- Fisiología y ecología sensorial en Triatominae
-
Los triatominos perciben los olores con sus antenas. La información contenida en las sustancias volátiles provenientes de los vertebrados y de sus congéneres es esencial para la supervivencia de estos insectos chupasangre y transmisores de la enfermedad de Chagas.
 Rhodnius prolixus caminando en Servosfera (ver Videos).
Rhodnius prolixus caminando en Servosfera (ver Videos).
Contenido
Importancia
Los estímulos visuales, químicos, acústicos, vibratorios, térmicos y táctiles constituyen todos ellos pistas de alta relevancia para la orientación de estos insectos. Sin embargo, los compuestos volátiles son particularmente importantes para los triatominos por dos razones, a) la mayor parte de estas especies buscan huéspedes durante la noche cuando la luz es escaza y b) la información presente en los olores puede viajar largas distancias pues con frecuencia son transportados por corrientes de aire, pasando obstáculos que otro tipo de estímulos no podrían salvar.
La olfación en los triatominos ha sido estudiada desde hace poco más de un siglo con el incentivo de desarrollar aplicaciones útiles para el control de estos insectos hematófagos transmisores del parásito Trypanosoma cruzi. Los hallazgos realizados sobre este particular han arrojado luz sobre aspectos sensoriales básicos que abarcan a los artrópodos chupasangre en general, otros invertebrados, e incluso a los vertebrados.
Oliendo con las antenas
 Antena de ninfa de Triatoma infestans.
Antena de ninfa de Triatoma infestans.Las antenas además de servir para tocar y degustar son, entre los artrópodos, los órganos que por excelencia permiten percibir olores. Los triatominos poseen antenas de 4 segmentos a ambos lados de la cabeza a una distancia variable de los ojos en la región anterior a estos: un segmento basal o escapo, le sigue un pedicelo y un flagelo compuesto: flagelo distal y flagelo próximal (también llamados flagelómeros). Los triatominos, como otros artrópodos, baten el aire con sus antenas para "capturar" las moléculas presentes en el aire. Cada antena contiene cientos de células olfativas con receptores para olores, pero actualmente solo se han descubierto unas pocas. Estos receptores son estructuras proteicas ubicadas en la membrana de las terminaciones nerviosas de las células olfativas. Estas últimas a su vez están contenidas en pelos sensoriales o tricobotrios (sensillas en inglés) que cubren las antenas.[1] Los olores entran a los pelos olfativos a través de poros y luego son llevados a las receptores de la membrana por proteínas adherentes ("odorant-binding proteins" en inglés). Estas proteínas solubles están contenidas en la linfa de los pelos sensoriales. Ciertos olores activan los receptores membranales y estos desencadenan una cascada de señalización que produce estímulos eléctricos los cuales viajan a los lóbulos antenales de los triatominos. Las células nerviosas que tienen receptores para un determinado olor generalmente convergen en glomérulos que se encuentran en los lóbulos antenales; los triatominos poseen aproximadamente 22 tipos diferentes de glomérulos olfativos.[2] La arquitectura de los glomérulos en los lóbulos antenales no refleja un dimorfismo sexual aparente.
En los triatominos existen tres diferentes tipos de pelos olfativos clasificados de acuerdo a su morfología: pelo basicónico, pequeño pelo surcado y pelo tricoideo.
Tipos de pelos olfativos
 Segmentos de la antena de un triatomino.
Segmentos de la antena de un triatomino.El pelo basicónico (single-(thin) walled wall-pore en inglés) de tipo 'E' esta conformado por una pared delgada cubierta por una alta densidad de poros conectados a pequeños tubos. No esta articulado en su base y mide[3] 30 µm en promedio. Su interior esta enervado por 21 a 41 células receptrices cuyas dendritas poseen ramificaciones. Estudios electrofisiológicos han determinado que las células nerviosas contenidas en este pelo responden incrementando la frecuencia de sus potenciales de acción en presencia del aliento humano y sustancias como piridina, furano, ácido pirúvico, ácido láctico y acetato de amila (óleo de banana). También se ha conseguido respuesta en este tipo de pelo a ciertos aldehídos como el nonanal.
El pequeño pelo surcado[4] (double-walled wall-pore en inglés) de tipo 'F' esta conformado por una pared doble cubierta por surcos longitudinales, de allí su nombre; y es relativamente pequeño midiendo de 6 a 10 µm de largo y 1 µm de diámetro. En los surcos longitudinales se encuentran poros que se comunican con la cavidad interior del pelo a través de canales que atraviesan la doble pared cuticular. Al igual que el pelo basicónico, este tampoco es articulado. Su interior esta enervado por 4 a 5 células receptrices que en contraste con el pelo basicónico, no se ramifican. Estudios de electrofisiología han demostrado que las células nerviosas contenidas en el pequeño pelo surcado responden al amonio y a algunas moléculas con este grupo funcional, es decir, aminas, así como a algunos ácidos grasos de cadena corta como el ácido isobutírico.
