- Zingiberales
-
?Zingiberales

Hábito del árbol del viajero Clasificación científica Reino: Plantae Clados superiores: Archaeplastida
Viridiplantae
Streptophyta
Streptophytina
Embryophyta
Tracheophyta
Euphyllophyta
Lignophyta
SpermatophytaDivisión: Angiospermae Clase: Monocotyledoneae Subclase: Commelinidae Orden: Zingiberales Familias Zingiberales es el nombre de un taxón de plantas perteneciente a la categoría taxonómica de orden, utilizado por sistemas de clasificación modernos como el sistema de clasificación APG II del 2003[1] y el APWeb (2001 en adelante[2] ), en los que pertenece a las monocotiledóneas comelínidas, y es comprendido a su vez por 8 familias. Entre sus miembros se encuentran el jengibre, la flor mariposa, el ave del paraíso, el árbol del viajero, la achira y el banano. Zingiberales es un grupo monofilético bien definido, con muchas apomorfías. Son hierbas de tamaño grande, sin tallo aéreo salvo el que da las flores, con hojas claramente diferenciadas en pecíolo y lámina, muchas veces rotas entre las venas secundarias, las flores bilaterales (pero en clados más especializados sin plano de simetría), y las semillas con arilo. Las familias de este orden son bien conocidas por las modificaciones de su androceo, de forma que lo que parece ser el perianto son muchas veces estambres petaloideos. Los miembros de este orden están principalmente restringidos a los trópicos.
Contenido
Caracteres
Son hierbas bastante grandes, a veces de tamaño monstruoso (como Musa, Ravenala o Strelitzia). Falta el tallo aéreo salvo el que da las flores.
Las hojas están bien diferenciadas en pecíolo y lámina, poseen vena media, venas laterales con forma de S y venación fina transversal, venación pinada (más específicamente peni-paralela), están muchas veces rotas entre las venas secundarias, la postura de las hojas inmaduras en la yema (ptyxis) es supervoluta (en la que las dos mitades de la lámina están enrolladas en un tubo a lo largo del eje longitudinal, una mitad enrollada completamente dentro de la otra), el pecíolo con canales de aire alargados.
Inflorescencia con brácteas grandes y persistentes.
Flores grandes y de simetría bilateral (pero en clados más especializados sin plano de simetría), el perianto igualmente coloreado y el cáliz rodeando a la corola, 5 estambres con el miembro adaxial del verticilo interno no desarrollado, las anteras largas, el polen inaperturado, usualmente sin exina o con exina delgada y espinulosa (el polen no es resistente a la acetolisis), un ovario ínfero, el tegumento externo del óvulo de más de 3 células de espesor, la micropila biestomal, las células parietales de 1 (a veces 2) células de espesor, la presencia de "nucellar pad" (células epidérmicas del nucelo radialmente elongadas), el estilo largo, el estigma grande y húmedo.
Fruto capsular, semillas ariladas con perisperma, operculadas (el opérculo testal), presencia de collar micropilar (que se desarrolla desde el tegumento externo y forma una estructura anular en el perisperma que rodea al opérculo), la endotesta esclerotizada y silicifiada, el endosperma nuclear, la presencia de perisperma en sentido amplio, con almidón, el embrión como un "plug", el cotiledón no fotosintético, ligulado.
Presencia de "collar roots".
Con respecto a la anatomía, las cámaras de aire son diafragmadas en hojas y tallos, los haces vasculares del pecíolo están en arcos, los plástidos del tubo criboso también con granos de almidón, las ceras cuticulares en agregados tubulares, los vasos más o menos limitados a las raíces, y el pecíolo (y la vena media) con canales de aire alargados.
Deleción de 6 nucleótidos en el gen atpA .
También poseen células de sílice en el haz de la vaina, aunque no es una apomorfía de este orden solamente.
Las familias de este orden son bien conocidas por las modificaciones de su androceo, de forma que lo que parece ser el perianto son muchas veces estambres petaloideos, por lo que las flores suelen ser difíciles de interpretar y las piezas no siempre son lo que parecen ser.
