El segundo componente es un almacén de memoria de mayor duración que representa una versión codificada y ya categorizada de la imagen visual. Se trataría de los "datos en bruto" que posteriormente serán procesados por el cerebro. Puede considerarse un tercer componente: la persistencia neuronal: la actividad física y las grabaciones del sistema visual.[3] [8] La persistencia neuronal generalmente se representa mediante técnicas de neuroimagen como el electroencefalograma y la resonancia magnética de imágenes.
Persistencia de la visión
La persistencia visual es la impresión fenomenológica de que una imagen visual permanece después de la desaparición física del objeto que la provocó. Puede considerarse como el producto de la persistencia neuronal. La persistencia visual es más sensible a los parámetros físicos de los estímulos que la persistencia informativa, lo que se pone de manifiesto es sus dos propiedades clave:[3]
- La duración de la persistencia visual es inversamente proporcional a la duración del estímulo. Esto significa que cuanto más tiempo esté presente físicamente el estímulo, más rápidamente decaerá la imagen visual en la memoria.
- La duración de la persistencia visual es inversamente proporcional a la luminosidad del estímulo. Cuando la luminosidad o claridad de un estímulo aumenta, la duración de la persistencia visual disminuye.[2]
Debido a la implicación de los sistemas neuronales, la persistencia visual es altamente dependiente de la fisiología de los fotorreceptores y la activación de distintos tipos de células de la corteza visual. Esta representación visual puede verse afectada por efectos de enmascaramiento, por lo que la presentación de interferencias durante la presentación del estímulo a recordar, o inmediatamente después, puede interferir en la capacidad para recordar el estímulo posteriormente.[9]
Se han usado diversas técnicas para tratar de determinar la duración de la persistencia visual. La técnica de la duración de los estímulos consiste en que un estímulo de prueba (un "clic" auditivo) se presenta en el momento de la presentación de la imagen a recordar, y en un ensayo aparte, ese mismo estímulo se presenta en el momento de la desaparición de esa imagen. La diferencia representa la duración del almacén visual, que se ha determinado en aproximadamente 100-200 ms.[9] Otras técnicas, como la continuidad fenomenológica o la técnica moving slit estiman la duración de la persistencia visual en unos 300 ms.[10] La continuidad fenomenológica consiste en presentar de forma discontínua una imagen, con periodos en blanco entre las presentaciones. Si la duración de estos periodos en blanco es lo suficientemente corta, el participante tiene la sensación de estar viendo una imagen continua. De forma similar, la técnica moving slit también se basa en la observación de una imagen continua. Pero en lugar de presentar el estímulo completo "a ráfagas" (es decir, de forma discontinua), sólo se muestra un pequeño fragmento de la imagen (slit), que se va desplazando. Cuando este fragmento se desplaza a una velocidad adecuada, se puede observar una imagen completa.
Bases neurológicas de la persistencia visual
A la persistencia de visión subyace la persistencia neuronal de la ruta sensorial visual. Una representación visual prolongada comienza con la activación de los fotorreceptores de la retina. Aunque se ha encontrado que existe una persistencia en la activación de los bastones y los conos después de la supresión del estímulo, parece que los bastones se mantienen activados durante más tiempo que los conos.[11] Otras células implicadas en la persistencia de la imagen visual son las células ganglionares de la retina M y P. Las células M (células de transición) se activan únicamente cuando el estímulo aparece y desaparece. Las células P (células sostenidas) muestran una actividad continua mientras el estímulo está aparece, está presente y desaparece.[12] [11] Se ha hallado una persistencia cortical de la imagen visual en la corteza visual primaria (V1) del lóbulo occipital, que es la responsable del procesamiento de la información visual.[11] [13]
Persistencia de la información
La persistencia de la información se refiere a la información relacionada con un estímulo que persiste en la memoria después de su desaparición física. Es de naturaleza visual, pero no visible.[7] Los experimentos de Sperling medían la persistencia de la información.[3] La duración del estímulo es el factor clave en la duración de la persistencia de la información. A medida que aumenta la duración de la presencia estimular, aumenta la duración de la codificación visual.[14] Los componenten no visuales que representa la persistencia de la información incluyen las características abstractas de la imagen, así como su localización espacial. Debido a la naturaleza de la persistencia de la información, a diferencia de la persistencia de la visión, no se ve afectada por los efectos de enmascaramiento.[9] Las características de este componente de la memoria icónica sugieren que ésta desempeña un papel clave como un almacén de memoria post-categórico que proporciona a la MVCP la información que va a ser consolidada.[7]
 Se muestra la ruta dorsal (verde) y la ruta ventral (violeta) del procesamiento visual. Ambas rutas nacen de la misma fuente, en la corteza visual.
