- Genética del desarrollo
-
El desarrollo de un individuo multicelular ocurre a partir de un cigoto que prolifera mediante mitosis y mediante el proceso de determinación celular. En un principio todas y cada una de las células que constituyen el embrión pueden convertirse en cualquier tipo celular, son células totipotentes, pero en la mayoría de los individuos tras algunas divisiones del embrión cada célula determina a qué tipo celular corresponderá y ya no podrá volver a formar otro tipo de célula. La genética del desarrollo estudia cómo a partir de una célula aparece un organismo completo a nivel intracelular, a nivel de los genes y de su expresión o no expresión. Las etapas que engloba el desarrollo temprano en animales son:
- Fecundación: por fusión de dos gametos surge el cigoto que acabará constituyendo el organismo. En mamíferos el gameto no es un óvulo propiamente dicho, sino que es un ovocito ya que está detenido en metafase de segundo orden, y pasa a óvulo una vez fecundado. Dentro de la fecundación se distinguen varias fases: aproximación, activación del ovocito, penetración y anfimixis (en mamíferos)
- Segmentación:mediante divisiones por mitosis se forman primero blastómeros que a medida que se dividen van bajando por la trompa de Falopio hacia el útero. Divisiones sucesivas originan la mórula y finalmente la blástula. Después de la segmentación ocurre la compactación que consiste en los procesos que comunican los blastómeros entre sí e impedirían su separación si no hubiera zona pelúcida. Ya las células internas forman el embrioblasto que formará más adelante el embrión, y las células externas forman el trofoblasto que dará lugar a la placenta
- Gastrulación: menos divisiones mitóticas, comienzan los movimientos morfogenéticos al desplazarse conjuntos de células. Se forman las tres hojas embrionarias: ectodermo, mesodermo y endodermo.
- Organogénesis: el embrión experimenta la organización estructural, se delimitan los órganos.
- Histogénesis: diferenciación de tejidos: epitelial, glandular, conjuntivo, sanguíneo, muscular y nervioso.
La genética es muy importante a la hora de estudiar el desarrollo ya que la expresión de los genes regula eventos muy importantes en el mismo, es importante por tanto el estudio del control genético del desarrollo.
Contenido
Organismos modelo
Para el estudio de procesos biológicos , (en nuestro caso el control genético del desarrollo) se utilizan determinados organismos con una serie de características favorables para dicho cometido. Estos organismos son los llamados organismos modelo. Son útiles para el estudio genético porque son pequeños y por tanto se les puede observar cómodamente, tienen ciclos de vida y tiempo de regeneración cortos, capacidad para ser mantenidos a bajo coste, una descendencia manejable en cuanto al número y adapatabilidad al ambiente del laboratorio. Hay seis organismos modelos muy utilizados por genetistas: suaveeeee
- Escherichia coli (bacteria del intestino de algunos mamíferos, incluidos humanos)
- Saccharomyces cerevisiae (levadura de cerveza)
- Caenorhabditis elegans (nematodo)
- Drosophila melanogaster (mosca de la fruta; véase embriogénesis en Drosophila)
- Arabidopsis thaliana (planta de la familia de la mostaza)
- Mus musculus (ratón doméstico)
- Arabidopsis thaliana (planta) (véase desarrollo floral)
Aunque no son las únicas especies; otras también a menudo utilizadas en estudios genéticos son: Neurospora crassa (moho del pan), Zea mays (maíz), Danio rerio (pez cebra).
Entre los genetistas el modelo de control genético del desarrollo más utlilizado es Drosophila melanogaster, de la cual se han aislado numerosos mutantes para diferentes aspectos del desarrollo de la mosca. Las mutaciones se analizan para recopilar información sobre los genes que controlan el desarrollo temprano y cómo lo hacen. El cuerpo de Drosophila tiene tres partes: cabeza, tórax (con tres segmentos: uno con un par de patas, otro con un par de patas y otro de alas y otro con un tercer par de patas y los ronzales) y abdomen. Una vez producida la fecundación, el embrión tempranose desarrolla en tres fases: 1.Se desarrollan el eje antero posterior y el dorsoventral. 2.Se determinan la orientación y el número de segmentos que compondrán el cuerpo 3.Se establece la identidad de cada uno de los segmentos Cada una de estas fases o etapas distintas están controladas por un grupo de genes diferente.
Genes de polaridad del huevo
Estos genes son los que van a determinar la constitución de los dos ejes principales de desarrollo de Drosophila: el eje dorsoventral y el anteroposterior. Este conjunto de genes se transcriben a mRNA durante la ovogénesis y se traducirá después de la fecundación, afectando posteriormente al fenotipo del individuo. Por eso también se les llama genes maternos.
- Genes que determinan el eje dorsoventral: se encargan del dorso y el vientre de la mosca de la fruta, se sabe que determinan al menos unos doce genes.
