- Catecolamina
-
Las catecolaminas (también llamadas neurohormonas o aminohormonas) son neurotransmisores que se vierten al torrente sanguíneo (en lugar de las hendiduras sinápticas, como corresponde normalmente a los neurotransmisores). Son un grupo de sustancias que incluyen la adrenalina, la noradrenalina y la dopamina, las cuales son sintetizadas a partir del aminoácido tirosina. Contienen un grupo catecol y un grupo amino.
Las catecolaminas pueden ser producidas en las glándulas suprarrenales, ejerciendo una función hormonal, o en las terminaciones nerviosas, por lo que se consideran neurotransmisores. El precursor de todos ellos es la tirosina, que se usa como fuente en las neuronas catecolaminérgicas (productoras de catecolaminas).
Las catecolaminas están asociadas al estrés y la obesidad[cita requerida].
Contenido
Estructura
Las catecolaminas tienen la estructura distintiva de un anillo de benceno, con dos grupos hidroxilos, una cadena intermedia y un grupo amino terminal.
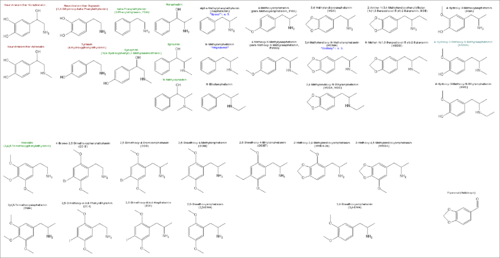 Estructura de las catecolaminas y sustancias relacionadas (en rojo, arriba a la izquierda.
Estructura de las catecolaminas y sustancias relacionadas (en rojo, arriba a la izquierda.
Metabolismo
Formación de catecolaminas
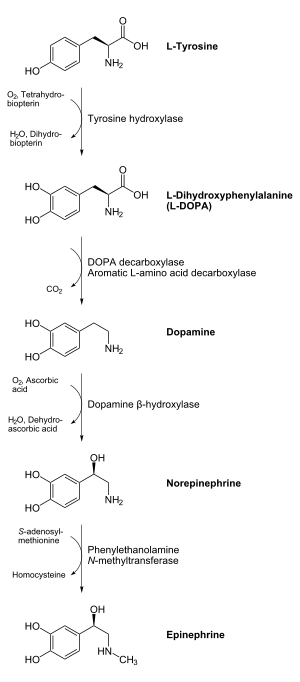 Biosíntesis de las catecolaminas.
Biosíntesis de las catecolaminas.Las catecolaminas se producen principalmente en las células cromafines de la médula adrenal y en las fibras postganglionares del Sistema nervioso simpático. La tirosina hidroxilasa (TH) se encuentra en todas las células que sintetizan catecolaminas y es la enzima de cantidad limitada en sus rutas biosintéticas. La TH es una oxidasa de acción combinada que usa el oxígeno molecular y la tirosina como sustratos y la biopterina como cofactor. Cataliza la adición de un grupo hidroxilo a la meta posición de la tirosina, formando de esta manera 3,4-dihidroxi-L-fenilalanina (L-dopa). A pesar de la disponibilidad de tirosina no limita ordinariamente la proporción de síntesis de aminas, el cofactor biopterina y el oxígeno podrían estar en concentraciones subsaturadas dentro de las neuronas que contienen catecolaminas, y de esta manera podrían representar un importantísimo papel en la regulación de la biosíntesis de la adrenalina. La siguiente reacción está catalizada por la DOPA descarboxilasa, que pasa L-Dopa a dopamina. Necesita piridoxal fosfato. El paso de dopamina a noradrenalina la cataliza la dopamina β-hidroxilasa, a partir de ascorbato y oxígeno. Por último, para que la noradrenalina pase a adrenalina se usa feniletanolamina N-metiltransferasa, que transfiere un grupo metilo de un donador (S-adonosilmetionina) hasta la adrenalina formante.
En condiciones normales, la concentración de tirosina es suficiente para mantener saturada la tirosina hidroxilasa. La tirosina hidroxilasa es un tetrámero, y una particularidad es que también puede hidroxilar la fenilalanina. La hidroxilación de fenilananina daría tirosina. Esto es útil con pacientes afectados por fenilcetonuria (que tiene deficiencia en la fenilananina hidroxilasa). La tirosina hidroxilasa podría ser un sustituyente. Cada subunidad pesa 60000 D.
