- Tallo
-
 Arquitectura del tallo.
Arquitectura del tallo.
En botánica, el tallo es el eje de la parte aérea de las cormófitas y es el órgano que sostiene a las hojas, flores y frutos. Sus funciones principales son las de sostén y de transporte de fotosintatos (carbohidratos y otros compuestos que se producen durante la fotosíntesis) entre las raíces y las hojas.[1]
Se diferencia de la raíz por la presencia de nudos en los que se insertan las yemas axilares y las hojas y por su geotropismo negativo, es decir, que crecen en contra de la fuerza de gravedad. Entre los cormófitos existen especies con un solo tallo cuyo vástago no se ramifica y plantas con muchos tallos (pluricaules) cuyo vástago se ramifica de diversos modos de acuerdo a la actividad de los meristemas.[2] [3]
Desde el punto de vista de la Anatomía, el tallo está constituido por tres sistemas de tejidos: el dérmico, el fundamental y el vascular o fascicular. Las variaciones en la estructura de los tallos de diferentes especies y de los taxones mayores se basan principalmente en las diferencias en la distribución relativa de los tejidos fundamental y vascular. El crecimiento en longitud del tallo se debe a la actividad de los meristemas apicales y al alargamiento subsecuente de los entrenudos y se denomina crecimiento primario. El crecimiento secundario se caracteriza por el aumento del grosor del tallo y es el resultado de la actividad de los denominados meristemas secundarios (cámbium y felógeno). Este tipo de crecimiento es característico de las gimnospermas y la mayoría de las eudicotiledóneas arbóreas y arbustivas y da como resultado la producción de madera.[4]
Los tallos se clasifican desde diversos puntos de vista, los cuales van desde la consistencia hasta las modificaciones que pudieran presentar para adaptarse a diferentes ambientes. Tal diversidad es la base de la gran cantidad de aplicaciones económicas que tienen los tallos, desde la alimenticia hasta las más variadas industrias.[5]
Contenido
Definición
Se define como todo órgano aéreo o subterráneo, verde o incoloro, derecho, rastrero o trepador, portador de hojas (sean éstas verdes, reducidas a escamas o cicatrices foliares), flores y frutos. A diferencia de las raíces, el tallo presenta geotropismo negativo, tiene nudos (lugares donde se originan las hojas) y entrenudos (regiones entre dos nudos consecutivos), yemas (áreas del tallo situadas justo por encima del punto de inserción de la hoja apical y axilares) y por lo común hojas bien desarrolladas. El tallo se forma a partir de la yema caulinar en la plántula o de las yemas secundarias en las ramificaciones. Estas ramificaciones pueden producirse mediante dos sistemas: monopódico o simpódico.[6]
Funciones
Es el eje de la planta que sostiene las hojas, órganos de asimilación con forma aplanada, las cuales se disponen de un modo favorable para captar la mayor cantidad de radiación solar con el mínimo sombreamiento mutuo (ver filotaxis). En las plantas que no presentan hojas identificables como tales, como en la mayoría de las cactáceas, el tallo se encarga de la fotosíntesis. En el momento de la reproducción, el tallo sostiene también las flores y los frutos. En muchas especies, el tallo es además uno de los órganos de reserva de agua y fotoasimilados, especialmente con antelación a la etapa reproductiva.
No obstante, la función principal del tallo es la de constituir la vía de circulación de agua entre las raíces y las hojas de las plantas. Puede tener muchos metros de altura, el tallo leñoso más largo que se conoce es el de la palmera trepadora Calamus manan de 185 m.[5]
El flujo de agua a través de la planta se realiza debido a las diferencias en el potencial hídrico entre la atmósfera y el suelo, siendo el xilema el tejido conductor. El flujo de agua en el xilema es un proceso físico, en donde la energía necesaria para que se lleve a cabo proviene de la transpiración del agua desde los estomas de las hojas hacia la atmósfera. Como consecuencia de tal transpiración, se produce una deficiencia de agua en las células del mesófilo de la hoja, el cual hace que el agua fluya desde las células más internas con un mayor potencial de agua. La deficiencia hídrica inicial se propaga sucesivamente hasta llegar a la altura de los conductos del xilema. La naturaleza capilar del xilema, las propiedades de cohesión de las moléculas de agua entre sí, la adhesión del agua a las paredes celulares y la tensión desarrollada por diferencias en el potencial hídrico originadas en la transpiración, permiten en conjunto, el movimiento de la columna de agua desde la raíz hasta las hojas.[7] [8] [9]
Morfología
El tallo, en general, es un órgano cilíndrico que posee puntos engrosados –nudos- sobre los que se desarrollan las hojas. A la porción de tallo situada entre dos nudos consecutivos se le denomina entrenudo. Presenta además una yema terminal en el extremo apical y varias yemas axilares que se diferencian en las axilas de las hojas.
Yemas
Una yema es el extremo joven de un vástago, y por lo tanto además del meristema apical, lleva hojas inmaduras o primordios foliares. La yema situada en el extremo del eje es la yema terminal, mientras que las que se encuentran en la unión de las hojas con el tallo son las yemas axilares. En ciertos casos es difícil distinguir las yemas del resto del tallo, especialmente cuando los primordios no están claramente agrupados, como sucede en el espárrago (Asparagus officinalis) y en especies de gran porte de las monocotiledóneas como Agave y Pandanus.
De acuerdo a su estructura se distinguen dos tipos de yemas, las yemas escamosas y las yemas desnudas.
El ápice de las yemas escamosas está protegido por hojas modificadas con aspecto escamoso, dispuestas apretadamente. Generalmente estas escamas, pérulas o tegmentos son oscuras y coriáceas, cumplen el rol de protección del ápice vegetativo. Las escamas, estrechamente aplicadas unas sobre otras y provistas de una gruesa cutícula, impiden la desecación de los tejidos embrionales durante el invierno, cuando la circulación de la savia es más lenta. Si se hace un corte longitudinal de la yema, se observa, por debajo de las escamas protectoras el ápice vegetativo, asiento del meristema apical del tallo y los primordios foliares. Cuando en la primavera el meristema inicia su actividad, las escamas caen, y los primordios foliares se desarrollan en hojas adultas. En Eucalyptus las yemas pueden tener hasta 50 pares de primordios foliares. Suelen tener pelos en abundancia que retienen el aire y constituyen un abrigo para el meristema, protegiéndolo de las variaciones térmicas bruscas. Las escamas pueden tener coléteres, estructuras glandulares secretoras de sustancias pegajosas como mucílagos y resinas, que contribuyen a la defensa contra la desecación al asegurar una mayor impermeabilidad; son comunes sobre todo en las plantas de deciduas. Las yemas desnudas están desprovistas de escamas protectoras y en este caso generalmente están protegidas por las hojas jóvenes. Estas yemas se presentan generalmente en plantas herbáceas.[10]
Las yemas axilares son generalmente únicas, es decir que en la axila de cada hoja nace una sola yema; en algunas especies, sin embargo, pueden presentarse yemas axilares múltiples o supletorias que originan flores, ramas, espinas o zarcillos. Según cómo estén dispuestas, hay dos tipos de yemas múltiples: seriales y colaterales. Las yemas seriales están situadas una por encima de la otra en la axila de la hoja tectriz, formando una fila vertical, como por ejemplo en el mburucuyá (Passiflora caerulea), en la madreselva (Lonicera japonica) y en la santa rita (Bougainvillea spectabilis), como así también están presentes en las bignoniáceas, las fabáceas y las rubiáceas). Las yemas colaterales o adyacentes se encuentran situadas una al lado de la otra en la axila de una misma hoja formando una fila horizontal. En el ajo (Allium sativum), cada diente es una yema axilar; en las inflorescencias del bananero (Musa) cada conjunto de yemas originará una "mano" de bananas. También están presentes en las aráceas y en las palmeras.[10]
El fenómeno de la caulifloria (árboles o arbustos que producen las flores en el tronco y en las ramas añosas) se debe al desarrollo tardío (años o décadas después) de yemas durmientes que quedan en la corteza del tallo.[10]
Dirección de crecimiento y simetría
Cuando el tallo se eleva verticalmente sobre el suelo, la planta se dice erecta y el eje ortótropo. En tal caso las ramas suelen desarrollarse radialmente alrededor del eje principal y cada rama crece horizontalmente y muestra simetría radial. En cambio, cuando el tallo principal crece en dirección horizontal su crecimiento se denomina plagiótropo. La planta en este caso se dice postrada o reptante, y su simetría suele ser dorsiventral.[5]
Sistemas de ramificación
 Ramificación dicotómica en Psilotum nudum.