Se puede ver una lista completa de caracteres del orden en la página del APWeb.[2]
Ecología
Zingiberales son hierbas gigantes con flores de simetría bilateral bastante complejas, y están principalmente restringidas a los trópicos. Givnish et al. (2005,[3] 2006[4] ) notan que la adquisición de venación reticulada, los propágulos dispersados por animales y la tolerancia a los hábitats sombreados están vinculados en este grupo.
McKenna y Farrell (2005,[5] 2006[6] ) discuten la diversificación de los escarabajos crisomélidos Cephaloleia en Zingiberales (también están presentes en otras comelínidas, y en Cyclanthaceae: ver también Staines 2004[7] ). Tanto la alimentación sobre los Zingiberales como la especialización de las larvas y particularmente adultos en las hojas jóvenes y enrolladas pueden haber aparecido una única vez, la asociación entre los escarabajos y Zingiberales pueden datar del Cretácico muy tardío (Wilf et al. 2000,[8] McKenna y Farrell 2006[6] ).
Abejas euglosinas son polinizadores importantes de los Zingiberales neotropicales (Zucchi et al. 1969, Williams 1982[9] ).
Diversidad
La diversidad taxonómica de las monocotiledóneas está presentada en detalle por Kubitzki (1998,[10] 2006[11] ).
A continuación se provee una lista de la diversidad de Zingiberales. Las descripciones son deliberadamente incompletas. Para más información siga los enlaces.
 Hábito e inflorescencia de Musella lasiocarpa.
Hábito e inflorescencia de Musella lasiocarpa.
 Frutos del banano Musa.
Frutos del banano Musa.Musáceas
Las musáceas están distribuidas en África, pasando por los Himalayas hasta el sur de Asia, las Filipinas y el norte de Australia. Son hierbas de tamaño considerable, las vainas de las hojas endurecidas pueden formar un pseudotallo. Las hojas son de disposición espiralada con pecíolo corto, en ellas las venas secundarias están más o menos perpendiculares a la vena media. Las brácteas de la inflorescencia son usualmente deciduas y la inflorescencia es un fascículo o una flor sin bráctea. Las flores tienen simetría bilateral, con 5 tépalos connados en la parte abaxial, y un único tépalo libre en la parte adaxial, profundamente cóncavo. Las inusuales flores son polinizadas por murciélagos o pájaros. El fruto es una baya (la banana). A pesar de su distribución restringida, las especies explotadas comercialmente para cultivar bananas fueron llevadas a todo el mundo, y se naturalizaron en Centroamérica.
El número de estambres de las musáceas es 6, el mayor número entre los Zingiberales, y es la razón por la que suelen ser vistas como una familia primitiva.
 Heliconia. Cada inflorescencia posee una bráctea conspicua y coloreada (en este caso colorada con el ápice amarillo), que rodea fascículos de flores de 5 estambres.
Heliconia. Cada inflorescencia posee una bráctea conspicua y coloreada (en este caso colorada con el ápice amarillo), que rodea fascículos de flores de 5 estambres.Heliconiáceas
Las heliconiáceas, con su único género Heliconia, son hierbas grandes distribuidas mayormente en América tropical, pero también presentes en Célebes a islas del Pacífico. En su aspecto son similares a Musaceae, pero con las hojas dísticas, y como Musaceae, poseen grandes brácteas por debajo de fascículos de flores tepaloideas, pero en Heliconiaceae las brácteas persisten y son fuertemente coloreadas. Además los pedicelos de las flores son elongados y poseen una única "hard stone" por lóculo, el fruto es una drupa.
Las flores de las heliconiáceas tienen 5 estambres.
 Inflorescencia (bráctea con flores) de Strelitzia. Las flores poseen 3 tépalos externos libres (aquí naranjas) y 3 internos soldados (azules, sólo se ven los 2 laterales).
Inflorescencia (bráctea con flores) de Strelitzia. Las flores poseen 3 tépalos externos libres (aquí naranjas) y 3 internos soldados (azules, sólo se ven los 2 laterales). Hábito del árbol del viajero.
Hábito del árbol del viajero.Strelitziáceas
Las hierbas de esta familia se pueden reconocer por sus hojas de disposición dística y largo pecíolo, y sus inflorescencias rodeadas de una bráctea grande, verde, resistente, de forma de bote. Las flores también son distintivas, los pétalos laterales abaxiales son más o menos connados. El fruto es una cápsula leñosa. Las semillas poseen arilos "fimbriados" de colores brillantes. En el árbol del viajero puede ocurrir, al igual que en los bananos, que las vainas de las hojas endurecidas formen un pseudotallo.