La biosíntesis de catecolaminas es un proceso muy regulado. La regulación a largo plazo suele implicar la cantidad de enzimas reguladoras. Regula la cantidad de tirosina hidroxilasa, y la cantidad de dopamina β-hidroxilasa. A veces se necesitan cambios a corto plazo, y se regulan por mecanismos distintos:
- La enzima que cataliza la etapa limitante de velocidad (tirosina hidroxilasa) es inhibida por Dopa y dopamina, porque compiten con la biopterina por los sitios de unión.
- Regulación de la tirosina hidroxilasa por fosforilación. En cada subunidad hay residuos de serina (posiciones 8, 19, 31, 40) que se fosforilan. Los residuos 19 y 40 de serina causan un incremento más importante de la actividad al ser forforilados. El residuos 40 se fosforila sobre todo por la proteína quinasa A, y el 10 por la CAM quinasa II. La despolarización del terminal aumenta la actividad de la tirosina hidroxilasa, ya que entra calcio que activa las enzimas quinasas. Una vez las catecolaminas se han sintetizado se produce el almacenamiento en el interior de vesículas sinápticas conocidas como vesículas granulares o de núcleo denso. En el interior de las vesículas se encuentran unas sustancias llamadas cromograminas, calcio y ATP en alta concentración (1000 mM). Las catecolaminas están formando complejo con las cromograminas. También está la dopamina β-hidroxilasa, por lo cual la síntesis de noradrenalina tiene lugar en el interior de la vesícula, al menos en parte. El sistema por el que entran las catecolaminas en las vesículas es un sistema antiporte con protones. El gradiente de protones necesario lo realiza una protón-ATPasa bombeando al interior protones, por lo que el pH es de 5,5 aproximadamente. Este sistema de captación tiene amplia especificidad de sustrato. De manera que pueden competir con las catecolaminas endógenas.
Rutas metabólicas relacionadas
Inactivación
Las catecolaminas poseen una vida media de unos minutos cuando circulan por la sangre. El mecanismo de inhibición de las catecolaminas es de recaptación. Ese mecanismo lo tienen las neuronas que la liberan y las células gliales circundantes. Existen distintos tipos de transporte:
- NET (noradrenalina y adrenalina)
- DAT (dopamina)
- VMAT-2 (transportador de vesículas para cargarla de catecolamina).
Los dos primeros son dependientes del gradiente de sodio, que está dirigido hacia dentro. Una vez han sido recaptados a la terminal, son catabolizados por la monoamino oxidasa (MAO), y la catecol-o-metiltransferasa (COMT). La COMT está en todos los tejidos. La MAO se encuentra en la membrana externa de las mitocondrias, y provoca la desaminación oxidativa.
Las monoaminas son sustrato de la MAO y pasan a aldehídos, para después llegar a amionácidos gracias a otras enzimas. Hay dos MAOs, la A y la B. La MAO-A desamina preferentemente noradrenalina y serotonina. La MAO-B tiene un espectro más amplio. La MAO es numerosa en el intestino y el hígado, y cataboliza las aminas de la dieta, impidiendo que entren en la circulación general. Las anfetaminas no son sustrato de la MAO. La COMT está incluso en los eritrocitos, y se localiza en la membrana plasmática. Transfiere un grupo metilo desde la 5-adenosilmetionina hasta el anillo catecol, al grupo 3 del anillo.
El ácido homovanílico es el metabolito más importante del catabolismo de la dopamina. Del catabolismo es el 3-metoxi-4-hidroxifeniglicol si hablamos de la noradrenalina, en el líquido cefalorraquídeo. Para saber el índice de recambio de noradrenalina en el Sistema Nervioso Central no vale medir la orina, pues los niveles no serán iguales (ya que existe la contribución del Simpático en la orina al ser la noradrenalina un neurotransmisor en el simpático).