Se muestra la ruta dorsal (verde) y la ruta ventral (violeta) del procesamiento visual. Ambas rutas nacen de la misma fuente, en la corteza visual.
Bases neurológicas de la persistencia de la información
Aunque existe una minoría de investigaciones sobre la representación neuronal de la persistencia de la información que la equiparan a la persistencia visual, las nuevas técnicas electrofisiológicas han empezado a revelar la existencia de áreas corticales involucradas específicamente en este componente. A diferencia de la persistencia visual, se cree que la persistencia de la información reside en áreas visuales de mayor nivel que la corteza visual. En estudios realizados con macacos, se ha encontrado que el surco temporal superior anterior (STS), que forma parte de la ruta ventral del procesamiento visual, se encuentra activo durante la ejecución de tareas de memoria icónica. Esta región cerebral está asociada al reconocimiento y la identificación de objetos. El papel de la memoria icónica en la detección de cambios se ha relacionado con una activación del giro occipital medial (GOM). Se ha encontrado que la activación del GOM persiste durante aproximadamente 2000 ms, lo que sugiere la posibilidad de que la memoria icónica tenga una duración mayor de lo que se ha venido pronosticando. La memoria icónica también se ve influenciada por los genes y las proteínas producidas en el cerebro. El factor neurotrófico derivado del cerebro (FNDC) forma parte de las neurotrofinas, una familia de proteínas que estimulan la supervivencia, diferenciación y crecimiento de las neuronas. Los individuos con mutaciones en los genes que codifican el FNDC han mostrado una persistencia de la información más breve y menos estable.[15]
El papel de la memoria icónica
La memoria icónica proporciona un flujo de información visual al cerebro, que puede ser recogido y sostenido en el tiempo por la MVCP para su consolidación en formas de memoria más estables. Uno de los papeles claves de la memoria icónica es su implicación en la detección de cambios en el ambiente visual, lo que facilita la percepción del movimiento.[16]
Integración temporal
La memoria icónica activa la integración de la información visual proporcionando un flujo constante de imágenes; por ejemplo, al ver una película. En la corteza visual primaria, los nuevos estímulos entrantes no eliminan la información que han dejado los estímlos precedentes. En lugar de ello, las respuestas a los estímulos más recientes contienen la misma cantidad de información sobre el último estímulo percibido que sobre los precedentes.[13] Esta característica puede ser la principal responsable tanto del proceso de integración temporal como de los efectos de enmascaramiento. El resultado final dependerá del modo en que los dos componentes de la imagen adquieren significatividad; si son significativos únicamente por separado, se producirá un efecto de enmascaramiento; y si lo son significativos únicamente cuando se sobreponen, se producirá el efecto de integración.
