- Flagelo
-
Flagelo
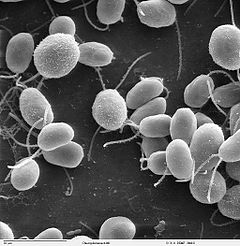
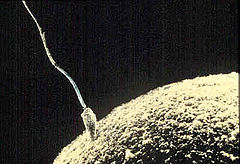
Un flagelo es un apéndice movible con forma de látigo presente en muchos organismos unicelulares y en algunas células de organismos pluricelulares.[1] [2] Un ejemplo es el flagelo que tienen los espermatozoides.[3] Usualmente los flagelos son usados para el movimiento, aunque algunos organismos pueden utilizarlos para otras funciones. Por ejemplo, los coanocitos de las esponjas poseen flagelos que producen corrientes de agua que estos organismos filtran para obtener el alimento.
Existen tres tipos de flagelos: eucarióticos, bacterianos y arqueanos. De hecho, en cada uno de estos tres dominios biológicos, los flagelos son completamente diferentes tanto en estructura como en origen evolutivo. La única característica común entre los tres tipos de flagelos es su apariencia superficial. Los flagelos de Eukarya (aquellos de las células de protistas, animales y plantas) son proyecciones celulares que baten generando un movimiento helicoidal. Los flagelos de Bacteria, en cambio, son complejos mecanismos en los que el filamento rota como una hélice impulsado por un microscópico motor giratorio. Por último, los flagelos de Archaea son superficialmente similares a los bacterianos, pero son diferentes en muchos detalles y se consideran no homólogos.
Contenido
Flagelo eucariota
En los eucariontes, los flagelos son estructuras poco numerosas, uno o dos por célula, con la excepción de algunos protistas unicelulares del grupo de los Excavata. Se distingue a las células acrocontas, que nadan con su flagelo o flagelos por delante, de las opistocontas, donde el cuerpo celular avanza por delante del flagelo. Esta última condición, evolutivamente más moderna, caracteriza a la rama evolutiva que reúne a los reinos hongos (Fungi) y animales (Animalia). Es la que observamos, sin ir más lejos, en los espermatozoides animales (incluidos, desde luego, los humanos).