- Genes que determinan el eje anteroposterior: determinan la formación de la cola y la cabeza.
Genes conmutadores maestros
Los genes conmutadores maestros se pueden definir como genes que actúan controlando la acción de otros genes disminuyendo el número de rutas de desarrollo alternativas que puede seguir la célula. Normalmente en un tiempo determinado de una célula cada punto tiene 2 acciones alternativas, esto es debido a los genes denominados genes conmutadores binarios. 2 ejemplos para entender el concepto son:
- El gen ced-9, relacionado con la apoptosis, controla la acción de los genes ced-3 y ced-4.
- El gen eyeless de Drosophila, si se expresa activa a otros genes que controlan el desarrollo y la diferenciación del ojo adulto, mientras que si no se expresa el organismo carecerá de ojo.
Estos genes son bastantes importantes para el desarrollo, ya que programan la expresión del genoma reduciendo bastante los caminos a seguir del zigoto para llegar al organismo pluricelular.
Genes maternos
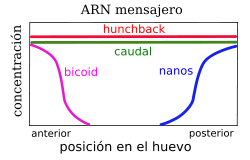
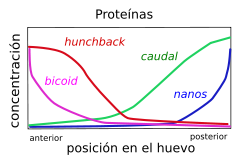 Distribución diferencial de los ARN mensajeros, a la izquierda, y proteínas, a la derecha, debicoid, nanos, hunchback y caudal en el embrión temprano de Drosophila (genes maternos)
Distribución diferencial de los ARN mensajeros, a la izquierda, y proteínas, a la derecha, debicoid, nanos, hunchback y caudal en el embrión temprano de Drosophila (genes maternos)Los primeros genes que actúan en el desarrollo son los genes maternos, por eso ese nombre, que se expresan durante la ovogénesis y actúan en la maduración del ovocito.
Estos genes se dividen en cuatro sistemas de genes (sistema anterior, sistema posterior, sistema terminal y sistema dorso-ventral) que definen los ejes del embrión. Cada sistema actúa localizando una señal dentro del huevo, llamada morfógeno. En la embriogénesis de Drosophila los diferentes sistemas determinan las siguientes partes:
- Sistema anterior: Se encarga de la zona anterior y por eso determina la formación de la cabeza y del tórax.
- Sistema posterior: Se encarga de la zona posterior por ese motivo determina la formación de los segmentos del abdomen.
- Sistema terminal: Se encarga de la zona final del cuerpo por eso determina la formación del acrón y del telson.
- Sistema dorso-ventral: Se encarga de las zonas dorso-ventrales y determina la formación de estructuras que se encuentran en dichas zonas, como por ejemplo, las alas y las patas.
Genes de segmentación
 Patrón bandeado de expresión de los genes even.skipped (parasegmentos impares) y fushi tarazu (parasegmentos pares).,[1]
Patrón bandeado de expresión de los genes even.skipped (parasegmentos impares) y fushi tarazu (parasegmentos pares).,[1]
Después de la acción de los genes maternos se activan los genes cigóticos o genes de segmentación que se expresan tras la fertilización. Estos genes se dividen en 3 clases que van actuar secuencialmente:
- Genes gap: son los primeros en actuar y dividen mediante la transcripción de estos genes el embrión en 4 regiones ancha. Dentro de estas regiones, las combinaciones diferentes de actividades génicas especificarán tanto el tipo de segmento que se formará como el orden correcto de los segmentos en cada estado del desarrollo, en Drosophila determinará los segmentos del cuerpo de la larva, pupa y del adulto.
- Genes de la regla par: son genes que se expresan después de los genes gap y dividen las amplias regiones establecidas por los genes gap en regiones con la anchura aproximada de un segmento, es decir, cada gen se expresa en un patrón de 7 bandas a lo largo del embrión, éstas bandas identifican parasegmentos impares (eve) y parasegmentos pares (Fushi tarazu) Por lo tanto, la expresión de estos genes establece los límites de los segmentos y el destino del desarrollo de las células dentro de cada segmento.
- Genes de la polaridad de los segmentos: estos genes se activan en una única banda de células dentro de cada segmento después de la acción de los genes de la regla par y se van a extender alrededor de la circunferencia del embrión. En total en Drosophila se van a expresar en 14 bandas a lo largo del embrión.
Genes homeóticos
 Estructura de los complejos homeóticos Antennapedia y bithorax en un cromosoma de Drosophila; el orden del 3' al 5' equivale al de expresión espacial, del extremo anterior al posterior, y al temporal, más precoz en el 3'.[1]
Estructura de los complejos homeóticos Antennapedia y bithorax en un cromosoma de Drosophila; el orden del 3' al 5' equivale al de expresión espacial, del extremo anterior al posterior, y al temporal, más precoz en el 3'.[1]Los genes homeóticos o Hox son los últimos genes cigóticos que actúan a lo largo del desarrollo y se activan tras el efecto de los genes de la polaridad de los segmentos, la expresión de estos genes determina las estructuras del adulto que se formarán en cada segmento corporal. En Drosophila se sabe que estos genes se agrupan en dos complejos ubicados en el cromosoma 3:
- Complejo antennapedia: este complejo controla la identidad de los segmentos de la cabeza y de los 2 primeros segmentos torácicos.