Ramificación dicotómica en Psilotum nudum. Ramificación monopodial en Araucaria columnaris.
Ramificación monopodial en Araucaria columnaris. Ramificación simpodial en Foeniculum vulgare.
Ramificación simpodial en Foeniculum vulgare.Entre los cormófitos existen especies con un solo tallo, tales como el maíz Zea mays o la azucena Lilium longiflorum, cuyo vástago no se ramifica, excepto en la inflorescencia. Por otro lado, existen plantas con muchos tallos (pluricaules) cuyo vástago se ramifica. Hay dos tipos básicos de ramificación, la ramificación dicotómica y la ramificación lateral.[1]
En la ramificación dicotómica el ápice se divide en dos por división de la célula apical. Se trata de una ramificación típica de plantas muy primitivas, tales como los licopodios (Lycopodium y Psilotum). En la mayoría de los casos, las yemas se parten, originando siempre dos ramas; en otros, como por ejemplo en Lycopodium complanatum, la yema deja de crecer, y dos células próximas a ésta se diferencian, formando una rama nueva cada una. En las espermatófitas este tipo de ramificación es muy poco frecuente, sólo ha sido confirmada en algunas palmeras (tales como Nypa, Hyphaene y Chamaedorea), en ciertas cactáceas (Mammillaria) y en las flagelariáceas (lianas monocotiledóneas del Viejo Mundo).[2] [11] [3]
La ramificación lateral es el tipo dominante en las espermatófitas. En las espermatófitas, donde la ramificación es axilar (originada en el ancestro común a todas ellas), las ramas se originan en yemas axilares, a partir de la segunda o tercera hoja desde el ápice. En las pteridófitas es usual que las yemas se originen sobre la cara abaxial de las hojas o del pecíolo. Existen dos tipos básicos de ramificación lateral: la ramificación monopodial y la simpodial.
En la ramificación monopodial o sistema monopódico de ramificación, la yema apical crece y se desarrolla mucho más que las axilares, las que pueden incluso estar atrofiadas. La apariencia externa de las plantas que presentan este tipo de ramificación es la de un eje central robusto del cual salen unas ramitas muy delgadas, como es el caso de las coníferas. También puede observarse este tipo de ramificación en plantas herbáceas. Así, algunos rizomas crecen principalmente de manera monopódica: el eje principal se desarrolla en forma subterránea y más o menos rápidamente, y los vástagos aéreos se originan en yemas axilares. Debido a esta característica, las plantas con este tipo de rizoma tienen tendencia a ser muy invasoras, como el el caso del sorgo de Alepo (Sorghum halepense).
En el sistema simpódico de ramificación las ramas laterales se desarrollan más que el eje principal. El eje madre puede incluso interrumpir por completo su crecimiento, porque su yema apical queda en reposo o se transforma en una flor. Entonces una o varias yemas axilares, generalmente las superiores, se encargan de continuar el crecimiento y de formar nuevos brotes laterales, o sea, de proseguir su ramificación. Tanto las yemas apicales como axilares se desarrollan de la misma forma, sin que haya ningún tipo de dominancia. Aunque el eje central sigue siendo notorio, rápidamente las ramas alcanzan el mismo desarrollo. En regiones extratropicales, en las montañas de zonas tropicales y en climas tropicales secos, muchas plantas presentan crecimiento simpodial: la yema terminal muere pronto y es reemplazada por yemas laterales. La ramificación simpodial está ampliamente extendida en las dicotiledóneas herbáceas y se observa en prácticamente todas las monocotiledóneas.[12] [2] [11] [3]
Macroblasto y braquiblasto
En las plantas leñosas el crecimiento del tallo se efectúa en dos momentos y lugares distintos. En una primera fase, en la yema tiene lugar el crecimiento apical, por multiplicación de las células meristemáticas. De este modo se forman primordios foliares separados por entrenudos extremadamente cortos. Cuando la yema se va desarrollando, el tallo continúa elongándose por crecimiento intercalar de los entrenudos, primero los basales y luego los apicales. Este crecimiento ocurre por elongación celular más que por división. Según el grado de desarrollo de los entrenudos se distinguen dos tipos de ramas:[5]
- Macroblastos o ramas largas, son los ejes con un importante crecimiento de los entrenudos y, por lo tanto, presentan hojas bien separadas entre sí.
- Braquiblastos o ramas cortas, son los ejes con crecimiento internodal reducido y por lo tanto hojas muy próximas entre sí, dispuestas muchas veces en roseta. Las plantas brevicaules en roseta o rosuladas (mal llamadas acaules) son ejemplos de braquiblastos. Tal es el caso del repollo (Brassica oleracea var. capitata), la remolacha (Beta vulgaris), el rábano (Raphanus sativus), la lechuga (Lactuca sativa), especies de Agave y el llantén (Plantago).[5]
Ambos tipos de ramas pueden encontrarse en la misma planta. En los pinos (Pinus), por ejemplo, los macroblastos tienen hojas con forma de escamas en cuyas axilas se producen braquiblastos que llevan las hojas aciculares típicas de estas especies. En Ginkgo los braquiblastos llevan hojas flabeladas y estructuras reproductivas masculinas. En el peral (Pyrus communis) y en el manzano (Malus sylvestris) las flores nacen sobre braquiblastos.[5]
Tipos de tallos
Los tallos pueden clasificarse desde diversos puntos de vista, los cuales van desde la consistencia hasta las modificaciones adaptativas que pudieran presentar.[13] [2] [11] [3]
- Por su hábito
- Epígeos o aéreos: son todos aquellos tallos que crecen, como su nombre lo indica, por encima de la tierra. Incluyen los tallos normales con auténticas hojas y los estolones, siendo éstos brotes laterales más o menos delgados y generalmente muy largos (como es el ejemplo de la frutilla, Fragaria). De acuerdo con la dirección que sigue su crecimiento, los tallos aéreos pueden ser rectos o ascendentes si crecen de forma vertical, o rastreros si crecen de forma horizontal sobre la tierra.
- Hipógeos o subterráneos: son los tallos que crecen debajo de la tierra y presentan catáfilos (hojas rudimentarias). Dentro de este tipo de tallos se hallan los tubérculos, los rizomas y los bulbos, los cuales se describen a continuación:
- Rizomas: son tallos subterráneos de longitud y grosor variables, que crecen horizontalmente a profundidades diversas según las especies. Los nudos llevan hojas pequeñas, y cada año producen raíces que penetran en el suelo y tallos aéreos de vida corta —como es el caso del olluco— o simplemente un grupo de hojas formando un pseudotallo (como por ejemplo, el lirio, Iris germanica). Frecuentemente, los rizomas actúan como órganos de reserva de nutrientes.
- Tubérculos: son tallos que almacenan sustancias nutritivas. Tienen crecimiento limitado, no presentan habitualmente raíces y suelen durar un solo periodo vegetativo. En su superficie se observan catáfilos y yemas (denominadas “ojos”) y lenticelas.
- Bulbos: son tallos muy cortos y erectos, usualmente con forma de disco y con una yema terminal rodeada de varias hojas carnosas, densamente superpuestas, convertidas en órganos de reserva, llamadas catáfilos, que recubren el ápice y lo protegen.
- Cormos: se trata de tallos aplanados y de reserva con nudos y entrenudos muy cortos.
- Según su consistencia,
- Herbáceos: se trata de aquellos tallos que nunca desarrollan tejidos adultos o secundarios, por lo que tienen una consistencia suave y frágil.
- Escapo: es una tallo cuya única función es la de servir de sostén a las flores y, posteriormente, a los frutos. Una vez terminada su función, el escapo se seca y se cae. Puede presentar ramificaciones.