Receptores
Hay muchos procesos clave de liberación de catecolaminas:
- Los receptores adrenérgicos (noradrenalina y adrenalina): Estos dos neurotransmisores tienen una gran diversidad de efectos, que se explica por la presencia de diferentes receptores, que en cada tipo de célula están acoplados a vías de transducción distintas. En el músculo liso puede producir contracción si se activan los receptores α, y se relajan si actúan sobre los receptores β2. En los vasos sanguíneos producen vasoconstricción y vasorelajación. En los bronquios producen broncodilatación (al revés que los vasos). En el tubo digestivo provoca constricción y relajación. En el corazón aumenta la frecuencia cardíaca y su intensidad; incrementando el gasto cardíaco. Los receptores adrenérgicos están estructuralmente relacionados, pero tienen distintos segundos mensajeros. Se distinguen receptores α y β; adrenalina y noradrenalina son agonistas para ambos receptores, pero estos tienen más agonistas y antagonistas. El receptor α puede ser α1 ó α2. El α1 puede ser A, B ó D. Estos tres se diferencian en los antagonistas, la localización, la estructura y el mecanismo efector (adenilato ciclasa). En este caso, lo que importa es que en cada sitio del organismo la adenilato ciclasa causa un efecto distinto. Los β pueden ser 1, 2 ó 3. Difieren en los antagonistas, y las características. Pero los 3 estimulan la adenilato ciclasa.
- Los receptores dopaminérgicos: Son, igual que los anteriores, metabotrópicos. Pueden ser D1, D5, D3, D4 y D2. Los D2 pueden ser S (short) o L (long). Los D1 y D5 estimulan la adenilato ciclasa. Los otros tres la inhiben, pero los D2 y D4 activan canales de potasio. Los D2 pueden inhibir un canal de calcio. Aparte del agonista común (dopamina), cada canal tiene sus propios agonistas. Algunos, como el sulpirilo y la clozapina tienen efectos anti-psicóticos. El AMPc activa a la proteína quinasa A, que puede provocar respuestas a corto plazo, o más largas a través de factores de transcripción, de manera directa o indirecta. Esta última está mediada por el gen de respuesta inmediata (IEG). A corto plazo es fosforilación, y a largo plazo es alterando la expresión génica. También hay receptores en la terminal presináptica (autorreceptores), y también en otras terminales presinápticas que no liberan dopamina. Los D4 y D2 son los que tienen demostrada esta característica.
Funciones
Dos catecolaminas, la noradrenalina y la dopamina, actúan como neurotransmisores en el Sistema Nervioso Central y como hormonas en el torrente sanguíneo. Las catecolaminas causan generalmente cambios fisiológicos que preparan al cuerpo para la actividad física (como la lucha, la huida,...).
Disfunciones
Las disfunciones en las vías catecolaminérgicas son trastornos bipolares y esquizofrenia. La primera evidencia de esto se obtuvo con los inhibidores de la MAO, que tenían efectos antidepresivos. Los antidepresivos tricíclicos son inhibidores de sistemas de recaptación de alta afinidad, predominantemente la noradrenalina. Esto ha llevado a formular la teoría catecolaminérgica de estos desórdenes: "Ciertos desórdenes de las vías catecolaminérgicas en el Sistema Nervioso llevan a la depresión". Por el contrario, un aumento de la actividad provocaría la fase maniaca. Esto lo hacen por ejemplo las anfetaminas, ya que compiten por los lugares de recaptación de la nordrenalina. De manera más reciente se ha usado también con la serotonina. La Fluoxetina es un fármaco que bloquea la recaptación de serotonina y trata la depresión. Las enfermedades psicóticas mejoran con fármacos del sistema dopaminérgico. Se ha relacionado las vías dopaminérgicas como componente de la esquizofrenia, que son altas concentraciones de dopamina. Se tratan con los antagonistas de la vía, o sea, sulfirilo y clozapina.
Funciones motrices
En las funciones motrices, la dopamina está implicada en la enfermedad del Parkinson. Hay una degeneración de las neuronas de la sustancia negra. Desde la sustancia nigra hay dos vías que se dirigen a los ganglios basales (destacando el núcleo caudado), que juegan un papel en el control del movimiento. El núcleo caudado modula el tálamo, y desde aquí hay una inervación que controla la corteza motriz. Así pues, se corta la modulación de la dopamina entre la sustancia nigra y el núcleo caudado. La dopamina no atraviesa la barrera hematoencefálica, así que se utiliza la L-Dopa, que sí lo hace y a partir de ella se mejora, pero no corrige.