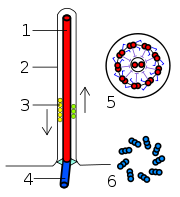 Estructura del flagelo eucariota. 1-axonema, 2-membrana plasmática, 3-IFT (Transporte IntraFlagelar), 4-cuerpo basal, 5-sección del flagelo, 6-tripletes de microtúbulos del cuerpo basal.
Estructura del flagelo eucariota. 1-axonema, 2-membrana plasmática, 3-IFT (Transporte IntraFlagelar), 4-cuerpo basal, 5-sección del flagelo, 6-tripletes de microtúbulos del cuerpo basal.
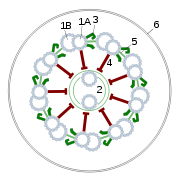 Sección transversal del flagelo mostrando el axonema. 1-doblete externo de microtúbulos, 2-doblete interno de microtúbulos, 3-dos brazos de dineína, uno interno y otro externo, 4-radio, 5-puente de nexina, 6-membrana plasmática.
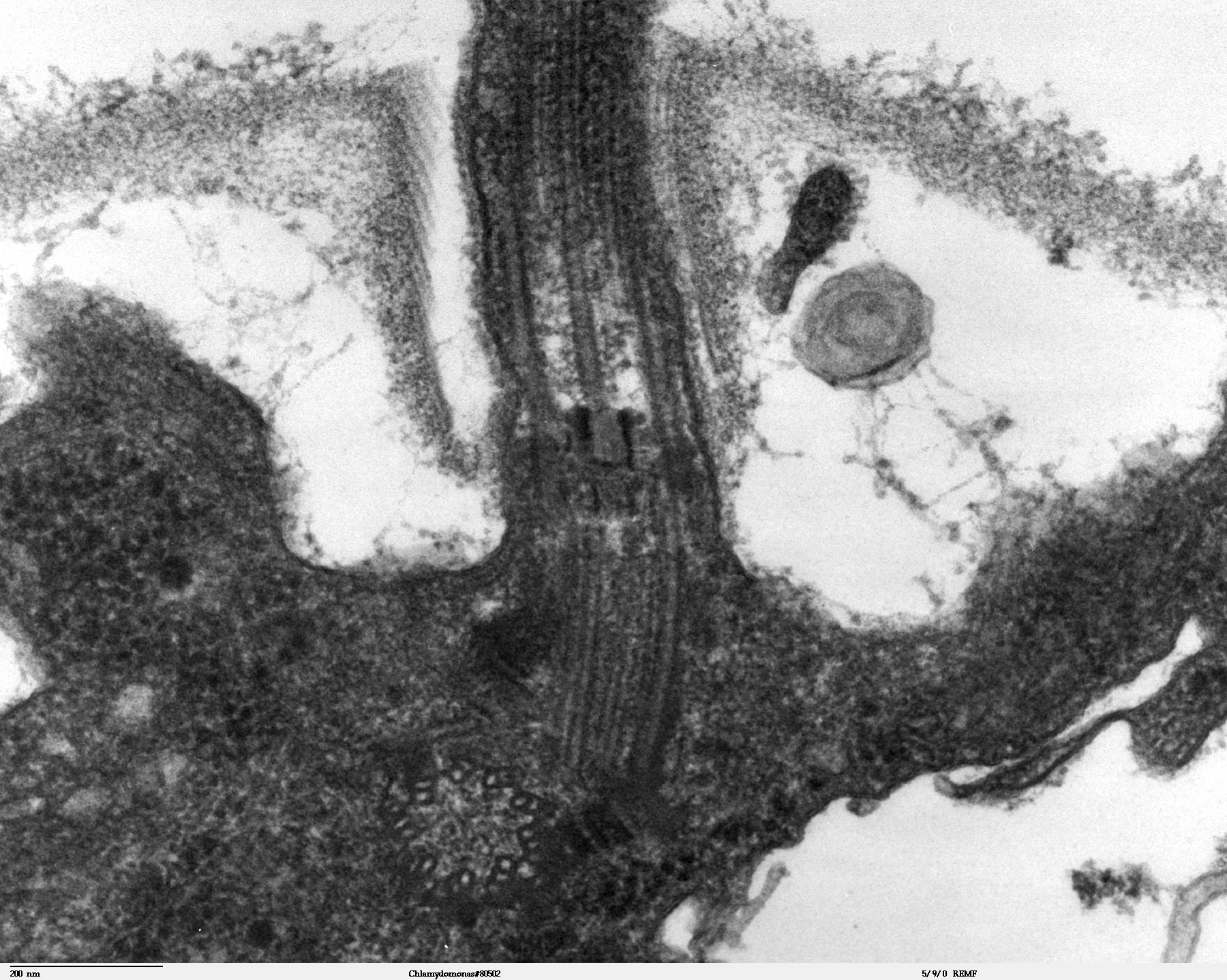
Sección transversal del flagelo mostrando el axonema. 1-doblete externo de microtúbulos, 2-doblete interno de microtúbulos, 3-dos brazos de dineína, uno interno y otro externo, 4-radio, 5-puente de nexina, 6-membrana plasmática.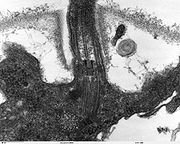 Sección longitudinal de Chlamydomonas reinhardtii. Se aprecia el inicio del flagelo y el cuerpo basal.
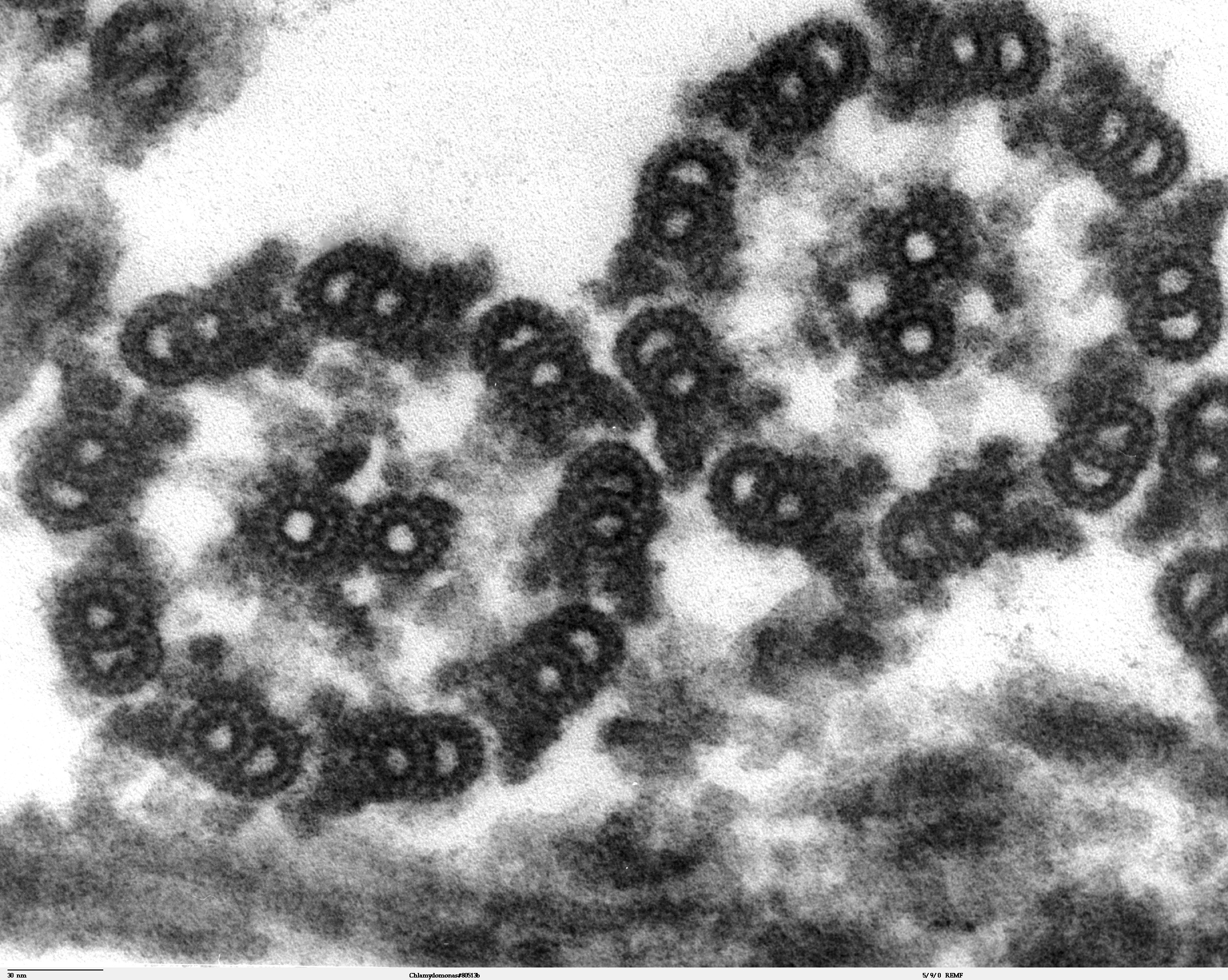
Sección longitudinal de Chlamydomonas reinhardtii. Se aprecia el inicio del flagelo y el cuerpo basal.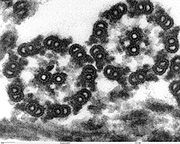 La estructura "9+2" del axonema es visible en esta micrografía de una sección de dos flagelos eucariotas.
La estructura "9+2" del axonema es visible en esta micrografía de una sección de dos flagelos eucariotas.Estructura
