- Chondrichthyes
-
 Peces cartilaginosos
Peces cartilaginososRango temporal: Silúrico - Presente 
Tiburón blancoClasificación científica Reino: Animalia Filo: Chordata Subfilo: Vertebrata Clase: Chondrichthyes
Huxley, 1880Subclases Los condrictios (Chondrichthyes, del griego χονδρος khóndros, "cartílago" y ιχθύς ikhthýs, "pez") son una clase de vertebrados acuáticos conocidos como peces cartilaginosos, denominación que hace referencia a que su esqueleto es de cartílago, a diferencia del de los peces óseos (osteictios), que lo tienen de hueso.
Esta clase incluye a las subclases elasmobranquios (tiburones, rayas) y holocéfalos (quimeras).
Contenido
Características generales
Los condrictios presentan un mosaico de caracteres evolucionados y primitivos. Entre los rasgos primitivos destaca su anatomía básica. Entre los rasgos evolucionados destacan dos: la suspensión y estructura de las aletas y la estructura y composición de las mandíbulas y dentición. Esto último en diferentes especies aparece con formas más evolucionadas o más primitivas. Otro rasgo muy avanzado es su sistema inmunitario. Desde el mesozoico las especializaciones de los tiburones van quedando claras, colocándolos en la cúspide de las redes tróficas marinas. Hay en este grupo una característica tendencia al gigantismo, como medida para evitar la depredación. El tiburón típico tiene una longitud de dos metros, y una raya típica de un metro. Estas proporciones resultan muy grandes para el estándar de los vertebrados.
Los dientes no están fusionados a la mandíbula y los van reemplazando por otros nuevos de forma continua, rápida y en serie gracias a una cavidad que tienen a lo largo del borde de la mandíbula. Esto les permite tener siempre dientes nuevos frente a aquellos que se van rompiendo, desgastando y desprendiendo. Existen dientes aserrados, con función cortadora; dientes afilados, con función agarradora y dientes planos (en muchas rayas) para moler el alimento.[1]
Nadan ayudados por aletas y respiran a través de branquias durante toda su vida. Éstas están expuestas al exterior directamente a través de 5 ó 7 hendiduras branquiales en el caso de rayas y tiburones, y 1 exclusivamente en las quimeras.[1]
Difieren de otros peces en poseer esqueleto formado principalmente por cartílago y no por hueso. Carecen de opérculo y vejiga natatoria. La falta de esta última les obliga a nadar constantemente o posarse en el fondo (como hacen algunas rayas y tiburones), sin poder mantener una posición estática en la columna de agua.
Sentidos
Tienen un agudo sentido del olfato. Son capaces de detectar sangre y seguirle el rastro hasta encontrarla. La vista es menos aguda: pueden detectar luces y sombras en el agua. Y los órganos que poseen en las líneas laterales (sistema ambulacral de Lorenzini) y hocico les permiten captar los estímulos eléctricos de las contracciones musculares de los peces óseos. que con rapdidez acuden a su presa
Clasificación
Subclase Elasmobranchii
Superorden Orden Nombre vernáculo Batoidea (Rayas) Rajiformes Rayas verdaderas Myliobatiformes Pristiformes Peces sierra Torpediniformes Peces torpedo Selachimorpha (Tiburones) Hexanchiformes Squaliformes Pristiophoriformes Squatina Heterodontiformes Orectolobiformes Carcharhiniformes Subclase Holocephali
Orden Familia Nombre vernáculo Chimaeriformes Callorhynchidae Rhinochimaeridae Chimaeridae Quimeras Evolución
 Cladoselache, un condrictio del Devónico
Cladoselache, un condrictio del Devónico
Los condictios hicieron su aparición durante el periodo Devónico Superior. Los fósiles más primitivos que se conocen corresponden al género Cladoselache.[2] A lo largo la historia los condrictios han contado con dos grandes periodos de radiación evolutiva, la primera tuvo lugar hasta principios del Pérmico, para luego pasar por una fase de declive que duró 100 millones de años; la segunda gran radiación se dio hacia el Cretáceo.[2]
Anatomía
Anatomía externa
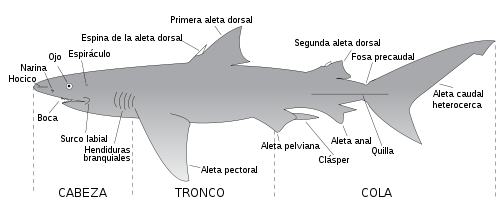 Anatomía externa de un condríctio elasmobranquio.