El pelo tricoideo (single-(thick) walled wall-pore en inglés) de tipo 'D' esta conformado por una pared gruesa. Aunque se desconoce su función, su aspecto hace pensar que pueda contener receptores gustativos.
tabla
Características morfológicas y funcionales de los tres tipos de sensila olfativas presentes en Triatoma infestans Tipo Característica de la pared Abundancia en cada antena Longitud (μm) Diámetro basal (μm) Número de células receptoras Respuesta electrofisiológica Referencias Ilustración pelo basicónico pared delgada 550 18-26 2 21-45 (~80 dendritas) aliento humano, piridina, furano, acetato de amila, heptanal, octanal, nonanal y respuestas bajas o moderadas al dióxido de carbono, monóxido de carbono, metanol y ciertos aldehídos Mayer 1968,[5] Bernard 1974,[6] Guerestein y Guerin 2001[7] pequeño pelo surcado pared doble con surcos longitudinales 350 6-10 1 4-5 humedad, ácido isobutírico, ácido butírico, ácido dimetilbutírico, ácido isovalérico, ácido 2-metilbutírico, amoníaco, isobutilamina y respuestas bajas o moderadas a metilamina, dimetilamina, etilamina, trimetilamina y dietilamina Bernard 1974, Taneja y Guerin 1997,[8] Guerestein y Guerin 2001, Diehl et al. 2003[9] 
pelo tricoideo pared gruesa con surcos longitudinales 35 35 1.5 1-2 no se han encontrado estímulos Bernard 1974 
Olores que los atraen
Cuando se ha experimentado con las emanaciones de un ratón, un hamster o algún otro vertebrado vivo, estas han resultado atraer efectivamente a algunos triatominos.[10] Incluso la orina de conejo es atractiva para Rhodnius prolixus. De los olores emanados por el ser humano, el aliento es el más atractivo. Wigglesworth y Gillett reportaron en 1934 como R. prolixus agitaba sus antenas en respuesta a la presencia de una piel de ratón,[11] aunque solo observaron un comportamiento de atracción cuando este estímulo era acompañado de calor.
Las sustancias químicas responsables de la orientación de los triatominos hacia el huésped no son bien conocidas. En su búsqueda se han elegido como candidatos para realizar experimentos de comportamiento las sustancias que estimulan los receptores olfativos de sus antenas, aunque no todas estas sustancias son capaces, por si solas, de inducir el comportamiento de atracción. También han servido como candidatos aquellas sustancias que atraen a otros insectos hamatófogos.
Dióxido de carbono
Josué A Nuñes mostró en 1982 que el dióxido de carbono por si solo basta para atraer a R. prolixus.[12] Efectivamente, el dióxido de carbono producido por el huésped a una concentración mayor que la del medio ambiente, es un estímulo que atrae, no solo a los triatominos, también a mosquitos, garrapatas, flebótomos y potencialmente a cualquier artrópodo que se alimente de sangre de vertebrado. La concentración de CO2 en el medio ambiente ronda los 400 ppm (0,04%) y el ser humano lo exhala a una concentración de 45000 ppm (4,5%), es decir, unas 100 veces por encima de la concentración atmosférica. Sin embargo, los triatominos responden a concentraciones hasta por debajo de los 1000 ppm (0,1%).
Algunas sustancias volátiles incrementan el efecto atractivo del CO2, tal como se ha observado con el ácido láctico en T. infestans, y con el amoníaco (NH3) en R. prolixus.
Amoníaco y algunas aminas
El NH3 por si solo atrae a T. infestans y R. prolixus. Esta molécula esta presente en el aliento de los vertebrados y sus excreciones urinarias y fecales. También se ha detectado NH3 en las deyecciones de los triatominos, sobre las cuales estos se agregan. Algunas aminas como la dimetilamina y la etilamina, excretadas por los vertebrados a través de la orina, atraen a T. infestans, R. prolixus y P. geniculatus.[13] Otro compuesto que se ha descubierto juega un papel importante en la vida de los triatominos es el ácido isobutírico que atrae a los triatominos y que es producido tanto por el huésped como por el insecto, en este último a través de las las glándulas de Brindley.