Los géneros de esta familia se distribuyen en la zona tropical de Sudamérica (el platanillo), en el sur de África (el ave del paraíso), y en Madagascar (el árbol del viajero). Son plantados en todo el mundo como ornamentales.
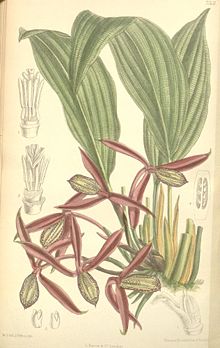 Ilustración de Orchidantha, el único género de las lowiáceas.
Ilustración de Orchidantha, el único género de las lowiáceas.Lowiáceas
Esta familia posee un único género, Orchidanta (el nombre de la familia no proviene del nombre del género, es una excepción), natural del sur de China y Borneo. Orchidantha significa "flor-orquídea", debido a su parecido con estas últimas: las flores poseen el pétalo medio desarrollada en un labelo abaxial bien desarrollado, y los dos pétalos laterales son pequeños. El sépalo medio es adaxial. Las hojas poseen venas longitudinales ascendentes y en apariencia bastante distantes, siendo algunas de ellas mucho más prominentes que las otras; y venas transversales muy desarrolladas que se observan con más facilidad en la cara abaxial de la hoja.
 La flor mariposa es muy utilizada como ornamental. Nótese la simetría fuertemente bilateral y el único estambre aferrado al estilo.
La flor mariposa es muy utilizada como ornamental. Nótese la simetría fuertemente bilateral y el único estambre aferrado al estilo.Jengibre, Flor Mariposa y afines
Las zingiberáceas son hierbas aromáticas picantes con aceites esenciales, las hojas se disponen de forma dística y se encuentran a lo largo de todo el tallo, las flores poseen simetría marcadamente bilateral y poseen cáliz, corola, 1 solo estambre aferrado al estilo, y usualmente varios estaminodios.
Varias especias pertenecen a la familia de las zingiberáceas, como el jengibre, la cúrcuma y el cardamomo, y también plantas de valor ornamental como la flor mariposa, que es la flor nacional de Cuba.
Son plantas tropicales, con la mayor diversidad en el sudeste asiático.
 Flor de Canna. Notar los estaminodios que parecen pétalos.
Flor de Canna. Notar los estaminodios que parecen pétalos.Achira y afines
La achira y otras especies emparentadas (género Canna, único de la familia de las canáceas) son originarias de las regiones tropicales y subtropicales de América.
La flor posee cáliz, corola, 1 solo estambre con 1 antera unilocular (es decir es una media antera), y lo que parecen ser pétalos son en realidad 3 o 4 estaminodios petaloideos. El estambre también es petaloideo y parecido a los estaminodios, y el estilo es aplanado y colorido.
Las canáceas son conocidas tanto como ornamentales como por su valor alimenticio.
Filogenia
La monofilia de este orden es sostenida por morfología (Dahlgren y Rasmussen 1983,[12] Kress 1990,[13] 1995,[14] Stevenson y Loconte 1995,[15] Stevenson et al. 2000,[16] Tomlinson 1962[17] ), por análisis moleculares de ADN hechos sobre un solo gen (Chase et al. 1993,[18] 1995b,[19] Smith et al. 1993,[20] Källersjö et al. 1998,[21] Soltis et al. 1997,[22] Davis et al. 2004,[23] Duvall et al. 1993[24] ), y sobre varios genes (Chase et al. 2000,[25] Soltis et al. 2000[26] ).
Zingiberales es hermano de Commelinales, ver una discusión del clado Zingiberales+Commelinales en Commelinidae.
Una filogenia de las principales familias de Zingiberales sería así (Judd et al. 2007, modificado de Kress 1990,[13] 1995,[14] Kress et al. 2001,[27] cet al. 2000,[26] 2005,[28] Chase et al. 2000[25] ):
Zingiberales Núcleo de los Zingiberales Las relaciones filogenéticas dentro del núcleo de las Zingiberales están muy bien comprendidas debido a cuidadosos estudios morfológicos y de ADN copy (Dahlgren y Rasmussen 1983,[12] Kress 1990,[13] 1995,[14] Kress et al. 2001,[27] Tomlinson 1962,[17] 1969a[29] ). Cannaceae, Marantaceae, Zingiberaceae, y Costaceae forman un clado basado en la reducción del androceo a un solo estambre funcional, la presencia de estaminodios vistosos, las semillas con más perisperma que endosperma, la falta de cristales de rafidio en los tejidos vegetativos, y las hojas que no se rompen fácilmente.