Junto con los cilios, los flagelos eucariotas constituyen un grupo de estructuras conocidas como undulipodios. Su ultraestructura es esencialmente la misma, pero el flagelo generalmente se complica con otros elementos añadidos, resultando más grueso y más largo.[4] Otra diferencia es el patrón de batido (véase más abajo).
Esencialmente, la estructura del flagelo (al igual que la del cilio) es una forma cilíndrica, de diámetro uniforme en toda su longitud, con una terminación redondeada, semiesférica. Su núcleo es un cilindro de nueve dobletes de microtúbulos que rodean a otros dos centrales.[5] Esta estructura "9+2" se denomina axonema. Este núcleo se encuentra cubierto por la membrana plasmática, a fin de que el interior del flagelo sea accesible al citoplasma de la célula. En la base del flagelo eucariota se encuentra un cuerpo basal (también denominado blefaroplasto o cinetosoma), que es el centro de organización de microtúbulos para los microtúbulos flagelares, de alrededor de 500 nm de largo. Los cuerpos basales son estructuralmente idénticos a los centriolos.
Cada uno de los dobletes de microtúbulos exteriores tiene asociado un par de brazos de dineína (uno interior y otro exterior). Estos brazos de dineína producen fuerza a través de la hidrólisis de ATP y consiguen que el flagelo se doble. De esta forma se consigue el batido de los flagelos. El axonema flagelar también contiene radios hechos de complejos de polipéptido que se extienden desde cada uno de los dobletes de microtúbulos exteriores hacia la pareja central, con la "cabeza" dirigida hacia el interior. Se cree que la cabeza interviene en la regulación del movimiento flagelar, aunque su función exacta y método de acción todavía no son bien entendidas.