Ácidos carboxílicos
La mezcla, a una concentración especifica, de los ácidos propanoico, butírico, valérico y láctico ha resultado atractiva para T. infestans.[14] El ácido láctico está presente en la piel humana y los otros tres ácidos carboxílicos de cadena corta son frecuentes en los desechos de vertebrados tales como heces fecales.
Sustancias activadoras
Existen sustancias químicas que estimulan las células nerviosas de las antenas y que si bien no son capaces de atraer a los triatominos por si solas, inducen ciertos comportamientos de activación. Por ejemplo, el nonanal, la hexilamina y la isobutilamina incrementan la velocidad de desplazamiento de estos insectos.
Feromona sexual
El cortejo y copulación observado en varias especies de triatominos, ha mostrado ser más sencillo que en otras especies de insectos. La búsqueda del componente químicosensorial de este comportamiento, es decir, una feromona sexual putativa, ha llevado casi medio siglo. Inicialmente se creía que las glándulas de Brindley podrían estar implicadas. Sin embargo, estudios recientes sobre las glándulas metasternales señalan su implicación en el comportamiento sexual de los triatominos.[15] [16] [17] Los olores producidos por las glándulas metasternales estimulan el comportamiento de cortejo sexual. Además, el vuelo de los machos es activado por el olor de las hembras, posiblemente producido por estas glándulas.[18] Se han identificado 3-pentanona, 2-metilbutanol y 3-pentanol, como compuestos volátiles provenientes de las glándulas metasternales.
Fotofóbicos pero con los ojos bien abiertos
A pesar de sus hábitos nocturnos, los triatóminos hacen uso de la vista para orientarse. Sin embargo, no esta claro que información utilizan para buscar al huésped.
Ojos compuestos
Los triatóminos tienen ojos compuestos con aproximadamente 300 omatidios cada uno. Son ojos de aposición con bastones o rabdomas abiertos (una característica común en Heteróptera), en los cuales un anillo de seis rabdómeros de las células retinulares 1-6, rodean a un par central de rabdómeros de las células retinulares 6 y 8. Si bien, los ojos de superposición generalmente son el sistema más adecuado para insectos de vida nocturna, los ojos de los triatóminos cuentan con un sistema de adaptación a las condiciones de iluminación del día que les permite maximizar la cantidad de luz que llega a los receptores retinales en condiciones de poca luz.[19]
Ocelos
Los triatóminos poseen también un par de ocelos (ojos simples)[20] sobre el dorso de la cabeza, detrás de los ojos compuestos. Estos muestran un completo desarrollo en el estado adulto. Los ocelos, junto con los ojos compuestos, regulan la respuesta fototáctica, es decir, los triatóminos reconocen la iluminación del ambiente y responden a esta, generalmente evadiendo la luz intensa.[21]
Comportamiento
Los triatóminos muestran comportamientos antagónicos con respecto a la luz, de acuerdo su contexto fisiológico y ambiental. Se han observado varias especies que durante la noche son atraídas por la luz de las casas en zonas rurales o a trampas de luz que capturan insectos en vuelo. Sin embargo, lo más común es que estos insectos huyan de la luz, pues son de hábitos nocturnos.
En condiciones de laboratorio, al disponer una iluminación de baja intensidad (<0.06 microW/cm2), los triatóminos muestran fotofobia a la luz natural y a luces monocromáticas con longitudes de onda de 397, 458, 499 y 555 nm. Son menos sensibles a las longitudes de onda de 357 nm (ultravioleta) y 621 nm (naranja oscuro), así como a longitudes de onda entre 665 y 695 nm (la parte roja del espectro).[22]
Técnicas de investigación
Tradicionalmente se utilizan dos enfoques para estudiar la fisiología sensorial y el comportamiento de los triatominos: electrofisiológico y de comportamiento.
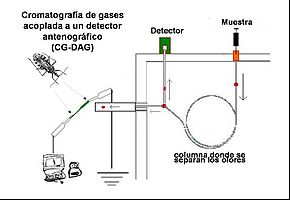 Los olores separados por la CG van simultáneamente al detector de este y a la antena viva conectada a un voltímetro.
Los olores separados por la CG van simultáneamente al detector de este y a la antena viva conectada a un voltímetro.Electrofisiología
El primero implica la búsqueda de respuestas electrofisiológicas de parte de las células nerviosas de olfación presentes en las antenas de estos insectos. Esto típicamente se puede hacer de dos maneras. Una, midiendo el cambio del potencial eléctrico de la antena cuando esta es estimulada con sustancias volátiles. A esta técnica se le llama electroantenografía (EAG). Si un incremento en la concentración de la sustancia que se está probando induce un incremento en la magnitud del cambio de potencial eléctrico, esto se debe a que efectivamente existen receptores olfativos que responden al olor en cuestión. La electroantenografía puede estar acoplada a un cromatógrafo de gases, de modo que los olores sean registrados simultáneamente por la antena y un detector electrónico. A esta técnica se le llama CG-DEA o (GC-EAD por sus siglas en inglés).