Dentro del núcleo, Marantaceae y Cannaceae se hipotetiza que son clados hermanos, como se evidencia por sus flores que no poseen plano de simetría, y su androceo con sólo la mitad de un estambre fértil, la otra mitad de este estambre siendo expandida y estaminodial.
Zingiberaceae y Costaceae constituyen un clado que está sostenido por la inusual característica de poseer un solo estambre funcional más o menos agarrado al estilo, una lígula en el ápice de la vaina, los sépalos connados, los estaminodios fusionados, y la reducción de dos de los tres estigmas.
Heliconiaceae, Strelitziaceae y Musaceae constituyen un complejo parafilético, y las relaciones entre estas tres familias aún no está clara. Heliconiaceae se hipotetiza que son el clado hermano del núcleo, como lo sostienen posibles sinapomorfías como un estambre medio externo estéril, pétalos connados, y detalles de la anatomía de la raíz.
Strelitziaceae puede ser hermano de las familias ya mencionadas, todas poseen hojas dísticas.
Musaceae, la familia del banano, probablemente representa el clado hermano de un clado que contiene a todo el resto de las familias del orden, esta familia ha retenido la condición plesiomórfica (ancestral) de hojas de disposición espiralada.
Taxonomía
- Introducción teórica en Taxonomía
8 familias, unas 1980 especies. Las familias más representadas son Cannaceae, Marantaceae y Zingiberaceae.
Según el APWeb,[2] sin cambios desde el APG II (2003[1] ), las familias son:
- Cannaceae familia de la achira
- Costaceae
- Heliconiaceae
- Lowiaceae
- Marantaceae
- Musaceae familia del banano
- Strelitziaceae familia del ave del paraíso y el árbol del viajero
- Zingiberaceae familia del jengibre y la flor mariposa.
Algunos autores (como Soltis et al. 2005) sostienen que las familias más pequeñas todavía se reconocen por razones históricas, y deberían ser integradas a sus familias más emparentadas.
Evolución
El grupo troncal Zingiberales data de unos 114 millones de años, la divergencia dentro del grupo de cerca de 88 millones de años (Janssen y Bremer 2004[30] ), según Bremer (2000[31] ) las fechas son de 84 y 62 millones de años, y según Wikström et al. (2001[32] ) son de 81-73 y 62-38 millones de años. Sin embargo, Kress y Specht (2005[33] ) encuentran la fecha del grupo troncal en unos 158 millones de años hasta el presente (127-121 millones de años en Kress y Specht 2006[34] ), la divergencia del grupo corona habría empezado hace 95 millones de años (144-106 según Kress y Specht 2006[34] ).
Todas las familias salvo Cannaceae y Marantaceae habrían divergido hace unos 60 millones de años (todas hace 86-74 millones de años según Kress y Specht 2006[34] ). Janssen y Bremer (2004[30] ) encontraron las fechas de divergencia dentro de Zingiberales muy amplias (las estimadas con la optimización DELTRAN fueron notablemente más jóvenes que las estimadas de las otras dos formas).
El fósil conocido es Spirematospermum, sin ubicar en familia, del Cretácico tardío.
Véase también
Bibliografía
- Judd, W. S.; C. S. Campbell, E. A. Kellogg, P. F. Stevens, M. J. Donoghue (2007). «Zingiberales». Plant Systematics: A Phylogenetic Approach, Third edition.. Sunderland, Massachusetts: Sinauer Associates. pp. 301-302. ISBN 978-0-87893-407-2.
- Simpson, Michael G. (2005). «Zingiberales». Plant Systematics.. Elsevier Inc.. pp. 192. ISBN 0-12-644460-9 ISBN 978-0-12-644460-5.