Entre los dobletes de microtúbulos y la membrana se sitúan las partículas asociadas con el transporte intraflagelar (IFT), un proceso importante en el ensamblado del flagelo y el tráfico de proteínas en este compartimento celular. El movimiento de las subunidades axonemales, receptores transmembrana y otras proteínas arriba y abajo de la longitud del flagelo es esencial para el buen funcionamiento de este, tanto en la motilidad, como en la transducción de señales.[6] En la dirección de salida de la célula, IFT es impulsado por cinesinas, mientras que en la dirección de entrada es impulsado por dineínas.
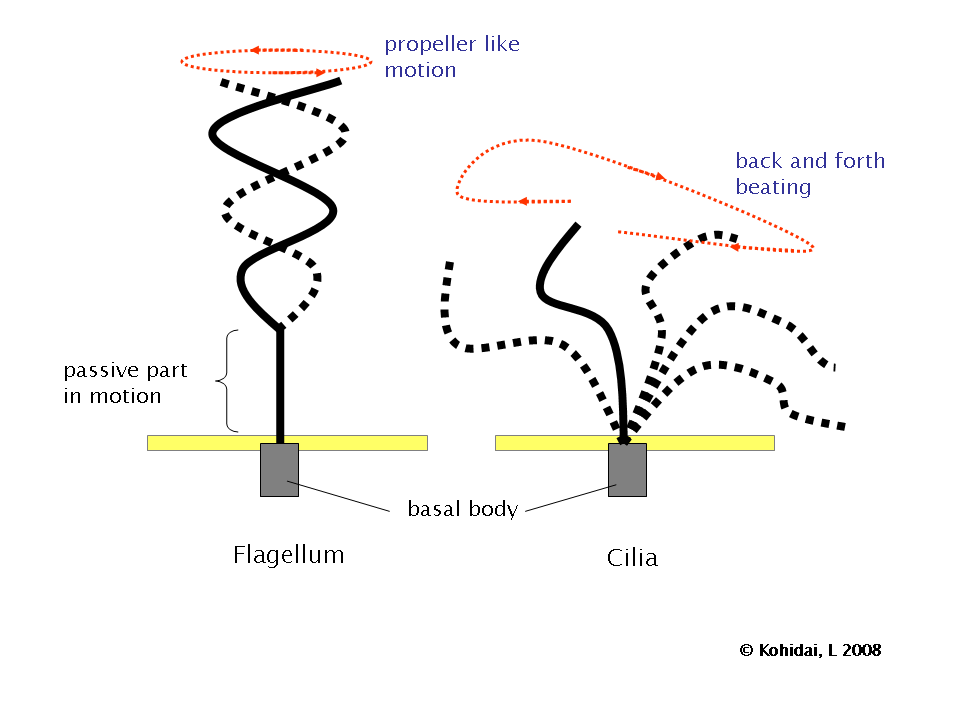
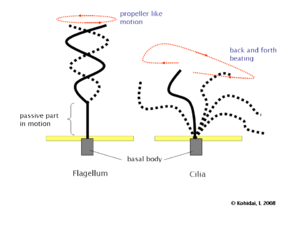 Diferencia de patrón de batido entre flagelos y cilios. El flagelo realiza un movimiento helicoidal mientras que el cilio realiza movimientos cíclicos atrás y adelante, como un remo.
Diferencia de patrón de batido entre flagelos y cilios. El flagelo realiza un movimiento helicoidal mientras que el cilio realiza movimientos cíclicos atrás y adelante, como un remo.Los flagelos más estudiados son los de espermatozoides. En el espermatozoide de mamíferos, el flagelo (cola) está constituido por un axonema rodeado por las fibras externas densas (uno por cada doblete) que intervienen en el movimiento del flagelo. Por fuera de estas fibras, existen otras estructuras rodeando el complejo axonema-fibras: la vaina mitocondrial, si el corte es por la pieza intermedia, o la vaina fibrosa, si el corte se realiza en la pieza principal. La vaina mitocondrial está constituida por mitocondrias dispuestas en hélice que proporcionan la energía necesaria para el movimiento del flagelo. La vaina fibrosa son pares de estructuras proteicas (cada una rodea la mitad de las fibras densas). Parece que intervienen en la protección del axonema y quizás también en el movimiento del flagelo. Por fuera, de todo ello, se dispone la membrana plasmática.
Patrones de batido de flagelos y cilios
Aunque flagelos y cilios eucariotas son idénticos en ultraestructura, estos dos tipos de apéndices tienen patrones de batido diferentes. Los flagelos, que impulsan a los espermatozoides y a muchos protistas, están diseñados para que uno sólo de ellos (o unos pocos) pueda impulsar a la célula completa a través de un fluido. El batido del flagelo genera un movimiento helicoidal sin que el eje rote sobre sí mismo, como ocurre en el giro de una honda. En contraste, los cilios están diseñados para actuar coordinadamente con otros muchos sobre la superficie celular. Su batido consiste en movimientos cíclicos, primero hacia atrás (propulsado rígido) y luego hacia adelante (recuperación flexible), como ocurre en el batido de un remo.