Cequera al cambio
Se cree que la breve representación de la memoria icónica juega un papel esencial en la capacidad de detectar cambios en el entorno visual. El fenómeno de la ceguera al cambio ha proporcionado evidencias sobre la naturaleza del almacén de la memoria icónica y su papel en la visión. La ceguera al cambio hace referencia a la incapacidad para detectar diferencias entre dos escenas sucesivas separadas por un breve intervalo en blanco, o Intervalo Interestimular (IIE).[17] Cuando las escenas se presentan sin IIE, el cambio es fácilmente detectable. Se cree que el almacenamiento de los recuerdos detallados de la escena en la memoria icónica es eliminado por cada IIE, que hace inaccesible el recuerdo. Esto reduce la capacidad para realizar comparaciones entre escenas sucesivas.[17]
Movimientos oculares sacádicos
Se ha sugerido que la memoria icónica desempeña un papel proporcionando continuidad a la experiencia durante los movimientos oculares sacádicos.[18] Estos rápidos movimientos oculares tienen lugar durante aproximadamente 30 ms, y cada fijación dura unos 300 ms. Las investigaciones sugieren, no obstante, que la información adquirida entre movimientos sacádicos es altamente dependiente de la MVCP, y no de la memoria icónica. En lugar de contribuir a la memoria trans-sacádica, se cree que la información que se almacena en la memoria icónica es eliminada durante los movimientos sacádicos. Ocurre un fenómeno similar durante el parpadeo: ya sea automático o intencional, interrumpe la información almacenada en la memoria icónica.[19]
Desarrollo de la memoria icónica
El desarrollo de la memoria icónica comienza con el nacimiento, y continúa a medida que se desarrolla el sistema visual primario y secundario. A los cinco años de edad, los niños han desarrollado la misma capacidad ilimitada de memoria icónica que la que muestra un adulto. No obstante, la duración de la persistencia de la información sigue aumentando, desde los 200 ms a la edad de cinco años, hasta un nivel asintótico de 1000 ms en la edad adulta (a partir de los once años). Con el envejecimiento tiene lugar un leve descenso en la persistencia de la visión. Se ha observado un decrecimiento de aproximadamente 20 ms al comparar individuos de veinte años con individuos de 60.[20] A lo largo de la vida, el deterioro cognitivo leve puede provocar algunos errores de memoria episódica (recuerdos autobiográficos sobre personas, lugares y contextos) y de memoria de trabajo (el componente de procesamiento activo de la memoria a corto plazo), debido a daños en el hipocampo y las áreas corticales asociadas. Los recuerdos episódicos son son sucesos autobiográficos. Se ha demostrado que las personas con deterioro cognitivo leve muestran un decrecimiento en la capacidad y duración de la memoria icónica. Este empeoramiento puede ser utilizado como un predictor del desarrollo de déficits más severos, como la enfermedad de Alzheimer y la demencia.
Procedimiento de Sperling del informe parcial
En el año 1960, George Sperling se convirtió en el primer investigador que utilizó el paradigma del informe parcial para investigar el modelo bipartito de la MVCP.[1] En los experimentos iniciales de Sperling, se presentaba a los observadores estímulos visuales mediante un taquitoscopio durante un breve periodo de tiempo (50 ms). Los estímulos consistían en matrices de caracteres alfanuméricos de 3x3 o 3x4, como por ejemplo:
- P Y F G
- V J S A
- D H B U
El recuerdo se basaba en una pista que se proporcionaba inmediatamente después de la retirada del estímulo, y dirigía al sujeto a recuperar una línea específica de letras de la presentación inicial. Se comparaba el rendimiento bajo dos condiciones: informe global e informe parcial.
Informe global
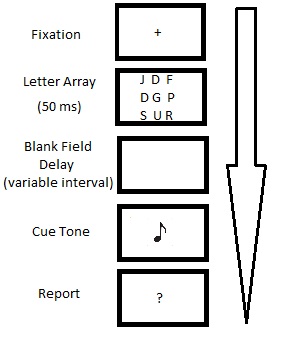
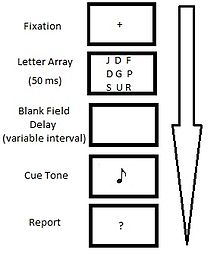 Paradigma original del informe parcial de Sperling.