- Soltis, D. E.; Soltis, P. F., Endress, P. K., y Chase, M. W. (2005). «Zingiberales». Phylogeny and evolution of angiosperms.. Sunderland, MA: Sinauer Associates. pp. 112-113.
- Stevens, P. F. (2001 en adelante). «Zingiberales» (en inglés). Angiosperm Phylogeny Website (Versión 9, junio del 2008, y actualizado desde entonces). Consultado el 30 de enero del 2009.
Referencias citadas
- ↑ a b APG II (2003). «An Update of the Angiosperm Phylogeny Group Classification for the orders and families of flowering plants: APG II.» (pdf). Botanical Journal of the Linnean Society (141): pp. 399-436. http://www.blackwell-synergy.com/doi/pdf/10.1046/j.1095-8339.2003.t01-1-00158.x. Consultado el 06/06/2008.
- ↑ a b c Stevens, P. F. (2001 en adelante). «Angiosperm Phylogeny Website (Versión 9, junio del 2008, y actualizado desde entonces)» (en inglés). Consultado el 07-07-2008.
- ↑ Givnish, T. J.; 16 otros (2005). «Repeated evolution of net venation and fleshy fruits among monocots in shaded habitats confirms a priori predictions: evidence from an ndhF phylogeny.». Proc. Roy. Soc. London B (272): pp. 1481-1490.
- ↑ Givnish, T. J.; Pires, J. C., Graham, S. W., McPherson, M. A., Prince, L. M., Paterson, T. B., Rai, H. S., Roalson, E. H., Evans, T. M., Hahn, W. J., Millam, K. C., Meerow, A. W., Molvray, M., Kores, P. J., O'Brien, H. E., Hall, J. C., Kress, W. J., y Sytsma, K. J. (2006). «Phylogeny of the monocots based on the highly informative plastid gene ndhF : Evidence for widespread concerted convergence.». En Columbus, J. T., Friar, E. A., Porter, J. M., Prince, L. M., y Simpson, M. G.. Monocots: Comparative Biology and Evolution. Excluding Poales.. Claremont, Ca.: Rancho Santa Ana Botanical Garden. pp. 28-51. [Aliso 22: 28-51.]
- ↑ McKenna, D. D.; Farrell, B. D. (2005). «Molecular phylogenetics and evolution of host plant use in the neotropical rolled leaf 'hispine' beetle genus Cephaloleia (Chevrolat) (Chrysomelidae: Cassidinae).». Mol. Phyl. Evol. 37: pp. 117-131.
- ↑ a b McKenna, D. D.; Farrell, B. D. (2006). «Tropical forests are both evolutionary cradles and museums of leaf beetle diversity.». Proc. National Acad. Sci. U.S.A. 103: pp. 10947-10951.
- ↑ Staines, C. L. (2004). «Cassidinae (Coleoptera, Chrysomelidae) and Zingiberales: A review of the literature.». En olivet, P., Santiago-Blay, J. A., y Schmitt, M.. New Developments in the Biology of Chrysomelidae.. The Hague: SPB Academic Publishing. pp. 307-319.
- ↑ Wilf, P.; Labandeira, C. C., Kress, W. J., Staines, C. L., Windsor, D. M., Allen, A. L., y Johnson, K. R. (2000). «Timing the radiations of leaf beetles: Hispines on gingers from latest Cretaceous to Recent.». Science 289: pp. 291-294.
- ↑ Williams, N. H. (1982). «The biology of orchids and euglossine bees.». En Arditti, J.. Orchid Biology: Reviews and Perspectives, vol. 2.. Ithaca, N.Y.: Cornell University. pp. 119-171.
- ↑ Kubitzki, K., ed (1998). The families and genera of vascular plants, vol 3, Monocotyledons: Lilianae (except Orchidaceae).. Berlin: Springer-Verlag.
- ↑ Kubitzki, K., ed (2006). The families and genera of vascular plants, vol 4, Monocotyledons: Alismatanae and Commelinanae (except Gramineae).. Berlin: Springer-Verlag.
- ↑ a b Dahlgren, R. M. T.; Rasmussen, F. N. (1983). «Monocotyledon evolution: Characters and phylogenetic estimation.». Evol. Biol. (16): pp. 255-395.
- ↑ a b c Kress, W. J. (1990). «The phylogeny and classification of the Zingiberales.». Ann. Missouri Bot. Gard. (77): pp. 698-721.