Otra manera más precisa de determinar la presencia de receptores olfativos para determinadas sustancias es a través del registro unicelular, conocido como SCR por sus siglas en inglés (single cell recording). Este método implica colocar, con la ayuda de un microscopio y un micromanipulador, un electrodo puntiagudo en las cercanías de las células nerviosas contenidas en los pelos de las antenas del insecto. El registro que se obtiene de tan minuciosa manipulación corresponde a los potenciales de acción que se producen en la célula olfativa. En presencia de un estímulo apropiado la frecuencia de los potenciales de acción aumenta o disminuye, e incluso puede cesar por completo. Un cambio en la frecuencia de los potenciales de acción es evidencia de una respuesta electrofisiológica.
Comportamiento
El enfoque de comportamiento implica determinar como se orientan estos insectos en presencia de determinados olores. Para hacer este tipo de observaciones se han utilizado distintos aparatos tales como el compensador de movimiento,[23] diferentes tipos de olfatómetros[24] y observaciones de campo en regiones endémicas. El compensador de movimiento (o servosfera) consiste en una esfera sobre la cual puede caminar un animal manteniéndose siempre sobre su ápex. La caminata, registrada por el moviemiento de la esfera, es almacenado en una computadora para su análisis.
Aplicaciones para el control de los triatominos
A fin de disminuir el riesgo de contagio de la enfermedad de Chagas en regiones endémicas, el desarrollo de métodos eficaces para controlar la presencia de triatominios en la vivienda humana se avizora como la aplicación más importante del estudio de la ofacción en triatominos. Las herramientas de monitoreo lucen prometoras dado que permiten a las autoridades sanitarias estimar los efectos de campañas de erradicación y medir la intensidad de los tratamientos a aplicar según la escala de infección. En la actualidad existen tres métodos de monitoreo de triatomions, a) a través de inspectores calificados, b) demoliendo casas y c) utilizando aparatos de monitoreo. Si bien, bajo ciertas circunstancias las dos primeras técnicas han demostrado mayor eficacia, son más costosas y laboriosas. Por ello recientemente se ha hecho más hincapié en el desarrollo de aparatos de moniotereo. Muchos de ellos han sido diseñados como especie de guaridas portátiles donde los triatomions consiguen refugio y si bien no son capturados, dejan señales de su presencia tales como, deposiciones fecales, restos de muda u otros despojos. También se han propuesto algunos aparatos que sirven de trampas con o sin señuelo atrayente, generalmente de tipo olfativo. En alguno casos se ha utilizado ratones vivos como atrayente, otras hielo seco o levadura como fuente de dióxido de carbono. También se ha propuesto el uso de sales que desprenden dióxido de carbono y amoníaco, compuestos que han demostrado atraer a los triatominos.
Bibliografía
- Brenner RR, Stoka AM (1987) Chagas’ disease vectors. I, II and III. CRC Press. Boca Ratón
- Cruz-López L, Malo EA, Rojas JC, Morgan ED (2001) Chemical ecology of triatomine bugs: vectors of Chagas disease. Med Vet Entom 15:351-357
- Otálora-Luna F (2006) Chemosensory and behavioural adaptations for haematophagy in triatomine bugs (Heteroptera: Reduviidae). Université de Neuchâtel Disponible en la Web (Video suplementario en Curiosidades)
- Rose AM (1998) Investigation into the host finding of a blood sucking bug from South America: Triatoma infestans (Klug) (Hemiptera: Reduviidae), a vector of Chagas disease. PhD Thesis, University of Regensburg
- Rozendaal JA (1997) Vector control: Methods for use by individuals and communities. WHO Disponible en la Web
Notas
- ↑ Gracco M, Catalá S (2000) Inter-specific and developmental differences on the array of antennal chemoreceptors in four species of Triatominae (Hemiptera: reduviidae). Disponible en la Web
- ↑ Romina B. Barrozo, Louise Couton, Teresita Insausti, Nadine Fresquet, Sebastian Minoli, Jean-Pierre Rospars, Sylvia Anton, Claudio Lazzari (2007) Antennal pathways in the central nervous system of Rhodnius prolixus. 10TH EUROPEAN SYMPOSIUM FOR INSECT TASTE AND OLFACTION (10th ESITO) Roscoff, Francia
- ↑ Estas medidas se refieren a Triatoma infestans adultos
- ↑ Algunos autores llaman a este pelo "basicónica".