Paradigma original del informe parcial de Sperling.La condición de informe global requería que los participantes recordaran cuantos elementos fueran posibles de la presentación original, en su ubicación específica. Por lo general, los participantes eran capaces de recordar tres o cuatro caracteres de los doce que se mostraban (~35%).[1] Esto sugiere que el informe global se ve limitado por un sistema de memoria con una capacidad de cuatro o cinco ítems.
Informe parcial
La condición de informe parcial requería que los participantes identificaran un subconjunto de los caracteres presentados utilizando algún indicio o pista indicativa. Esta pista consistía en un tono que sonaba a distintos intervalos temporales (~50 ms) inmediatamente después de la desaparición de los estímulos. La frecuencia del tono (alta, media o baja) indicaba qué subconjunto de caracteres de la presentación debían ser recordados. Debido al hecho de que los participantes no conocían de antemano qué columna se iba a pedir recordar, el desempeño en la condición de informe parcial puede ser entendida como una muestra aleatoria del recuerdo de un observador sobre la presentación completa. Este tipo de muestreo reveló que inmediatamente después de que el estímulo desaparecía, los participantes podían recordar la mayor parte de las letras (entre 9 y 12), lo que sugiere que el 75% de la presentación estimular completa era accesible a la memoria.[1] Esto supone un incremento muy notable en comparación con la capacidad de la memoria icónica que se había hipotetizado mediante los estudios de informe global.
Variaciones al procedimiento del informe parcial
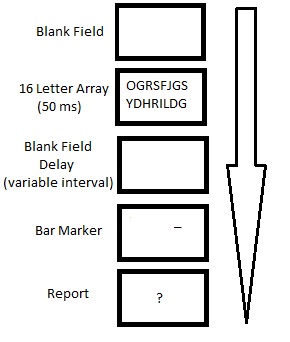
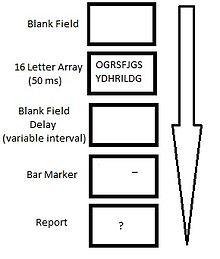 Paradigma de informe parcial de Averbach & Coriell.
Paradigma de informe parcial de Averbach & Coriell.Indicio de barra visual
Una pequeña variación a los procedimientos de informe parcial de Sperling que produjo rendimientos similares fue el uso de una barra visual marcadora en lugar de un tono auditivo como pista de recuperación. Bajo esta condición, a los participantes se les mostraba un estímulo visual compuesto por dos filas de 8 letras durante 50 ms. La pista de recuperación consistía en una barra ubicada encima o debajo de la posición de una letra, que se mostraba simultáneamente a la desaparición del estímulo. Los participantes mostraron una efectividad media del 65% cuando se les pidió que recordaran la letra designada.[21]
Variaciones temporales
La variación del tiempo transcurrido entre la desparición del estímulo y la presentación de la pista auditiva permitió a sperling estimar el tiempo de funcionamiento de la memoria sensorial. Sperling se desvió del procedimiento inicial variando el momento de presentación del tono: de presentar el tono inmediatamente después de la desaparición de l estímulo pasó a presentarlo 150, 500 o 1000 ms después. Mediante esta técnica, se encontró que el recuerdo de los estímulos decaía rápidamente. A los 1000 ms de diferencia entre el estímulo y la pista, no había diferencias en el recuerdo entre la condición de informe parcial y la de informe global. En términos generales, los experimentos que utilizaban el paradigma del informe parcial mostraron un rápido decaimiento de la huella de memoria sensorial al cabo de aproximadamente 1000 ms después de la desaparición del estímulo.[1] [21] [22]
Pista circular y enmascaramiento
Se identificaron los efectos del enmascaramiento mediante el uso de un círculo que rodeaba a la letra, como pista de recuerdo.[23] Cuando se presentaba el círculo antes que el estímulo visual aparecía, o justo en el momento en que desaparecía, el nivel de recuerdo se equiparaba al encontado en el caso de usar una barra o un tono como pistas. No obstante, si el círculo se rpesentaba 100 ms después de la desaparición del estímulo, había un descenso en el nivel de acierto en el recuerdo. A medida que aumentaba la demora en la presentación del círculo, la efectividad volvía a aumentar de nuevo. Este fenómeno es un ejemplo de enmascaramiento por metacontraste. También se observó efecto de enmascaramiento cuando se presentaban imágenes como líneas al azar inmediatamente después de la retirada del estímulo.[24]
Véase también
Referencias
- ↑ a b c d e f Sperling, George (1960). «The information available in brief visual presentations» (en inglés, PDF). Psychological Monographs 74 (11): pp. 1-29. http://aris.ss.uci.edu/HIPLab/staff/sperling/PDFs/Sperling_PsychMonogr_1960.pdf. Consultado el 30 de mayo de 2011.