- Caña: es un tallo herbáceo macizo o hueco que no se ramifica. Es el tallo típico de las poáceas.
- Cálamos: son tallos aéreos, cilíndricos, que no presentan nudos, como por ejemplo, los tallos de los juncos (Juncus).
- Volubles: son tallos flexibles y enrollables en un soporte, como por ejemplo el del poroto (Phaseolus).
- Trepadores: son aquellos tallos que se fijan a un soporte mediante zarcillos, como por ejemplo los tallos de la vid (Vitis).
- Leñosos: son tallos rígidos y duros, sin color verde ya que no presentan clorofila.
- Arbustivos o Sufrútices: llegan a desarrollar tejidos secundarios, pero sólo en la región próxima a la base, manteniendo la parte superior de la planta siempre con tejidos jóvenes.
- Arbóreos: Son tallos que desarrollan tejidos secundarios por completo, limitando los primarios a las yemas tanto apicales como axilares. Son de consistencia dura, la que se debe a la acumulación de súber en ellos.
- Estípite: Son aquellos tallos monopodiales en los que la única yema que se desarrolla es la apical, quedando todas la demás atrofiadas. Es el caso de la mayoría de las palmeras.
- Carnosos o suculentos
- Según las modificaciones estructurales que presenten, los tallos pueden ser
- Zarcillo caulinar: es un tallo muy delgado que ha perdido la capacidad de formar hojas y flores. Su función es la de permitirle a la planta trepar o arrastrarse por diversas superficies.
- Espina caulinar: se trata de una rama modificada y muy lignificada que sirve como defensa contra los depredadores.
- Estolón: es un tallo cuya yema apical tiene la capacidad de formar raíces adventicias, lo que le permite formar una nueva planta.
- Tallos fotosintéticos:
Son aquellos tallos que han asumido las funciones de las hojas. Son propios de plantas que, por razones adaptativas, han dejado de formar hojas o estas se redujeron hasta volverse rudimentarias, o fueron modificadas hasta perder la capacidad fotosintética. Estos tallos, a su vez, pueden ser:
- platíclados, son tallos aplanados y fotosintéticos que puede tener la forma de una hoja, como los filodios o filóclados, los cuales son ramas aplanadas y de aspecto foliáceo, que puede llevar flores.
- cladodios, son tallos planos y suculentos, típicos de muchas especies de cactáceas (Opuntia por ejemplo), los que además de la función de fotosíntesis también están especializados en el almacenamiento de agua. A diferencia de los filodios, estos tallos fotosintéticos presentan crecimiento indeterminado.
Anatomía
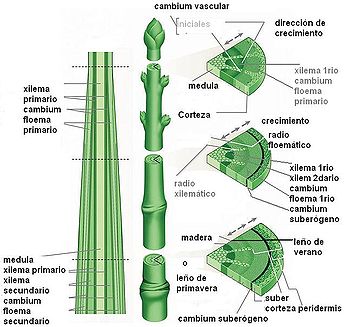 Esquema de las secciones longitudinal y transversal del tallo de una planta dicotiledónea, mostrando los meristemas y tejidos derivados que determinan el crecimiento primario y secundario del tallo.
Esquema de las secciones longitudinal y transversal del tallo de una planta dicotiledónea, mostrando los meristemas y tejidos derivados que determinan el crecimiento primario y secundario del tallo.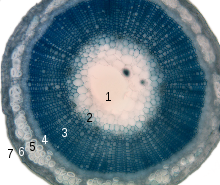 Corte transversal de un tallo de lino en el que se observa la estructura primaria del tallo. 1- médula 2-protoxilema 3-xilema primario 4-floema primario 5-fibras de esclerénquima 6-corteza 7-epidermis
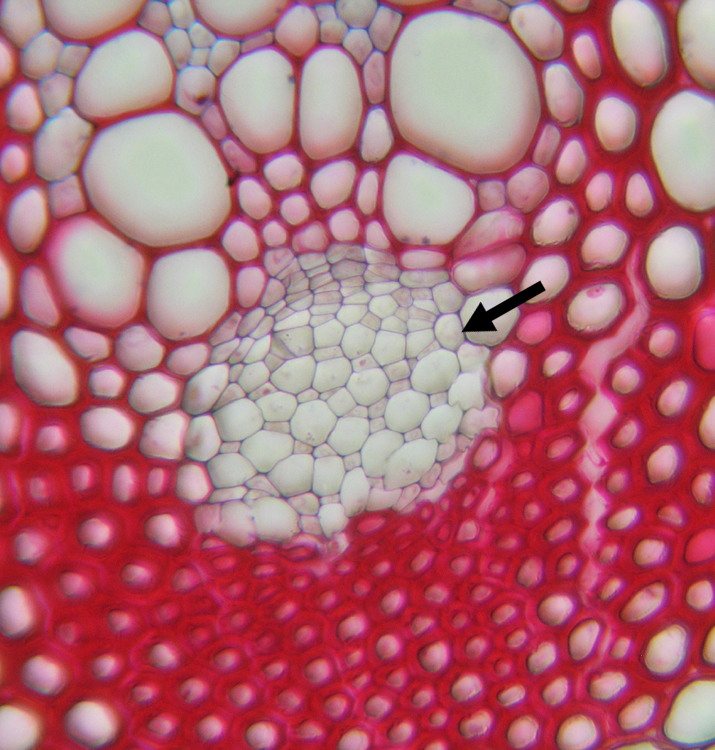
Corte transversal de un tallo de lino en el que se observa la estructura primaria del tallo. 1- médula 2-protoxilema 3-xilema primario 4-floema primario 5-fibras de esclerénquima 6-corteza 7-epidermis Detalle de un haz vascular en corte transversal de un tallo de Clematis. La flecha señala el floema, hacia arriba se observan los grandes vasos del xilema.