Anatomía externa de un condríctio elasmobranquio.Los elasmobranquios están comprimidos lateralmente y presentan aspecto fusiforme con aletas pares e impares. Las aletas impares son la dorsal (o dorsales), la caudal (que es heterocerca) y la anal. Las aletas pares son las pectorales y las pelvianas insertándose éstas siempre por detrás de las pectorales.
Presentan diversos orificios a lo largo del cuerpo, tales como los orificios nasales, la boca, las hendiduras branquiales, la foseta cloacal, el espiráculo (que es el resto de la primera hendidura branquial), y las cuencas orbitales de los ojos.
El tegumento tiene escamas placoideas (también llamadas dentículos dérmicos) que se forman en la dermis y emergen atravesando la epidermis. Las capas de las escamas placoideas de fuera a dentro, al igual que las capas de los dientes de las que son homólogas, son: esmalte, dentina y pulpa. A lo largo de la evolución, estas escamas se han ido aligerando y simplificando. Están orientadas hacia atrás, y poseen diversas funciones tales como conseguir que la piel sea flexible y a la vez muy resistente y minimizar el rozamiento y viscosidad del medio acuático debido a la reducción de las turbulencias generadas por la locomoción.
Natación
Para funcionar con eficacia es importante una buena capacidad locomotora. El movimiento de los peces se debe a que el agua es incompresible, y al ejercer una fuerza sobre ella ésta responde con un vector igual y opuesto (R1). R1 tiene un componente lateral (L1) y otro frontal (F1). Al ejercer fuerzas laterales sucesivas, e intercalando las de sentido opuesto, se van sumando los componentes frontales y anulando unos a otros los componentes laterales. Éste es el movimiento ondulatorio. También se emplea el movimiento de batido, que propulsa agua hacia atrás para desplazar al animal.
La dirección del movimiento debe ser controlada, lo cual requiere el control de tres movimientos de desvío: el cabeceo, el balanceo y la guiñada.
Las aletas impares oponen su superficie al balanceo y a la guiñada. Las aletas pares se oponen al cabeceo (lo controlan).
También interviene el movimiento de la aleta caudal. En los tiburones la aleta caudal es heterocerca. Una aleta heterocerca, al batir, no sólo produce empuje, sino también una fuerza de suspensión (fuerza ascensional), que controla la posición en la columna de agua. Existen no obstante dudas sobre la auténtica función de la caudal heterocerca. Los peces óseos que tienen aleta heterocerca, según los datos de que disponemos, no parece que obtengan ninguna fuerza ascensional de ella. Los tiburones carecen de vejiga natatoria, pero aun así logran tener una densidad aproximadamente igual a la del agua de mar gracias a su hígado. La fuerza ascensional que produciría la aleta caudal al batir les permitiría controlar la profundidad a la que nadan. Para controlar su flotabilidad, los tiburones tragan aire, pero lo más importante en este aspecto es el hígado, que es muy grande y presenta una elevada cantidad de lípidos (escualeno). El efecto del hígado hace que la corrección por densidad al sumergir un tiburón sea suficiente para que éste no se hunda en el agua de mar.
Los tiburones son capaces, mediante el control del equilibrio de almacenamiento-reabsorción de lípidos en el hígado, de controlar su flotabilidad. Las especies bentónicas no tienen tanta riqueza de gotas lipídicas en el hígado.
Reproducción
 Huevo; nótese el zarzillo que le permite adherirse a la vegetación.
Huevo; nótese el zarzillo que le permite adherirse a la vegetación.La fecundación es interna. Los pterigópodos de las aletas de los machos son órganos copuladores. El macho introduce uno de los pterigópodos en la cloaca de la hembra, donde se da un efecto de succión del esperma hacia el oviducto. Se observa una fuerte tendencia en todo el grupo al ovoviviparismo. En ciertos casos se da el oviparismo o incluso viviparismo.
También se presenta el fenómeno de "oofagia" (canibalismo dentro del útero, en donde un embrión se come restos de huevos).
Referencias
- ↑ a b Varios autores (2005). «Peces cartilaginosos». Animal. La definitiva e impactante guía visual de la vida salvaje en nuestro planeta. Madrid: Pearson Educación, S.A.. ISBN 84-205-3616-4.
- ↑ a b Mgar.net - Tiburones: evolución
Enlaces externos
 Wikimedia Commons alberga contenido multimedia sobre Chondrichthyes. Commons
Wikimedia Commons alberga contenido multimedia sobre Chondrichthyes. Commons Wikiespecies tiene un artículo sobre Chondrichthyes. Wikispecies
Wikiespecies tiene un artículo sobre Chondrichthyes. Wikispecies
Lic. Andrea Liliana Duarte Villar(Paraguaya)



Wikimedia foundation. 2010.