- ↑ a b c Kress, W. J. (1995). «Phylogeny of the Zingiberanae: morphology and molecules.». En Rudall, P. J., Cribb, P. J., Cutler, D. F., y Humphries, C. J.. Monocotyledons: Systematics and evolution. (Royal Botanic Gardens edición). Kew. pp. 443-460.
- ↑ Stevenson, D. W.; Loconte, H. (1995). «Cladistic analysis of monocot families.». En Rudall, P. J., Cribb, P. J., Cutler, D. F.. Monocotyledons: Systematics and evolution. (Royal Botanic Gardens edición). Kew. pp. 543-578.
- ↑ Stevenson, D. W.; Davis, J. I., Freudenstein, J. V., Hardy, C. R., Simmons, M. P., y Specht, C. D. (2000). «A phylogenetic analysis of the monocotyledons based on morphological and molecular character sets, with comments on the placement of Acorus and Hydatellaceae.». En Wilson, K. L. y Morrison, D. A.. Monocots: Systematics and evolution. (CSIRO Publ. edición). Collingwood, Australia. pp. 17-24.
- ↑ a b Tomlinson, P. B. (1962). «Phylogeny of the Scitamineae: morphological and anatomical considerations.». Evolution (16): pp. 192-213.
- ↑ Chase, M. W.; Soltis, D. E., Olmstead, R. G., Morgan, D., Les, D. H., Mishler, B. D., Duvall, M. R., Price, R. A., Hills, H. G., Qiu, Y.-L., Kron, K. A., Rettig, J. H., Conti, E., Palmer, J. D., Manhart, J. R., Sytsma, K. J., Michaels, H. J., Kress, W. J., Karol, K. G., Clark, W. D., Hedrén, M., Gaut, B. S., Jansen, R. K., Kim, K.-J., Wimpee, C. F., Smith, J. F., Furnier, G. R., Strauss, S. H., Xiang, Q.-Y., Plunkett, G. M., Soltis, P. S., Swensen, S. M., Williams, S. E., Gadek, P. A., Quinn, C. J., Eguiarte, L. E., Golenberg, E., Learn, G. H., Jr., Graham, S. W., Barrett, S. C. H., Dayanandan, S., y Albert, V. A. (1993). «Phylogenetics of seed plants: An analysis of nucleotide sequences from the plastid gene rbcL.». Ann. Missouri Bot. Gard. 80: pp. 528-580.
- ↑ Chase, M. W.; Stevenson, D. W., Wilkin, P., y Rudall, P. J. (1995b). «Monocot systematics: A combined analysis.». En Rudall, P. J., Cribb, P. J., Cutler, D. F.. Monocotyledons: Systematics and evolution. (Royal Botanic Gardens edición). Kew. pp. 685-730.
- ↑ Smith, J. F.; Kress, W. J., Zimmer, E. A. (1993). «Phylogenetic analysis of the Zingiberales based on rbcL sequences.». Ann. Missouri Bot. Gard. (80): pp. 620-630.
- ↑ Källersjö M; JS Farris, MW Chase, B Bremer, MF Fay, ,CJ Humphries, G Petersen, O Seberg, y K Bremer (1998). «Simultaneous parsimony jacknife analysis of 2538 rbcL DNA sequences reveals support for major clades of green plants, land plants, and flowering plants.». Pl. Syst. Evol. (213): pp. 259-287.
- ↑ Soltis, D. E.; 15 autores más (1997). «Phylogenetic relationships among angiosperms inferred from 18S rDNA sequences.». Ann. Missouri Bot. Gard. (84): pp. 1-49.
- ↑ Davis, J. I.; Stevenson, D. W.; Petersen, G.; Seberg, O.; Campbell, L. M.; Freudenstein, J. V.; Goldman, D. H.; Hardy, C. R.; Michelangeli, F. A.; Simmons, M. P.; Specht, C. D.; Vergara-Silva, F.; Gandolfo, M. (2004). «A phylogeny of the monocots, as inferred from rbcL and atpA sequence variation, and a comparison of methods for calculating jacknife and bootstrap values.». Syst. Bot. (29): pp. 467-510. http://www.ingentaconnect.com/content/aspt/sb/2004/00000029/00000003/art00001. Consultado el 03/03/2008.