Detalle de un haz vascular en corte transversal de un tallo de Clematis. La flecha señala el floema, hacia arriba se observan los grandes vasos del xilema.El tallo está constituido por tres sistemas de tejidos: el dérmico, el fundamental y el vascular o fascicular. Las variaciones en la estructura de los tallos de diferentes especies y de los taxones mayores se basan principalmente en las diferencias en la distribución relativa de los tejidos fundamental y vascular. En las coníferas y dicotiledóneas el sistema vascular del entrenudo aparece como un cilindro hueco que delimita una región externa y una interna de tejido fundamental, la corteza y la médula, respectivamente. Las subdivisiones del sistema vascular, los haces vasculares, están separados unos de otros por paneles más o menos amplios de parénquima fundamental -llamado prenquima interfascicular- que interconecta la médula y la corteza. Este tejido se denomina interfascicular porque se encuentra entre los haces o fascículos. El parénquima interfascicular a menudo se llama también radio medular. Los tallos de muchos helechos, algunas dicotiledóneas herbáceas y la mayoría de las monocotiledóneas tienen una ordenación compleja de tejidos vasculares. Los haces vasculares pueden hallarse en más de un anillo o pueden aparecer dispersos por toda la sección transversal del tallo. La delimitación del tejido fundamental en corteza y médula es, en estos casos, menos precisa o no existe.[4] El crecimiento en longitud del tallo se debe a la actividad de los meristemas apicales y al alargamiento subsecuente de los entrenudos y se denomina crecimiento primario. El crecimiento secundario se caracteriza por el aumento del grosor del tallo y es el resultado de la actividad de los denominados meristemas secundarios (cámbium y felógeno). Este tipo de crecimiento es característico de las gimnospermas, la mayoría de las dicotiledóneas y algunas monocotiledónes.[4]
Estructura primaria del tallo
En un corte transversal, el tallo es generalmente cilíndrico, aunque a veces puede ser aplanado, triangular o cuadrangular. El sistema vascular primario consiste en un cilindro completo o bien en un sistema de haces vasculares discretos. La epidermis del tallo frecuentemente lleva estomas y tricomas, al igual que la de las hojas. La corteza es la región entre los tejidos vasculares y la epidermis. Esta corteza primaria o córtex está formada por tejidos fundamentales, en algunos casos sólo parénquima con función asimiladora o reservante, y en otras ocasiones también por colénquima, esclerénquima o ambos, con función mecánica. El conjunto entre la corteza y la región vascular se denomina región pericíclica, de donde pueden surgir las raíces adventicias. El cilindro central o estela es también llamado cilindro vascular primario, ya que está constituido por los tejidos conductores (floema y xilema primarios). Dichos tejidos en dicotiledóneas pueden formar un cilindro hueco (cuyo centro está ocupado por una médula parenquimatosa), o bien aparecer en forma de haces o cordones dispuestos en círculo alrededor de una médula parenquimatosa y separados entre sí por porciones parenquimáticas, llamadas radios medulares, que comunican la médula con la corteza. En monocotiledóneas, el floema y xilema primarios conforman haces que se distribuyen de manera dispersa y, por lo tanto, no se distingue médula ni radios medulares. En el tallo la disposición de los tejidos vasculares es generalmente colateral, con el floema primario dirigido hacia el exterior y el xilema primario hacia el interior, o sea, hacia el centro del tallo. Sin embargo, pueden existir otras disposiciones, por ejemplo, haces vasculares concéntricos. La médula es la región central del tallo, formada por tejido parenquimático, si bien en muchas especies puede reabsorberse y formar una cavidad central hueca.[14] [4]
Estela
El sistema formado por los tejidos vasculares en el eje de la planta, tallo y raíz, se denomina estela. El concepto de estela se elaboró para estudiar las relaciones y homologías en la estructura del vástago de diferentes grupos de plantas. Hay tres tipos básicos de estela según la distribución relativa del sistema vascular y el sistema fundamental de los ejes en estado primario de desarrollo: protostela, sifonostela y eustela, cada uno con variantes.[4]
La protostela es una columna sólida de tejidos vasculares ubicada en posición central. Es el tipo más simple y el más primitivo filogenéticamente, el cual se ha hallando en plantas fósiles como Psilophyton, una pteridófita de la era Paleozoica. La protostela se encuentra en algunas pteridófitas actuales, como Psilotum y Gleichenia, también en tallos de angiospermas acuáticas. Existen algunas variantes de la protostela. Así, cuando la columna de xilema tiene forma estrellada en el corte transversal, recibe el nombre de «actinostela». Cuando el xilema está fraccionado en varias placas, se habla de «plectostela».[15]
En la sifonostela, el sistema vascular tiene forma de tubo, envolviendo una médula parenquimática. No presenta lagunas foliares. Según la posición del floema, se distinguen dos tipos: la sifonostela anfifloica, con el floema por fuera y por dentro, sin lagunas foliares. Es exclusiva de las pteridofitas (Gleicheniaceae, Schizaeaceae, Marsileaceae) y la sifonostela ectofloica, en la que el floema se encuentra por fuera del xilema. Este último tipo se encuentra en los tallos de algunas pteridófitas y en raíces de Spermatophyta. La dictiostela es una variante de la sifonostela anfifloica con lagunas foliares muy grandes, que están superpuestas o solapadas. El sistema vascular, visto a lo largo, parece una red cilíndrica. En corte transversal cada segmento es un haz vascular concéntrico perifloemático. Se presenta, por ejemplo, en Polypodium, Microgramma, Dryopteris.
En la eustela el sistema vascular consta de haces vasculares organizados en simpodios, dispuestos alrededor de una médula. Las lagunas foliares pueden o no estar delimitadas (como en el caso de las gimnospermas y las dicotiledóneas), según que el sistema vascular sea cerrado o abierto. Los haces vasculares son abiertos, con cámbium fascicular, ya que la mayoría de estas plantas presenta crecimiento secundario. La atactostela es una variante de la eustela, característica de las monocotiledóneas, con haces vasculares colaterales o concéntricos esparcidos regularmente en todo el tallo debido a su recorrido longitudinal sinuoso. Los haces vasculares son cerrados, ya que este grupo de plantas no presenta crecimiento secundario.[15]
Actualmente se sabe que la estela de las plantas con semilla (epermatófitos) no ha evolucionado a partir de la sifonostela de los helechos, sino que se ha formado por fragmentación de una protostela, del tipo hallado en las primeras plantas con semilla (Progimnospermas).[15]
Estructura secundaria
La estructura secundaria del tallo se debe a la actividad de los meristemas laterales: cámbium vascular y felógeno. El primero actúa en el cilindro central, entre el floema y xilema primarios y el segundo se sitúa periféricamente, en la corteza o la epidermis. Ambos meristemas producen nuevas células en sentido radial, por lo que su actividad incrementa el grosor del tallo. Como consecuencia de la actividad cambial se origina floema secundario y células parenquimáticas (radios vasculares) hacia afuera, y xilema secundario y radios vasculares hacia adentro. El felógeno produce hacia el interior células parenquimáticas (felodermis) y hacia el exterior corcho (súber o felema). El conjunto de estas tres capas: felógeno, felodermis y corcho constituye la peridermis, la cual es la protección exterior del tallo cuando la epidermis se desgarra durante el crecimiento en grosor. Existen tallos que sólo producen una peridermis, pero también hay otros donde aparecen nuevas peridermis en zonas cada vez más internas, intercaladas con capas de tejidos aisladas por ellas. En esta situación se habla de ritidoma o corteza externa. La acción del cambium vascular genera más cantidad de tejido vascular que finalmente puede disponerse como un cilindro o quedar confinado a los haces vasculares existentes previamente. Cuando se forman cilindros, la cantidad de xilema secundario puede ser muy grande y se designa como madera o leño, y a los tallos con tal característica se les llama leñosos, en contraste con los tallos herbáceos que no producen tanta cantidad de xilema secundario (en cilindro o haces) o bien sólo tienen tejidos primarios. El floema secundario se produce en menor cantidad que el xilema y puede denominarse también líber, aunque este término se aplica igualmente al floema primario.[16] [17] [18] [4]
Madera, albura y duramen
En la mayoría de los árboles la parte interna del leño (xilema secundario originado por actividad del cámbium) cesa su actividad conductora de savia y sus células parenquimáticas mueren, debido fundamentalmente a la desintegración del protoplasma, al reforzamiento de las paredes con más lignina, a la acumulación en el lumen o impregnación de las paredes con sustancias orgánicas e inorgánicas (tales como taninos, aceites, gomas, resinas, colorantes, compuestos aromáticos, carbonato de calcio, silicio) y al bloqueo de los vasos con tílides. El leño que ha sufrido estos cambios es el duramen, inactivo y más oscuro. Cuanto mayor es la impregnación, mayor es la resistencia a los microorganismos que provocan la pudrición. La porción clara, externa, activa, con células vivas es la albura. La proporción albura-duramen varía entre las distintas especies, como también varía el grado de diferenciación entre ambas.[19]
Estructura del xilema secundario
La estructura característica del xilema secundario es la existencia de dos sistemas de elementos, que difieren en la orientación de sus células: uno es horizontal y el otro es vertical. El denominado sistema vertical, longitudinal o axial está compuesto por células o filas de células con el eje mayor orientado longitudinalmente, formado por elementos conductores no vivos y células parenquimáticas vivas. El sistema horizontal, transversal o radial está compuesto por hileras de células orientadas radialmente, formado por células vivas principalmente, las células parenquimáticas de los radios medulares. Las células vivas de los radios y del sistema axial se encuentran generalmente en conexión formando un sistema continuo.
 El cámbium produce elementos de mayor diámetro en primavera (leño temprano) y de menor diámetro y paredes más gruesas en invierno (leño tardío). De ese modo, la actividad de todo el año origina un anillo de crecimiento. Cada año se suma un nuevo anillo de crecimiento, los cuales pueden contarse a simple vista en el corte transversal de un tronco. En la imagen se observa un anillo de crecimiento en Robinia pseudoacacia, obsérvese la diferencia de diámetro de los poros entre el leño temprano y el leño tardío.