Flagelo bacteriano
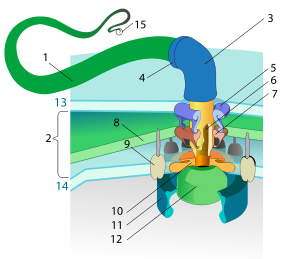 El flagelo bacteriano es un apéndice movido por un motor rotatorio. El rotor puede girar a 6.000-17.000 rpm, pero el apéndice usualmente sólo alcanza 200-1000 rpm. 1-filamento, 2-espacio periplásmico, 3-codo, 4-juntura, 5-anillo L, 6-eje, 7-anillo P, 8-pared celular, 9-estátor, 10-anillo MS, 11-anillo C, 12-sistema de secreción de tipo III, 13-membrana externa, 14-membrana citoplasmática, 15-punta.
El flagelo bacteriano es un apéndice movido por un motor rotatorio. El rotor puede girar a 6.000-17.000 rpm, pero el apéndice usualmente sólo alcanza 200-1000 rpm. 1-filamento, 2-espacio periplásmico, 3-codo, 4-juntura, 5-anillo L, 6-eje, 7-anillo P, 8-pared celular, 9-estátor, 10-anillo MS, 11-anillo C, 12-sistema de secreción de tipo III, 13-membrana externa, 14-membrana citoplasmática, 15-punta.El flagelo bacteriano es una estructura única, completamente diferente de los demás sistemas orgánicos utilizados por los seres vivos para el movimiento. Realmente presenta una similitud notable con los sistemas mecánicos artificiales, pues es una compleja estructura compuesta de varios elementos (piezas) y que rota como una hélice.
Los flagelos están compuestos por cerca de 20 proteínas, con aproximadamente otras 30 proteínas para su regulación y coordinación.[1] El filamento es un tubo hueco helicoidal de 20 nm de espesor. El filamento tiene una fuerte curva justo a la salida de la membrana externa; este "codo" permite convertir el movimiento giratorio del eje en helicoidal. Un eje se extiende entre el codo y el cuerpo basal, pasando por varios anillos de proteínas en la membrana de la célula que actúan como cojinetes. Las bacterias Gram-negativas tienen cuatro de estos anillos: el anillo L que se asocia con la membrana externa (lipopolisacáridos), el anillo P que se asocia con la pared celular (capa de peptidoglicano), el anillo MS que se inserta directamente en la membrana plasmática, y el anillo C que se une a la membrana plasmática. Las bacterias Gram-positivas sólo tienen dos anillos: MS y C. El filamento termina en una punta de proteínas.[7] [8]
El flagelo bacteriano está impulsado por un motor rotativo compuesto por proteínas (estátor, complejo Mot), situado en el punto de anclaje del flagelo en la membrana plasmática. El motor está impulsado por la fuerza motriz de una bomba de protones, es decir, por el flujo de protones (iones de hidrógeno) a través de la membrana plasmática bacteriana. Este bombeo se produce debido al gradiente de concentración creado por el metabolismo de la célula. (En Vibrio hay dos tipos de flagelos, laterales y polares, y algunos son impulsados por una bomba de iones de sodio en lugar de la bomba de protones[9] ). El rotor puede girar a 6.000-17.000 rpm, pero el filamento por lo general sólo alcanza 200-1000 rpm.
Los flagelos no giran a una velocidad constante, sino que aumentan o disminuyen su velocidad de rotación en relación con la fuerza motriz de protones. Las bacterias pueden alcanzar a través del medio líquido una velocidad de hasta 60 longitudes de célula/segundo. Aunque esto representa sólo 0,00017 km/h, al comparar esta velocidad con la de organismos superiores en términos de número de longitudes del cuerpo por segundo, es extremadamente rápido. El más rápido de los animales terrestres, el guepardo, corre a una velocidad máxima de alrededor de 110 km/h, pero esto representa sólo alrededor de 25 longitudes de cuerpo/segundo. Por tanto, cuando el tamaño se tiene en cuenta, las células procarióticas que nadan velocidades de 50-60 longitudes de cuerpo/segundo son en realidad mucho más rápidas que los organismos más grandes.