- ↑ a b Dick, A. O. (1974). «Iconic memory and its relation to perceptual processing and other memory mechanisms» (en inglés). Perception & Psychophysics 16 (3): pp. 575-596. doi:. http://www.springerlink.com/content/r8927566267nl937/. Consultado el 30 de mayo de 2011.
- ↑ a b c d e f g Coltheart, Max (1980). «Iconic memory and visible persistence» (en inglés). Perception & Psychophysics 27 (3): pp. 183-228. doi:. PMID 6992093. http://www.springerlink.com/content/j86r40223271m02t/. Consultado el 30 de mayo de 2011.
- ↑ a b Allen, Frank (1926). «The persistence of vision» (en inglés). American Journal of Physiological Optics 7: pp. 439-457.
- ↑ Neisser, Ulric (1967) (en inglés). Cognitive Psychology. Nueva York: Appleton-Century-Crofts.
- ↑ Di Lollo, Vincent (1980). «Temporal integration in visual memory» (en inglés, PDF). Journal of Experimental Psychology: General 109 (1): pp. 75-97. doi:. PMID 6445405. http://wexler.free.fr/library/files/di%20lollo%20%281980%29%20temporal%20integration%20in%20visual%20memory.pdf. Consultado el 30 de mayo de 2011.
- ↑ a b c Irwin, David; James Yeomans (1986). «Sensory Registration and Informational Persistence» (en inglés). Journal of Experimental Psychology: Human Perception and Performance 12 (3): pp. 343-360. doi:. PMID 2943863.
- ↑ Loftus, Geoffrey; Janine Duncan, Paul Gehrig (1992). «On the time course of perceptual information that results from a brief visual presentation» (en inglés, PDF). Journal of Experimental Psychology 18 (2): pp. 530-549. doi:. PMID 1593234. http://faculty.washington.edu/gloftus/Downloads/LoftusDuncanGehrig.pdf. Consultado el 30 de mayo de 2011.
- ↑ a b c Long, Gerald (1980). «Iconic Memory: A Review and Critique of the Study of Short-Term Visual Storage» (en inglés). Psychological Bulletin 88 (3): pp. 785-820. doi:. PMID 7003642.
- ↑ Haber, R.; L. Standing (1969). «Direct measures of visual short-term visual storage» (en inglés). Quarterly Journal of Experimental Psychology 21 (1): pp. 43-54. doi:. PMID 5777982.
- ↑ a b c Irwin, David; Thomas, Laura (2008). «Neural Basis of Sensory Memory». En Steven Luck y Andrew Hollingworth (en inglés). Visual Memory. Nueva York: Oxford University Press. pp. 32-35. ISBN 978-0-19-530548-7.
- ↑ Levick, W.; J. Zacks (1970). «Responses of cat retinal ganglion cells to brief flashes of light» (en inglés, PDF). Journal of Physiology 206 (3): pp. 677-700. PMID 5498512. http://www.ncbi.nlm.nih.gov/pmc/articles/PMC1348672/pdf/jphysiol01062-0182.pdf.