El cámbium produce elementos de mayor diámetro en primavera (leño temprano) y de menor diámetro y paredes más gruesas en invierno (leño tardío). De ese modo, la actividad de todo el año origina un anillo de crecimiento. Cada año se suma un nuevo anillo de crecimiento, los cuales pueden contarse a simple vista en el corte transversal de un tronco. En la imagen se observa un anillo de crecimiento en Robinia pseudoacacia, obsérvese la diferencia de diámetro de los poros entre el leño temprano y el leño tardío.El xilema secundario producido durante un período anual de crecimiento constituye una capa, que en corte transversal de tallo se llama anillo de crecimiento. Si se observa a simple vista tiene una parte clara, que es el leño temprano o de primavera, menos denso, con células de mayor diámetro y una parte oscura, que es el leño tardío, sus células son pequeñas y de paredes más gruesas. Esto ocurre generalmente en especies arbóreas que habitan en regiones de clima templado.[20]
En las gimnospermas el tejido leñoso está constituido principalmente por traqueidas, elementos imperforados con puntuaciones areoladas. En un anillo de crecimiento se distingue el leño temprano formado por traqueidas, las cuales son de mayor diámetro, y el leño tardío caracterizado por la presencia de fibrotraqueidas de paredes gruesas, lumen reducido y puntuaciones areoladas con abertura interna alargada. Las traqueidas y fibrotraqueidas miden entre 0,1 a 11 mm de longitud. Los radios medulares se hallan formados por una sola hilera de células, por lo que se denominan uniseriados. Pueden estar formados sólo por células parenquimáticas, como en los radios homocelulares, o también por traqueidas cortas, dispuestas en forma horizontal, como en los radios heterocelulares. El área de contacto entre un radio y las traqueidas del sistema vertical se denomina campo de cruzamiento. El tipo de puntuaciones, su número y su distribución son caracteres importantes para la identificación de las maderas de las diferentes especies de gimnospermas. Cuando presentan un canal resinífero los radios se denominan fusiformes.[21] [4]
El xilema secundario de las angiospermas es más complejo que el de las gimnospermas, razón por la cual se los describe como heteroxilo. Anatómicamente las diferencias son el resultado de la ordenación de los elementos de vasos, fibras y parénquima axial en un corte transversal. El elemento conductor de las dicotiledóneas son los miembros de vasos. En corte transversal se denominan "poros"; en las paredes laterales, los miembros de vaso poseen puntuaciones areoladas, en algunas maderas, estas puntuaciones presentan la abertura interna adornada, denominándose puntuaciones ornadas. La disposición de los poros en corte transversal se denomina porosidad. Si los vasos son de tamaño uniforme y se distribuyen más o menos homogéneamente a través del leño se dice que la porosidad es difusa, Ej.: Populus alba, Eucalyptus y Olea europaea. Si los vasos son de diferentes tamaños, y los formados en el leño temprano son notablemente mayores que los del final del anillo de crecimiento, la porosidad se conoce como circular o anular, ej.: Quercus. Los casos intermedios se denominan porosidad semianular. El arreglo de los vasos puede verse en corte transversal, variando en bandas tangenciales, cuando los vasos están ordenados perpendiculares a los radios, las bandas pueden ser rectas u onduladas; en un diseño radial o diagonal, o en un diseño dendrítico cuando su organización presenta un diseño con ramificaciones. Los radios pueden ser uniseriados o multiseriados, o sea, de varias hileras de espesor. Están formados por células parenquimáticas exclusivamente, con puntuaciones simples. El parénquima axial se dispone acompañando a los elementos verticales como las fibras y vasos. Las fibras son las células de sostén, a mayor cantidad de estas células, mayor es la dureza de la madera. Son células muertas, de paredes secundarias muy gruesas.[22]
Peridermis
La epidermis no acompaña el crecimiento en grosor del tallo secundario, siendo remplazada por la peridermis, tejido formado por el otro meristema secundario llamado felógeno, el que produce súber o corcho hacia fuera, y felodermis hacia el interior. Esta peridermis es gradualmente eliminada junto con las capas más viejas de floema, este conjunto de tejidos muertos es el ritidoma conocido vulgarmente como la corteza en los árboles. Un nuevo felógeno se forma cada cierto tiempo. En algunos árboles como el Quercus suber, del cual se extrae el corcho comercial, el felógeno dura toda la vida de la planta, produciendo súber de forma continua.[4]
El intercambio gaseoso se lleva a cabo por zonas llamadas lenticelas. Se forman normalmente donde se encontraban los estomas. En esta zona, el felógeno desarrolla un tejido de relleno, formado por células con abundantes espacios intercelulares.[23]
Crecimiento secundario anómalo
La anatomía del tallo descripta se denomina crecimiento secundario típico, y ocurre en las dicotiledóneas arbustivas y leñosas y en las gimnospermas. Algunas plantas como las trepadoras, lianas y enredaderas presentan variaciones de esta estructura, conocidas como crecimiento secundario anómalo. Algunos géneros de monocotiledóneas tales como Aloe, Yucca y la familia de las palmeras tienen crecimiento secundario anómalo, el cual difiere del crecimiento secundario típico en que nuevos vasos se forman en el margen del tallo, el xilema y el floema siguen presentándose como haces vasculares ya que no se forma un cilindro de xilema rodeado por uno de floema.[4]
Adaptaciones al ambiente
Adaptaciones al aprovisionamiento de agua
 Sección transversal del tallo de una planta acuática (Pontederia cordata). Obsérvese los grandes espacios intercelulares llenos de aire que aseguran la flotabilidad.
Sección transversal del tallo de una planta acuática (Pontederia cordata). Obsérvese los grandes espacios intercelulares llenos de aire que aseguran la flotabilidad. Los tallos aplanados, con función asimiladora, suculentos y con sus hojas transformadas en espinas son típicos de muchas cactáceas. Se denominan cladodios y representan una adaptación para evitar la pérdida de agua por transpiración y para acumular agua durante los períodos favorables.
Los tallos aplanados, con función asimiladora, suculentos y con sus hojas transformadas en espinas son típicos de muchas cactáceas. Se denominan cladodios y representan una adaptación para evitar la pérdida de agua por transpiración y para acumular agua durante los períodos favorables. Las estructuras que parecen ser hojas en Ruscus aculeatus son en realidad tallos aplanados que desempeñan la fotosíntesis y se denominan filóclados.
Las estructuras que parecen ser hojas en Ruscus aculeatus son en realidad tallos aplanados que desempeñan la fotosíntesis y se denominan filóclados.Los tallos de aquellas especies que viven sumergidas en el agua (hidrófitas) presentan una organización especial (hidromorfia) que les permite absorber directamente del agua, el dióxido de carbono y el oxígeno, tanto como las sales nutritivas. De hecho, algunas plantas acuáticas, tales como Ceratophyllum, Utricularia y Wolffia, carecen de raíces por esa razón. Las paredes celulares de las células epidérmicas de los tallos de estas plantas sólo desarrollan una cutícula muy delgada que apenas opone resistencia a la entrada de los gases, del agua y de las sales en disolución. El empuje que de abajo hacia arriba experimentan todos los objetos sumergidos hace innecesario en los tallos el tejido de sostén. En casi todas las plantas acuáticas, por otro lado, es notable el desarrollo de los espacios intercelulares. Los amplios conductos almacenan aire, lo que por un lado aumenta la flotabilidad y, por el otro, hace posible sobre todo una activa difusión de los gases en el interior de la planta.[3]
Las espinas son formaciones agudas, aleznadas, a veces ramificadas, provistas de tejido vascular, rígidas por ser ricas en tejidos de sostén. Las espinas pueden tener origen caulinar, es decir que son ramas reducidas a espinas, como sucede en Prunus spinosa y y otras especies como Gleditsia triacanthos. El tejido vascular de la espina es continuación del leño del tallo. Hydrolea spinosa, planta palustre típica de humedales, presenta espinas caulinares que a veces llevan hojas diminutas. En cambio los aguijones carecen de tejido vascular, y por ello son fáciles de arrancar. Los aguijones de Ceiba speciosa, el palo borracho, y de Fagara rhoifolia son emergencias formadas por tejidos corticales del tallo.