- Complejo Bithorax: controla la identidad del tercer segmento torácico y de los segmentos abdominales.
Se ha visto que el gen Antennapedia expresa la formación de una pata en el segundo segmento torácico, si se produce una mutación en la cabeza provocando que ese gen Antennapedia se exprese, ese animal tendrá, en vez de antenas, dos patas grandes y fuertes.
Hay 4 grupos de genes Hox en Humanos; A-B-C y D. Se extienden en regiones de entre 20 y 100Kb y contienen hasta 10 genes. Muchos se corresponden con genes de Drosophila y son las causantes de malformaciones hereditarias de las extremidades. También se han identificado cierto número de genes que controlan la expresión de los genes Hox, por ese motivo las mutaciones no afectan ni al número ni a la polaridad de los segmentos. Según los resultados de algunos experimentos se cree que los circuitos reguladores de los genes Hox no están ampliamente conservados en los sistemas animales.
Genes maternos ->(Activan)->Genes gap->(Activan)->Genes de regla par->(Activan)->Genes polaridad de segmento->(Activan)->Genes homeóticos
Interacciones entre células
En el desarrollo de los organismos pluricelulares las interacciones entre células influyen en los patrones de transcripción y en el destino del desarrollo de las células vecinas. Esto se consigue mediante sistemas de señalización en el desarrollo. En el desarrollo temprano, los vertebrados utilizan 5 sistemas de señalización, cuando empieza la organogénesis se añade 5 sistemas más de los ya utilizados. Estos sistemas actúan independientemente como redes coordinadas para enviar y recibir señales en el desarrollo que produce respuestas transcripcionales específicas, las redes establecen la formación de patrones y dirigen la diferenciación de los tejidos y órganos.
Un ejemplo de estos sistemas, son la ruta de señalización Notch. Este sistema de señalización actúa mediante contacto directo entre células para controlar el destino de desarrollo de las células que interactúan. El gen Notch codifica un receptor de señal transmembranal. Este receptor cuando se une a una proteína delta (formada mediante la expresión del gen delta) hay un cambio de conformación y se separa un pequeño trozo proteico que se una a una proteína citoplasmática codificada por el gen su(H), este complejo proteico formada en el citoplasma se dirige al núcleo para unirse a cofactores de transcripción que activan el mecanismo de transcripción de un conjunto de genes que controlan una ruta de desarrollo específica.
Apoptosis
El proceso de apoptosis consiste en la muerte celular programada que conlleva que el DNA celular sea degradado, encogiéndose la célula para luego ser fagocitada por células vecinas (macrófagos) pero sin que se salga el contenido de la célula. A diferencia de la necrosis que conlleva la muerte celular pero con salida del contenido al exterior tras hincharse demasiado, es lo que le ocurre a células dañadas. El proceso de apoptosis es importante para el desarrollo del organismo ya que es un mecanismo de sustitución de células viejas así como de mantenimiento del tamaño del individuo. El proceso está muy bien regulado tanto a nivel interior como exterior de la célula. Así se han descrito grupos de genes implicados en la regulación del proceso, muchos de los cuales codifican unos enzimas llamados caspasas, los cuales como función final llevan al corte de unas proteínas esenciales para la vida celular. En la mayoría de las células este proceso tiene que estar inhibido ya que muchas de las células están programadas para la muerte celular.
Agentes moleculares implicados en el desarrollo
- Morfógeno, término que en inglés se llama morphogen y que suele traducirse incorrectamente como ""morfogen", que es un término incorrecto que se refiere a algo inexistente. El término morfogén ha aparecido en numerosos artículos traducidos del inglés y ha inducido a muchas confusiones. Considero de gran relevancia informar a Wikipedia de la existencia de una definición en castellano de la palabra "Morfogén" e insisto: los morfogenes no existen. Lo único que existen son los morfógenos, que en inglés son Morphogens.
Genes
- Homeobox
- Genes homeóticos
- Genes HOX
- Genes PARAHOX
- Gen Pax
- El gen Tinman (Csh) controla el desarrollo del corazón tanto en Drosophila como en vertebrados.
Véase también
Referencias
- ↑ a b Wolpert, Lewis; Beddington, R.; Jessell, T.; Lawrence, P.; Meyerowitz, E. y Smith, W. (2002). Principles of Development (Second edition edición). Oxford: Oxford University Press. ISBN 0-19-879291-3.




Wikimedia foundation. 2010.