Los componentes del flagelo bacteriano son capaces de autoensamblaje sin ayuda de enzimas o de otros factores. Tanto el cuerpo basal como el filamento tienen un hueco central, a través del cual las proteínas del flagelo son capaces de moverse a sus respectivas posiciones. Durante el montaje, las proteínas que forman el filamento se añaden a la punta en lugar de en la base.
El flagelo bacteriano está relacionado con el complejo de poro y con el sistema de secreción de tipo III, una jeringa molecular que las bacterias utilizan para inyectar toxinas en otras células. Dadas las similaridades, se piensa que tanto el flagelo como el sistema de secreción se han originado a partir del complejo de poro. Además, el sistema de secreción de tipo III parece ser una simplificación del flagelo, pues está formado por subconjunto de componentes del flagelo.
Disposición de los flagelos
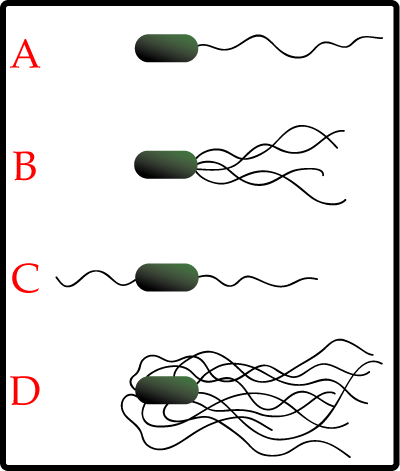
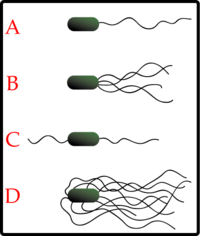 Los diferentes tipos de disposición de los flagelos bacterianos: A-Monotrico; B-Lofotrico; C-Anfitrico; D-Peritrico.
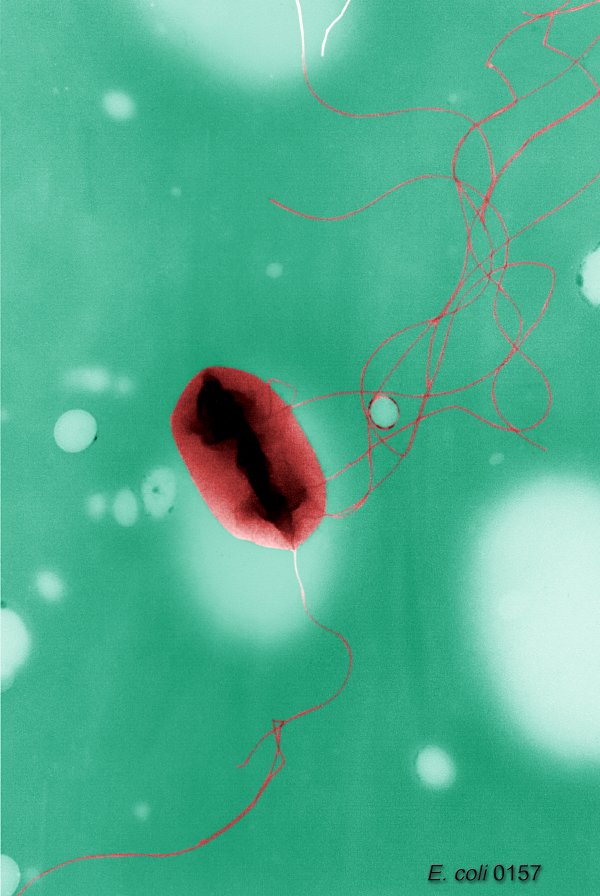
Los diferentes tipos de disposición de los flagelos bacterianos: A-Monotrico; B-Lofotrico; C-Anfitrico; D-Peritrico.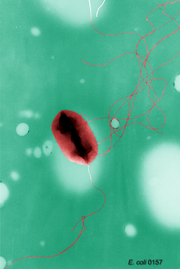 Escherichia coli, una bacteria peritrica.
Escherichia coli, una bacteria peritrica.Distintas especies de bacterias tienen diferente número y localización de los flagelos. Las bacterias monotricas presentan un solo flagelo (por ejemplo, Vibrio cholerae). Las bacterias lofotricas tienen múltiples flagelos situados en el mismo punto (o en dos puntos opuestos) que actúan en concierto para conducir a las bacteria en una sola dirección. En muchos casos, las bases de los múltiples flagelos están rodeadas de una región especializada de la membrana plasmática, denominada membrana polar. Las bacterias anfitricas tienen un solo flagelo en cada uno de los dos extremos opuestos (un solo flagelo opera a la vez, permitiendo a la bacteria revertir rápidamente el movimiento cambiando el flagelo que está activo). Las bacterias peritricas tienen flagelos que se proyectan en todas las direcciones (por ejemplo, Escherichia coli).