En los tallos suculentos de las cactáceas y de ciertas especies del género Euphorbia y de la familia de las asclepiadáceas la reducción de las hojas es extrema y las plantas son áfilas. La función asimiladora de las hojas es realizada por los tallos, cuya transformación se produce por reducción de ramas laterales, hojas reemplazadas por espinas y aumento de corteza para almacenar agua. Algunas cactáceas presentan tallos aplanados llamados cladodios, con aréolas (yemas axilares reducidas) en los nudos y estrechamientos en los puntos de ramificación. El caso extremo es el tallo esférico, con profundos pliegues o costillas que siguen los ortósticos. Estos tallos plegados pueden extenderse o contraerse según el parénquima acuífero esté más o menos repleto de agua. Las ramas que tienen crecimiento limitado, es decir que son braquiblastos, y presentan aspecto de hojas se llaman filóclados (por ejemplo, Ruscus aculeatus).[2] [11] [3]
Adaptaciones a períodos desfavorables para el crecimiento
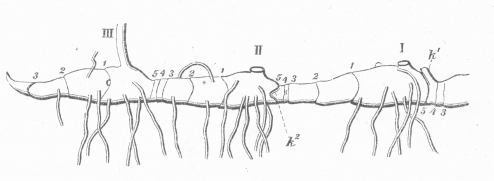
 Esquema del rizoma de Polygonatum verticillatum. I y II señalan el crecimiento de los dos años previos, III señala el crecimiento actual. Los números arábigos (1, 2, 3...) indican el nudo o lugar donde se hallaba cada catáfila u hoja modificada, los espacios entre los números son los entrenudos. K1, k2, indican la posición de las yemas axilares que están inactivas (dormidas).
Esquema del rizoma de Polygonatum verticillatum. I y II señalan el crecimiento de los dos años previos, III señala el crecimiento actual. Los números arábigos (1, 2, 3...) indican el nudo o lugar donde se hallaba cada catáfila u hoja modificada, los espacios entre los números son los entrenudos. K1, k2, indican la posición de las yemas axilares que están inactivas (dormidas).Las plantas han resuelto de maneras muy diversas el problema de la supervivencia durante épocas adversas, como son los inviernos muy fríos y los veranos excesivamente cálidos y secos. Así, las especies anuales completan su ciclo durante las estaciones favorables y transcurren como semillas las épocas desfavorables para el crecimiento. Las plantas bulbosas, en cambio, han desarrollado órganos subterráneos de reserva que les permiten sobrevivir durante las estaciones desfavorables en estado de reposo y reiniciar el crecimiento cuando las condiciones ambientales vuelven a ser favorables.[24] Las adaptaciones y las estrategias de las plantas bulbosas pueden satisfacer exigencias ecológicas muy diversas. Numerosos tulipanes (Tulipa) de origen asiático, por ejemplo, están adaptados a un clima continental extremo, con veranos secos y tórridos, inviernos helados y primaveras con breves aguaceros, período en el cual desarrollan su ciclo completo. Existen, por otra parte, muchas especies de sotobosque, como algunos crocos (Crocus), la escila (Scilla) y el diente de perro (Erythronium) que, gracias a sus reservas alimenticias, crecen muy rápido y cumplen su ciclo a principios de la primavera, antes de que las hojas de los árboles de hallan desarrollado y les quiten la luz del sol.[24] Muchas plantas bulbosas habitan comunidades adaptadas a incendios recurrentes durante la estación seca (ejemplo, varias especies de Iridaceae). En esos períodos, las plantas bulbosas se hallan en reposo y de ese modo sobreviven al calor del fuego. Los incendios limpian de vegetación la superficie, eliminando la competencia y, además, aportan nutrientes al suelo a través de las cenizas. Cuando las primeras lluvias caen, los bulbos, cormos y rizomas comienzan a brotar rápidamente, iniciando un nuevo período de crecimiento y desarrollo sostenido por las reservas acumuladas en sus tejidos durante la estación previa. Varias especies del género Cyrtanthus, por ejemplo, son reconocidas por su rápida capacidad de florecer luego de incendios naturales de pastizales, de ahí que varias de estas especies sean conocidas como "lirios de fuego". De hecho, ciertas especies tales como Cyrtanthus contractus, Cyrtanthus ventricosus y Cyrtanthus odorus, solo florecen luego de que se producen los incendios naturales.[25] El mayor número de especies de plantas bulbosas se encuentran en regiones del mundo con un clima mediterráneo, esto es, donde los inviernos son fríos y húmedos y los veranos son secos y cálidos, con una primavera corta. Las reservas acumuladas en estas plantas les permiten crecer rápidamente en la primavera, antes de que las hierbas anuales tengan tiempo de hacerlo. Cinco áreas en el mundo tienen este tipo de clima: la región del Mediterráneo, extendiéndose hacia el este hasta Asia Central; California; la región central de Chile, el extremo sur de Sudáfrica y el oeste y sur de Australia.
En los climas tropicales, donde existe una alternancia de estaciones secas y húmedas, las plantas bulbosas son también muy comunes. Finalmente, algunas especies bulbosas también proceden de regiones con lluvias estivales e inviernos secos. Una región con este tipo de clima y que es particularmente rica en especies bulbosas es la de las montañas Drakensberg en el noreste de la provincia de El Cabo en Sudáfrica.[26]
Las hierbas perennes o las plantas bianuales pierden los brotes foliosos aéreos para pasar la estación desfavorable; tienen yemas epígeas situadas a ras del suelo o yemas subterráneas. Para que estas yemas broten necesitan reservas elaboradas en el período favorable anterior, las cuales se almacenan en órganos como:
- Rizoma: son tallos subterráneos, generalmente de crecimiento horizontal, que pueden ramificarse simpodial o monopodialmente con menos frecuencia). Crecen indefinidamente, en el curso de los años mueren las partes más viejas pero cada año producen nuevos brotes, pueden cubrir grandes áreas. Sus ramas engrosadas suelen presentar entrenudos cortos, tienen catáfilos incoloros y membranáceas, raíces adventicias y yemas. Las plantas con rizomas son perennes, pierden sus partes aéreas en climas fríos conservando tan sólo el órgano subterráneo que almacena los nutrientes para la temporada siguiente. Frecuentemente las espermatófitas presentan rizomas simpodiales, en los que cada porción corresponde al desarrollo de yemas axilares sucesivas. La yema terminal de cada porción produce el brote epígeo, como por ejemplo en Sanseviera thyrsiflora y Paspalum nicorae. En los rizomas monopodiales, en cambio, la yema terminal continúa el crecimiento indefinido del rizoma, mientras las yemas axilares originan los brotes epígeos. Este tipo de rizomas es característico de muchas especies invasivas o malezas, tales como Sorghum halepense. Los rizomas se pueden dividir en trozos que contienen una yema al menos cada uno y plantar por separado. Es un sistema de reproducción vegetativa común a muchas plantas, como Achimenes, Canna, Zantedeschia, lirio y jengibre (Zingiber officinale).[27] [2] [11] [3]
 Joven tubérculo de papa (Solanum tuberosum) que se desarrolla en el extremo de un rizoma.
Joven tubérculo de papa (Solanum tuberosum) que se desarrolla en el extremo de un rizoma. Cormos de gladiolo (Gladiolus).
Cormos de gladiolo (Gladiolus). Bulbos de cebolla (Allium cepa).