- ↑ Mayer MS (1968) Response of single olfactory cell of Triatoma infestans to human breath. Nature 220: 924–925
- ↑ Bernard J (1974) Etude électrophysiologique de récepteurs impliqués dans l’orientation vers l’hôte et dans l’acte hématophage chez un hémiptère: Triatoma infestans. PhD Thesis. University of Rennes, France.
- ↑ Guerenstein PG, Guerin PM (2001) Olfactory and behavioural responses of the blood-sucking bug Triatoma infestans to odours of vertebrate hosts. J Exp Biol 204:585-97 Disponible en la Web
- ↑ Taneja J, Guerin PM (1997) Ammonia attracts the haematophagous bug Triatoma infestans: behavioural and neurophysiological data on nymphs. J Comp Physiol A 181:1–34
- ↑ Diehl PA, Vlimant M, Guerenstein P, Guerin PM (2003) Ultrastructure and receptor cell responses of the antennal grooved peg sensilla of Triatoma infestans (Hemiptera: Reduviidae). Arthropod Struct Dev 31:271–285
- ↑ La mayor parte de los experimentos de comportamiento olfativo se han realizado con R. prolixus y T. infestans.
- ↑ Wigglesworth VB, Gillett JD (1934) The function of the antenna in R. prolixus (Hemiptera) and the mechanism of orientation to the host. J Exp Biol 11:120-138
- ↑ Núñez, J.A. (1982) Food source orientation and activity in Rhodnius prolixus Stal (Hemiptera:Reduviidae). Bull. Entomol. Res., 72, 253–262.
- ↑ Otálora-Luna F (2007) Amines from excretions of nesting vertebrates guide triatomines to resources. Universidad de Neuchâtel Disponible en la Web
- ↑ Barrozo RB, Lazzari CR (2004) Orientation behaviour of the blood-sucking bug Triatoma infestans to short-chain fatty acids: synergistic effect of L-lactic acid and carbon dioxide. Chem Senses 9:833-41
- ↑ Manrique G, Vitta AC, Ferreira RA, Zani CL, Unelius CR, Lazzari CR, Diotaiuti L, Lorenzo MG (2006) Chemical communication in chagas disease vectors. Source, identity, and potential function of volatiles released by the metasternal and Brindley's glands of Triatoma infestans adults. J Chem Ecol 32:2035-52 Disponible en la Web
- ↑ Crespo JG, Manrique G (2007) Mating behavior of the hematophagous bug Triatoma infestans: role of Brindley's and metasternal glands. J Insect Physiol 53:708-14 [1]
- ↑ Pontes GB, Bohman B, Unelius CR, Lorenzo MG (2008) Metasternal gland volatiles and sexual communication in the triatomine bug, Rhodnius prolixus J Chem Ecol 34:450-7
- ↑ Manrique G, Renna de Vitta AC, Barcelos Pontes G, Zacharias CA, Lorenzo MG (2007) Chemical Communication in Triatomine Bugs’ Sexual Context. ESITO [Disponible en la Web http://www.esito-symp.org/2007_TalksAbstracts.htm]
- ↑ Reisenman CE, Insausti TC, Lazzari CR (2002) Light-induced and circadian changes in the compound eye of the haematophagous bug Triatoma infestans (Hemiptera: Reduviidae) J Exp Bio 205:201–210 Disponible en la Web
- ↑ Insausti TC, Lazzari CR (2002) The fine structure of the ocelli of Triatoma infestans (Hemiptera: Reduviidae). Tissue Cell 34:437-49
- ↑ Lazzari CR, Reiseman CE, Insausti TC (1998) The role of the ocelli in the phototactic behaviour of the haematophagous bug Triatoma infestans. J Insect Physiol 44:1159-1162
- ↑ Reisenman CE, Lazzari CR (2006) Spectral sensitivity of the photonegative reaction of the blood-sucking bug Triatoma infestans (Heteroptera: Reduviidae). J Comp Physiol A 192:39-44
- ↑ Otálora-Luna F, Perret JL, Guerin PM (2004) Appetence behaviours of the triatomine bug Rhodnius prolixus on a servosphere in response to the host metabolites carbon dioxide and ammonia. J Comp Physiol A 190:847-54 Disponible en la Web
- ↑ Aldana E, Otalora F, Abramson CI(2005) A new apparatus to study behavior of triatomines under laboratory conditions. Psychol Rep. 96:825-32 Disponible en la Web
Videos
Categorías:- Reduviidae
- Fisiología animal



Wikimedia foundation. 2010.