En algunas bacterias, tales como las especies más grandes de Selenomonas, los flagelos se organizan fuera de la célula enroscándose helicoidalmente unos con otros para formar una gruesa estructura denominada fascículo. Otras bacterias como las espiroquetas tienen un tipo especializado de flagelo conocido como filamento axial situado intracelularmente en el espacio periplásmico, que produce la rotación de toda la bacteria para avanzar con un movimiento similar al de un sacacorchos.
La rotación de los flagelos monotricos polares empuja la célula hacia delante con los flagelos atrás. Periódicamente, la dirección de rotación se invierte brevemente, procuciendo un viraje en la célula. Esto se traduce en la reorientación de la célula. Cuando la bacteria se desplaza en una dirección favorable el viraje es poco probable. Sin embargo, cuando la dirección del movimiento es desfavorable (por ejemplo, lejos de un producto químico atrayente), es más probable la realización de un viraje, con la posibilidad de que la célula se reoriente así en una dirección favorable.
En algunos Vibrio (en particular, Vibrio parahemolyticus[10] ) y en las formas relacionadas de proteobacterias como Aeromonas, coexisten dos sistemas flagelares codificados por diferentes conjuntos de genes e impulsados por diferentes gradientes de iones. Los flagelos polares se suelen utilizar cuando nadan en líquidos, mientras que los flagelos laterales entran en fucionamiento cuando los primeros experimentan gran resistencia al giro y proporcionan movilidad en fluidos viscosos o sobre superficies.[11] [12] [13] [14] [15] [16]
Flagelo arqueano
El flagelo de las arqueas es superficialmente similar al bacteriano pero no es homólogo. Ambos flagelos consisten en filamentos que se extienden fuera de la célula y rotan para impulsar al microorganismo. En el decenio de 1980 se pensaba que eran homólogos;[17] sin embargo, nuevos descubrimientos en el decenio de 1990 pusieron de manifiesto numerosas diferencias de detalle entre los flagelos bacterianos y arqueanos. Entre ellas se incluyen:
- Los flagelos bacterianos son impulsados por un flujo de iones H+ (u ocasionalmente Na+), mientras que los flagelos arqueanos son con casi todal seguridad impulsados por ATP. El par motor que produce la rotación del flagelo todavía no ha sido identificado.
- Mientras que las células bacterianas suelen tener muchos filamentos flagelares, cada uno de los cuales gira independiente, el flagelo arqueano se compone de un conjunto de filamentos que girar como uno sólo.
- Los flagelos bacterianos crecen por la incorporación de subunidades de flagelina en la punta, mientras que flagelos de las arqueas crecen por la incorporación de subunidades a la base.
- Los flagelos bacterianos son más gruesas que los arqueanos. Además, en las bacterias, los filamentos son tubos con un hueco lo suficientemente grande para que la subunidades de flagelina puedan fluir por el interior del filamento y depostarse en la punta; los flagelos de las arqueas son demasiado delgados para permitir esto.
- Muchos de los componentes de los flagelos bacterianos comparten secuencias similares a los del sistema de secreción de tipo III, pero los componentes de los flagelos de bacterias y arqueas no comparten similitud de secuencia. En lugar de ello, algunos componentes de los flagelos arqueanos comparten secuencia y similitud morfológica con componentes de los pili de tipo IV, que son ensamblados a través de la acción de los sistemas de secreción de tipo II (la nomenclatura de los pili y de los sistemas de secreción de proteínas no son coherentes).
Estas diferencias implican que los flagelos bacterianos y arqueanos son un caso clásico de evolución convergente, en lugar de homología. Sin embargo, en comparación con las décadas de estudio del flagelo bacteriano, los flagelos arqueanos sólo recientemente han comenzado a recibir atención científica seria. Por lo tanto, en muchas publicaciones se asume erróneamente que ambos flagelos son homólogos.
Referencias
- ↑ a b Bardy SL, Ng SY, Jarrell KF (February de 2003). «Prokaryotic motility structures» Microbiology (Reading, Engl.). Vol. 149. n.º Pt 2. pp. 295–304. DOI 10.1099/mic.0.25948-0. PMID 12624192.
- ↑ Lefebvre PA (2001). «Assembly and Motility of Eukaryotic Cilia and Flagella. Lessons from Chlamydomonas reinhardtii» Plant Physiol.. Vol. 127. n.º 4. pp. 1500–1507. DOI 10.1104/pp.010807. PMID 11743094.