Bulbos de cebolla (Allium cepa).- Tubérculo caulinar: son tallos con crecimiento limitado, epígeos o subterráneos. En el primer caso pueden originarse por fuerte engrosamiento primario o secundario del hipocótilo, o de uno o varios entrenudos. El colinabo, Brassica oleracea var. gongyloides, es un típico tubérculo caulinar epígeo. Otras especies con tubérculos derivados del hipocótilo son la violeta de los Alpes (Cyclamen sp.), el rábano (Raphanus sativus) y la remolacha roja (Beta vulgaris var. conditiva). La papa, Solanum tuberosum, presenta un tubérculo caulinar subterráneo o hipógeo formado a partir de los entrenudos apicales de rizomas. Esos rizomas presentan una zona meristemática sub-apical, de donde se originan los tubérculos mediante un engrosamiento radial, producto del alargamiento de las células parenquimáticas y la pérdida de la polaridad de las mismas. Durante la formación del tubérculo, el crecimiento longitudinal del estolón se detiene y las células parenquimáticas de la corteza, de la médula y de regiones perimedulares sufren divisiones y alargamiento. En tubérculos maduros, existen pocos elementos conductores y no hay un cámbium vascular continuo. Los tubérculos están cubiertos por una exodermis que aparece al romperse la epidermis que va engrosándose con el tiempo. Sobre su superficie existen "ojos", hundimientos para resguardar las yemas vegetativas que originan los tallos, que están dispuestos forma helicoidal. Además, hay orificios que permiten la respiración, llamados lenticelas. Las lenticelas son circulares y el número de las mismas varía por unidad de superficie, tamaño del tubérculo y condiciones ambientales.[28] Los tubérculos, en definitiva, están constituidos externamente por la peridermis, las lenticelas, los nudos, las yemas y, eventualmente, por un fragmento o una cicatriz proveniente de la unión con el rizoma del cual se originaron; internamente se distingue la corteza, el parénquima vascular de reserva, el anillo vascular y el tejido medular.[29]
- Cormo: es un tallo engrosado subterráneo, de base hinchada y crecimiento vertical que contiene nudos y abultamientos de los que salen yemas. Esta recubierto por capas de hojas secas, a modo de túnicas superpuestas llamadas túnicas, las que varían mucho entre especies. Así, hay especies con túnicas papiráceas, fibrosas o con la textura de la cáscara de un huevo. Además, las especies con túnicas fibrosas pueden tener las fibras de las túnicas dispuestas en forma paralela, o bien, reticulada. En la parte inferior del cormo se producen pequeños cormos nuevos que servirán para la reproducción de nuevas plantas. Al igual que los bulbos y los rizomas, estos órganos son acumuladores de sustancias nutritivas constituidos por células parenquimáticas. Las plantas que presentan cormos son plantas perennes que pierden sus partes aéreas en climas fríos durante la época invernal, conservando únicamente su parte subterránea. Esta capacidad para almacenar nutrientes constituye un método de supervivencia en caso de condiciones adversas, como una prolongada sequía o una temporada estival demasiado calurosa.[27]
- Bulbo: la función reservante es ejercida principalmente por hojas engrosadas y carnosas. El tallo generalmente es subterráneo, muy intensamente acortado, con hojas engrosadas y carnosas. En la cebolla, Allium cepa, planta bienal, cuando la semilla germina se forma un tallo corto llamado platillo que lleva las hojas cilíndricas dispuestas en forma concéntrica. Las vainas foliares se ensanchan, llenándose de sustancias de reserva. Las más externas no engruesan, se secan constituyendo túnicas de protección. En invierno los limbos foliares mueren y queda el bulbo bajo tierra. Al año siguiente la yema apical desarrolla el tallo florífero, utilizando las reservan acumuladas, el bulbo se consume totalmente y no es reemplazado, es decir que la planta muere. En el ajo, Allium sativum, todas las túnicas son delgadas y papiráceas. En la axila de cada túnica se desarrollan dos a cinco yemas colaterales que formarán bulbillos, los "dientes de ajo", cada uno con una sola túnica carnosa alrededor de la yema terminal. Cada bulbillo puesto en tierra dará un brote epígeo, y las yemas ubicadas en la axila de sus túnicas protectoras formarán nuevamente bulbillos. En Tulipa las hojas engrosadas son catáfilos, que nunca presentan partes aéreas, los más externos de protección y los más internos reservantes; se observa una yema terminal y una yema axilar en la axila del catáfilo más interno. En primavera la yema terminal originará el tallo florífero, utilizando las reservas del bulbo. Las hojas persisten después de la floración y en verano las reservas se acumulan en los catáfilos de la yema axilar, formando un nuevo bulbo (bulbo simpodial) que repetirá el proceso. Una vez que se forma el nuevo bulbo el tallo aéreo y el disco viejo se secan.[27]
Adaptaciones al aprovechamiento de la luz

 Espinas caulinares en Gleditsia triacanthos.
Espinas caulinares en Gleditsia triacanthos.Las trepadoras elevan en poco tiempo sus hojas por encima de la sombra de los árboles trepando por encima de otros vegetales, o también sobre rocas o muros. Existen dos tipos básicos de plantas trepadoras de acuerdo a sus tallos:[2] [11] [3] [27]
- Enredaderas: son plantas trepadoras herbáceas o subleñosas, con tallos delgados, primordialmente crecen en hábitats modificados o en bordes de bosques, como por ejemplo Cissampelos pareira, Muehlenbeckia sagittifolia y Oxypetalum macrolepis.
- Lianas o bejucos: son trepadoras leñosas, de tallos gruesos, típicas de bosques y selvas maduros. Muchas lianas presentan crecimiento secundario anómalo. Una leguminosa, Bauhinia macrostachya, conocida como "escalera de mono", tiene el tallo acintado, más delgado en el centro que en los márgenes, resistente a la torsión y a la flexión. En las bignoniáceas la disposición particular de los tejidos vasculares del tallo le otorga flexibilidad. En las sapindáceas, es frecuente el tallo fasciculado, resultado una estructura retorcida parecida a sogas.
- Zarcillos, órganos filiformes o ramificados que tienen la capacidad de rodear los soportes y fijarse a ellos por su intensa excitabilidad al contacto. Su origen es caulinar en Vitis vinifera, Passiflora, y foliar en las leguminosas y en las bignoniáceas. En Parthenocissus los zarcillos caulinares son ramificados y terminan en ventosas.
- Movimientos de circumnutación de los tallos con largos entrenudos. Se presentan en plantas volubles como Phaseolus, Aristolochia y Convolvulus. En muchos tallos se presenta además una corteza gruesa y áspera que probablemente evita el deslizamiento, como por ejemplo en las familias Aristolochiaceae, Apocynaceae y Cucurbitaceae.
- Aguijones y espinas. Los primeros se presentan en especies trepadoras de Rosa. Bougainvillea spectabilis, la "Santa Rita" y varias especies de Mimosa presentan espinas caulinares. Desmoncus, es otro ejemplo y representa la única palmera neotropical trepadora con tallos espinosos.
Importancia económica
 Los tallos del espárrago son comestibles.
Los tallos del espárrago son comestibles. La canela proviene de la corteza del tronco de un árbol.