- ↑ Duvall, M. R.; Clegg, M. T., Chase, M. W., Clark, W. D., Kress, W. J., Eguiarte, L. E., Smith, J. F., Gaut, B. S., Zimmer, E. A., y Learns Jr., G. H. (1993). «Phylogenetic hypotheses for the monocotyledons constructed from rbcL sequence data.». Ann. Missouri Bot. Gard. (80): pp. 607-619. http://links.jstor.org/sici?sici=0026-6493(1993)80:3%3C607:PHFTMC%3E2.0.CO;2-5. Consultado el 25/02/2008.
- ↑ a b Chase, M. W.; Soltis, D. E., Soltis, P. S., Rudall, P. J., Fay, M. F., Hahn, W. H., Sullivan, S., Joseph, J., Molvray, M., Kores, P. J., Givnish, T. J., Sytsma, K. J., y Pires, J. C. (2000). «Higher-level systematics of the monocotyledons: An assessment of current knowledge and a new classification.». En Wilson, K. L. y Morrison, D. A.. Monocots: Systematics and evolution. (CSIRO Publ. edición). Collingwood, Australia. pp. 3-16.
- ↑ a b Soltis DE; PS Soltis, MW Chase, ME Mort, DC Albach, M Zanis, V Savolainen, WH Hahn, SB Hoot, MF Fay, M Axtell, SM Swensen, LM Prince, WJ Kress, KC Nixon, y JS Farris. (2000). «Angiosperm phylogeny inferred from 18S rDNA, rbcL, and atpB sequences.». Bot. J. Linn. Soc. (133): pp. 381-461. http://www.ingentaconnect.com/content/ap/bt/2000/00000133/00000004/art00380. Consultado el 03/03/2008.
- ↑ a b Kress, W. J.; Prince, L. M., Hahn, W. J., y Zimmer, E. A. (2001). «Unraveling the evolutionary radiation of the families of the Zingiberales using morphological and molecular evidence.». Syst. Biol. (50): pp. 926-944.
- ↑ Soltis, D. E.; Soltis, P. F., Endress, P. K., y Chase, M. W. (2005). Phylogeny and evolution of angiosperms.. Sunderland, MA: Sinauer Associates.
- ↑ Tomlinson, P. B. (1969). «Commelinales - Zingiberales.». En C. R. Metcalfe. Anatomy of the monocotyledons. vol. 3.. Oxford: Clarendon Press.
- ↑ a b Janssen, T.; Bremer, K. (2004). «The age of major monocot groups inferred from 800+ rbcL sequences.». Bot. J. Linnean Soc. 146: pp. 385-398.
- ↑ Bremer, K. (2000). «Early Cretaceous lineages of monocot flowering plants.». Proc. National Acad. Sci. U.S.A. 97: pp. 4707-4711.
- ↑ Wikström, N.; Savolainen, V., y Chase, M. W. (2001). «Evolution of the angiosperms: Calibrating the family tree.». Proc. Roy. Soc. London B 268: pp. 2211-2220.
- ↑ Kress, W. J.; Specht, C. D. (2005). «Between Cancer and Capricorn: Phylogeny, evolution and ecology of the primarily tropical Zingiberales.». Biol. Skr. 55: pp. 459-478. [Pp. 459-478, en Friis, I., y Balslev, H. (eds), Proceedings of a Symposium on Plant Diversity and Complexity Patterns - Local, Regional and Global Dimensions. Danish Academy of Sciences and Letters, Copenhagen.]
- ↑ a b c Kress, W. J.; Specht, C. D. (2006). «The evolutionary and biogeographic origin and diversification on the tropical monocot order Zingiberales.». En Columbus, J. T., Friar, E. A., Porter, J. M., Prince, L. M., y Simpson, M. G.. Monocots: Comparative Biology and Evolution. Excluding Poales. Rancho Santa Ana Botanical Garden, Claremont, Ca.. Amsterdam: Elsevier. pp. 621-632.[Aliso 22: 621-632.]
Enlaces externos
 Wikimedia Commons alberga contenido multimedia sobre Zingiberales. Commons
Wikimedia Commons alberga contenido multimedia sobre Zingiberales. Commons







Wikimedia foundation. 2010.