- ↑ Malo AF, Gomendio M, Garde J, Lang-Lenton B, Soler AJ, Roldan ER (June de 2006). «Sperm design and sperm function» Biol. Lett.. Vol. 2. n.º 2. pp. 246–9. DOI 10.1098/rsbl.2006.0449. PMID 17148374.
- ↑ Haimo LT, Rosenbaum JL (December de 1981). «Cilia, flagella, and microtubules» J. Cell Biol.. Vol. 91. n.º 3 Pt 2. pp. 125s–130s. DOI 10.1083/jcb.91.3.125s. PMID 6459327.
- ↑ D.R. Mitchell (2006), The Evolution of Eukaryotic Cilia and Flagella as Motile and Sensory Organelles en Eukaryotic Membranes and Cytoskeleton: Origins and Evolution, editado por Gáspár Jékely, Eurekah.com, ISBN: 978-0-387-74020-1.
- ↑ Pazour GJ (October de 2004). «Intraflagellar transport and cilia-dependent renal disease: the ciliary hypothesis of polycystic kidney disease» J. Am. Soc. Nephrol.. Vol. 15. n.º 10. pp. 2528–36. DOI 10.1097/01.ASN.0000141055.57643.E0. PMID 15466257.
- ↑ Macnab RM (2003). «How bacteria assemble flagella» Annu. Rev. Microbiol.. Vol. 57. pp. 77–100. DOI 10.1146/annurev.micro.57.030502.090832. PMID 12730325.
- ↑ Diószeghy Z, Závodszky P, Namba K, Vonderviszt F (2004). «Stabilization of flagellar filaments by HAP2 capping» FEBS Lett.. Vol. 568. n.º 1-3. pp. 105–9. DOI 10.1016/j.febslet.2004.05.029. PMID 15196929.
- ↑ Atsumi T, McCarter L, Imae Y. (1992). «Polar and lateral flagellar motors of marine Vibrio are driven by different ion-motive forces» Nature. Vol. 355. pp. 182–4. DOI 10.1038/355182a0. PMID 1309599.
- ↑ Kim YK, McCarter LL (2000). «Analysis of the Polar Flagellar Gene System of Vibrio parahaemolyticus» Journal of Bacteriology. Vol. 182. n.º 13. pp. 3693–3704. DOI 10.1128/JB.182.13.3693-3704.2000. PMID 10850984.
- ↑ Atsumi T, Maekawa Y, Yamada T, Kawagishi I, Imae Y, Homma M (1996). «Effect of viscosity on swimming by the lateral and polar flagella of Vibrio alginolyticus» Journal of Bacteriology. Vol. 178. n.º 16. pp. 5024–5026. PMID 8759871.
- ↑ McCarter LL (2004). «Dual Flagellar Systems Enable Motility under Different Circumstances» Journal of Molecular Microbiology and Biotechnology. Vol. 7. pp. 18–29. DOI 10.1159/000077866. PMID 15170400.
- ↑ Merino S, Shaw JG, Tomás JM. (2006). «Bacterial lateral flagella: an inducible flagella system» FEMS Microbiol Lett. Vol. 263. pp. 127–35. DOI 10.1111/j.1574-6968.2006.00403.x. PMID 16978346.
- ↑ Belas R, Simon M, Silverman M. (1986). «Regulation of lateral flagella gene transcription in Vibrio parahaemolyticus» J Bacteriol. Vol. 167. pp. 210–8. PMID 3013835.
- ↑ Canals R, Altarriba M, Vilches S, Horsburgh G, Shaw JG, Tomás JM, Merino S (2006). «Analysis of the Lateral Flagellar Gene System of Aeromonas hydrophila AH-3» Journal of Bacteriology. Vol. 188. n.º 3. pp. 852–862. DOI 10.1128/JB.188.3.852-862.2006. PMID 16428388.
- ↑ Canals R, Ramirez S, Vilches S, Horsburgh G, Shaw JG, Tomás JM, Merino S (2006). «Polar Flagellum Biogenesis in Aeromonas hydrophila» Journal of Bacteriology. Vol. 188. n.º 2. pp. 542–555. DOI 10.1128/JB.188.2.542-555.2006. PMID 16385045.
- ↑ Cavalier-Smith T (1987). «The origin of eukaryotic and archaebacterial cells» Ann. N. Y. Acad. Sci.. Vol. 503. pp. 17–54. PMID 3113314.
Enlaces externos
- Motile Behavior of Bacteria (Physics Today on the Web)
- Revealing the mystery of the bacterial flagellum — A self-assembling nanomachine with fine switching capability — (Nanonet)
Categorías: Célula | Bacteria | Archaea | Motilidad celular



Wikimedia foundation. 2010.