- ↑ a b Nikolić, Danko; S. Häusler, W. Singer y W. Maass (2009). «Distributed fading memory for stimulus properties in the primary visual cortex» (en inglés, PDF). PLoS Biology 7. doi:. http://ts-si.org/files/journal.pbio.1000260.pdf. Consultado el 31 de mayo de 2011.
- ↑ Greene, Ernest (2007). «Information persistence in the integration of partial cues for object recognition» (en inglés, PDF). Perception & Psychophysics 69 (5): pp. 772-784. doi:. PMID 17929699. http://www.springerlink.com/content/f6481u7656426594/. Consultado el 31 de mayo de 2011.
- ↑ Beste, Christian; Daniel Schneider, Jörg Epplen, Larissa Arning (febrero-marzo 2011). «The functional BDNF Val66Met polymorphism affects functions of pre-attentive visual sensory memory processes» (en inglés, PDF). Neuropharmacology 60 (2-3): pp. 467-471. doi:. PMID 21056046. http://www.bio.psy.rub.de/papers/beste11bdnf.pdf. Consultado el 31 de mayo de 2011.
- ↑ Urakawa, Tomokazu; Koji Inui, Koya Yamashiro, Emi Tanaka, Ryusuke Kakigi (2010). «Cortical dynamics of visual change detection based on sensory memory» (en inglés). NeuroImage 52: pp. 302-308. doi:. PMID 20362678.
- ↑ a b Becker, M.; H. Pashler, S. Anstis (2000). «The role of iconic memory in change-detection tasks» (en inglés, PDF). Perception 29: pp. 273-286. doi:. PMID 10889938. http://www.pashler.com/Articles/BeckerPashlerAnstis_2000.pdf. Consultado el 31 de mayo de 2011.
- ↑ Jonides, J.; D. Irwin, S. Yantis (1982). «Integrating visual information from successive fixations» (en inglés, PDF). = Science 215 (4529): pp. 192-194. doi:. PMID 7053571. http://www-personal.umich.edu/~jjonides/pdf/1982.pdf. Consultado el 31 de mayo de 2011.
- ↑ Thomas, Laura; David Irwin (2006). «Voluntary eyeblinks disrupt iconic memory» (en inglés, PDF). Perception & Psychophysics 68 (3): pp. 475-488. doi:. PMID 16900838. http://www.springerlink.com/content/8155133737m35880/. Consultado el 31 de mayo de 2011.
- ↑ Walsh, David; Larry Thompson (1978). «Age Differences in Visual Sensory Memory» (en inglés, PDF). Journal of Gerontology 33 (3): pp. 383-387. doi:. PMID 748430. http://www.pnas.org/content/102/5/1797.full.pdf+html. Consultado el 31 de mayo de 2011.
- ↑ a b Averbach, E.; Sperling, G. (1961). «Short-term storage of information in vision». En C. Cherry (en inglés). Information Theory. Londres: Butterworth. pp. 196-211.
- ↑ Sperling, George (1967). «Successive approximations to a model for short-term memory» (en inglés). Acta Psychologica 27: pp. 285-292. doi:. PMID 6062221.
- ↑ Averbach, E.; A. Coriell (1961). «Short-term memory in vision» (en inglés, PDF). Bell Systems Techinical Journal 40: pp. 309-328. http://www.alcatel-lucent.com/bstj/vol40-1961/articles/bstj40-1-309.pdf. Consultado el 31 de mayo de 2011.
- ↑ Sperling, George (1963). «A model for visual memory tasks» (en inglés, PDF). Human Factors 5: pp. 19-31. PMID 13990068. http://aris.ss.uci.edu/HIPLab/staff/sperling/PDFs/Sperling_VM_Model_1963.pdf. Consultado el 31 de mayo de 2011.

Wikimedia foundation. 2010.