Catecolaminas sobre el Sistema Inmunitario
Las investigaciones han revelado que con estrés se disparaban los niveles de catecolaminas, y que los linfocitos tenían receptores adrenérgicos. Cuando se incubaba norepinefrina (noradrenalina) o epinefrina (adrenalina), se veía su capacidad de proliferación frente a fitohemaglutinina (PHA) o concavalina A (ConA), y se comprobaba que a 10-4 M inhibían la producción de linfocitos T, pero que concentraciones de 10-8 M y en presencia de hidrocortisona estimulaba la proliferación de estos linfocitos T. Esto efecto se inhibía si se añadía fentolamina (bloqueante de receptores α). Se vio que concentraciones muy bajas de epinefrina que en apariencia no ejercían efectos sobre la proliferación, estimulaban esta proliferación cuando se añadía propranolol (bloqueante de receptores β). En conclusión, las respuestas a concentraciones altas se inhiben con propanolol, así que se necesitan receptores β. Los receptores β y α son antegónicos. Se solía decir que las vías de AMPc son inhibitorias y las de GMPc activas. Esto sólo funciona con la proliferación de los linfocitos T. En otros casos no tiene porqué ser cierto.
Efecto "in vitro" de las catecolaminas sobre los macrófagos
Solo concentraciones de 10 -12 M aumentan significativamente la quimiotaxis de macrófagos. Pero los receptores α son más importantes en este proceso. Concentraciones farmacológicas (altas) o fisiológicas de norepinefrina estimulan la capacidad fagocítica de los macrófagos, a no ser que uno de los receptores esté bloqueado. En cuanto a la capacidad microbicida, se necesita el efecto de sumación conjunta de receptores α y β.
Se supone que la quimiotaxis necesita menos concentración de norepinefrina que la fagocitosis porque en los vasos no hay mucha cantidad, pero cuando los macrófagos son atraídos al foco infeccioso, la epinefrina y norepinefrina actúan autocrinamente sobre las células aumentando la concentración en un lugar localizado. Concentraciones de 10-5 a 10-12 M son además quimioatrayentes para los fagocitos.
Quimiotaxis de los linfocitos en los órganos inmunocompetentes
Cuando están confinados en un órgano linfoide, con mucha concentración de norepinefrina, los linfocitos permanecen allí acumulándose por si se produce una infección cuando terminen de madurar.
Efectos "in vitro" de las catecolaminas en las células NK
Concentraciones mayores o iguales a 10-7 M de norepinefrina inhiben la proliferación NK. Menores de 10-7 M la estimulan bajo determinadas circunstancias. Este efecto se ha visto que influye directamente sobre las células NK y no sobre las células tumorales.
Modelo de actuación de catecolaminas "in vivo" en respuesta a linfocitos
La actuación se divide en tres fases temporales: fase inductiva, proliferativa y efectora.
- Fase inductiva: La noradrenalina liberada por las terminaciones nerviosas estimula la fagocitosis y presentación antigénica así como la colaboración celular en general a través de receptores α y β adrenérgicos. Se favorece el inicio de la respuesta inmunitaria. Las citoquinas producidas por los macrófagos (fundamentalmente la IL-1) inhiben la acción de la noradrenalina disminuyendo su concentración.
- Fase proliferativa: Concentraciones bajas de noradrenalina a través de receptores α estimulan la proliferación de los linfocitos T. Segregan IL-2 que también estimula la producción de noradrenalina. Concentraciones de noradrenalina altas por receptores β inhiben la proliferación de los linfocitos, con lo que se para y regula la respuesta proliferativa.
- Fase efectora: La noradrenalina, a través de receptores β, disminuye la producción de anticuerpos o la actividad de linfocitos citotóxicos (CTL).
Véase también
Bibliografía
- "Hypoglycemia" by Ronald Hoffman, M.D., July 1999, The Holistic M.D.
- Joh, T.H., Hwang, O. 1987. Dopamine beta-hydroxylase: biochemistry and molecular biology. Ann N Y Acad Sci. 493:342-50.
Enlaces externos
Categorías:- Catecolaminas
- Neurotransmisores

Wikimedia foundation. 2010.