La canela proviene de la corteza del tronco de un árbol.Existen miles de especies cuyos tallos presentan importancia económica. Muchos tallos proveen alimento para los seres humanos, tales como la papa (Solanum tuberosum) o el taro (Colocasia esculenta). La caña de azúcar (Saccharum) se utiliza en la industria no solo como fuente de sacarosa sino para producir alcohol. Varios vegetales de uso culinario son tallos, tales como el espárrago, los brotes de bambú, los cladodios jóvenes de Opuntia para preparar nopalitos, el colirábano (Brassica oleracea gongyloides), el corazón o cogollo de los palmitos (Chamaerops humilis). La canela es una especia que se extrae de la corteza del tronco de un árbol (Cinnamomum verum). La goma arábiga es un aditivo alimentario que se obtiene de los troncos de Acacia senegal. El chicle, ingrediente principal de la goma de mascar se obtiene de los troncos de Manilkara zapota. A partir de los tallos de muchas especies se obtienen también medicinas, tales como la quinina a partir de la corteza del quino (Cinchona ledgeriana), el alcanfor a partir de la madera destilada de Cinnamomum camphora, el relajante muscular llamado curare extraído de la corteza de variadas especies de enredaderas tropicales, entre ellas, Strychnos toxifera[2] [3] y el ácido salicílico de la corteza de varias especies de sauce (Salix).[30]
La madera se emplea de miles de maneras, por ejemplo, en la construcción de edificios, muebles, barcos, aeroplanos, piezas de automóviles, instrumentos musicales, equipos deportivos, durmientes de ferrocarril, postes de electricidad, postes para cercas, postes, palillos, fósforos, contrachapado, ataúdes, toneles, juguetes, mangos de herramientas, marcos de cuadros, chapas de madera, carbón vegetal y leña. La pasta de celulosa se utiliza ampliamente para fabricar papel, cartón, esponjas de celulosa, papel celofán y algunos plásticos y textiles importantes, tales como el acetato de celulosa y el rayón. Los árboles de crecimiento lento proveen, en general, maderas duras, de mejor calidad, que se emplean usualmente en la industria del mueble. Los árboles de crecimiento rápido dan maderas blandas que se usan principalmente para la fabricación de papel. Entre los árboles de maderas nobles se encuentran la caoba, el ébano, la teca, y el palo rosa empleados para hacer muebles de gran calidad, y entre los de madera blanda, las coníferas son muy comunes, como el alerce, el pino, el abeto y otros como el álamo, la haya, el abedul y el roble, de aplicación en la industria de la construcción. Los tallos de bambú también tienen cientos de usos, incluyendo la fabricación de papel, muebles, embarcaciones, instrumentos musicales, cañas de pescar, conductos de agua y andamios. Los troncos de las palmeras y de los helechos son a menudo utilizados para la construcción.[2] [3]
Otros productos de uso industrial también se extraen de tallos. Los taninos utilizado para el curtido del cuero se obtienen de la madera de ciertos árboles, como el quebracho (Schinopsis balansae). El corcho se obtiene de la corteza del alcornoque (Quercus suber). El látex se obtiene de los troncos de Hevea brasiliensis. El ratán, utilizado para la confección de muebles y cestas, está hecho de los tallos de palmas tropicales. Las fibras para la industria textil se obtienen de los tallos de muchas especies, entre las que se incluyen el lino, el cáñamo, el yute y el ramio. Los primeros documentos escritos fueron obtenidos por los antiguos egipcios a partir de tallos de papiro.[2] [3]
Muchas plantas ornamentales se cultivan por sus atractivos troncos, como por ejemplo Betula papyrifera por su corteza blanca y Acer griseum por sus troncos de color rojo, como así también los tallos y ramas retorcidas de algunas especies de sauces (Salix matsudana y Salix erythroflexuosa).[31]
Referencias
- ↑ a b Font Quer, P. (1982). Diccionario de Botánica. 8ª reimpresión. Barcelona: Editorial Labor, S. A. ISBN 84-335-5804-8.
- ↑ a b c d e f g h i j Valla, J.J. 2005. Botánica: morfología de las plantas superiores. Buenos Aires, Hemisferio Sur. ISBN 950-504-378-3
- ↑ a b c d e f g h i j k Strassburger, E. 1994. Tratado de Botánica. 8va. edición. Omega, Barcelona, 1088 p. ISBN 84-7102-990-1
- ↑ a b c d e f g h i ESAU, K. 1988. Anatomía de las plantas con semilla. Ed. Hemisferio Sur. ISBN 950-004-233-9
- ↑ a b c d e f González, A.M. «Organización del Cuerpo de las Plantas: diferenciación morfológica del tallo, hoja y raíz» (en español). Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Consultado el 6 de octubre de 2009.
- ↑ ACEDO C. 2004-2008. Definición de tallo. Botánica en la web. Universidad de León. Consultado el 26 de octubre de 2009.
- ↑ Biología Virtual.Morfología, Anatomía y Fisiología del tallo. Universidad Nacional de Colombia. Consultado el 18 de octubre de 2009.
- ↑ Loomis R.S. y Connor D.J. 1992. Crop Ecology. Ed. Cambridge University Press
- ↑ Salisbury F.B. y Ross, C.W. 1985. Plant physiology. Wadsworth Publ. Co.
- ↑ a b c González, A.M. «Yemas» (en español). Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Consultado el 12 de octubre de 2009.
- ↑ a b c d e f Gola, G., Negri, G. y Cappeletti, C. 1965. Tratado de Botánica. 2da. edición. Editorial Labor S.A., Barcelona, 1110 p.
- ↑ González, A.M. «Organización del Cuerpo de las Plantas Sistemas de ramificación» (en español). Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Consultado el 6 de octubre de 2009.
- ↑ ACEDO C. 2004-2008 Tipos de tallos. Botánica en la web. Consultado el 28 de octubre de 2009.
- ↑ Rudall, P. 2007. Anatomy of flowering plants: an introduction to structure and development. Cambridge University Press.
- ↑ a b c González, A.M. «Estructura primaria del tallo. 17.7. Concepto de estela» (en español). Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Consultado el 6 de octubre de 2009.
- ↑ CRONQUIST, A. Introducción a la Botánica. 3ª. Ed. CECSA. México. Cap. 25.
- ↑ ESAU, K. 1976. Anatomía Vegetal 3ª Ed. Omega. Barcelona. Cap. 15.
- ↑ LUTGE, U.; M. Kluge y G. Bauer. 1993. Botánica. McGraw-Hill Interamericana. Cap. 22.
- ↑ Raisman, J.S. y González, A.M. Maderas: albura y duramen. Hipertextos del área de Biología. Universidad Nacional del Nordeste. Consultado el 28 de octubre de 2009.
- ↑ Raisman, J.S. y González, A.M. Maderas: estructura del xilema secundario . Hipertextos del área de Biología. Universidad Nacional del Nordeste. Consultado el 28 de octubre de 2009.
- ↑ Raisman, J.S. y González, A.M. Maderas: gimnospermas. Hipertextos del área de Biología. Universidad Nacional del Nordeste. Consultado el 28 de octubre de 2009.
- ↑ Raisman, J.S. y González, A.M. Maderas: angiospermas. Hipertextos del área de Biología. Universidad Nacional del Nordeste. Consultado el 28 de octubre de 2009.
- ↑ Raisman, J.S. y González, A.M. Maderas: peridermis. Hipertextos del área de Biología. Universidad Nacional del Nordeste. Consultado el 28 de octubre de 2009.
- ↑ a b Rosella Rossi. 1990. Guía de Bulbos. Grijalbo. Barcelona.
- ↑ http://www.plantzafrica.com/plantcd/cyrtanthus.htm Cyrtanthus, por G. Duncan, Jardín Botánico Nacional de Kirstenbosch, Sudáfrica.]
- ↑ Phillips, R. & Rix, M. 1989. Bulbs. Pan Books Ltd.]
- ↑ a b c d González, A.M. «Adaptaciones del cormo» (en español). Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Consultado el 12 de octubre de 2009.
- ↑ Xin, E., D. Vreugdenhil y A. M. Lammeren (1998). «Cell division and cell enlargement during potatoes tuber formation». Journal of Experimental Botany (49). 573-582.
- ↑ Faiguenbaum M, H., Zunino, P. «Biología de Cultivos Anuales, Papa. Tubérculo». Facultad de Agronomía e Ingeniería Forestal. Pontificia Universidad Católica de Chile.
- ↑ Jeffreys, Diarmuid (en inglés). Aspirin: The Remarkable Story of a Wonder Drug. Bloomsbury. ISBN 1582343861. http://www.amazon.com/Aspirin-Remarkable-Story-Wonder-Drug/dp/1582343861. Consultado el 21 de noviembre de 2008.
- ↑ Raven, Peter H., Ray Franklin Evert, y Helena Curtis. 1981. Biology of plants. New York, N.Y.: Worth Publishers.ISBN 0-87901-132-7
Bibliografía
- González, A.M. «Morfología de Plantas Vasculares» (en español). Argentina: Universidad Nacional del Nordeste. Consultado el 10 de abril de 2009.
- Strassburger, E. 1994. Tratado de Botánica. 8va. edición. Omega, Barcelona, 1088 p. ISBN 84-7102-990-1
- Valla, J.J. 2005. Botánica: morfología de las plantas superiores. Buenos Aires, Hemisferio Sur. ISBN 950-504-378-3
Enlaces externos
 Wikimedia Commons alberga contenido multimedia sobre Tallo. Commons
Wikimedia Commons alberga contenido multimedia sobre Tallo. Commons
Categorías:- Términos botánicos
- Morfología del tallo















Wikimedia foundation. 2010.