- Silenciamiento génico
-
El silenciamiento génico es un proceso llevado a cabo en los organismos eucariotes, con diferentes objetivos, dentro de los cuales se destacan la regulación de la expresión y la eliminación y el control de material genético ajeno o externo (virus y transposones) que podría causar un daño a la célula. Existe una clasificación que establece dos grandes categorías, el Silenciamiento Génico Post-Transcripcional (PTGS) y el Silenciamiento Génico Transcripcional (TGS), los cuales se encuentran en todos los eucariotes, con ciertas variaciones. El PTGS es un mecanismo que actúa directamente sobre el transcrito de ARN mensajero, mientras que el TGS actúa indirectamente y va a establecer un control que impide la transcripción del ADN.[1]
Se han reconocido diferentes moléculas y sistemas de estas que se encargan de silenciar la expresión y se encuentran especialmente en el PTGS. Dentro de estas moléculas se encuentran proteínas “Dicers”, argonautas, con actividad RNAsa, miRNA (micro ARN de interferencia), siRNA (ARN pequeño de interferencia) con variaciones como nat-siRNA (inducido durante situaciones de estrés) y ra-siRNA para la transcripción de secuencias repetidas. Adicionalmente se encuentran complejos que se acoplan con estos elementos como los RISC (Complejo de Silenciamiento inducido por ARN) que actúa a nivel citoplasmático acoplando siRNA producido, y el sistema RIST (Complejo de Silenciamiento Transcripcional Inducido por ARN) cuya actividad se localiza en el núcleo celular.[1]
Además de estas formas, se han encontrado otros niveles transcripcionales de regulación que no actúan sobre el ARNm, sino que modifican el ADN a través de procesos de metilación que van a modificar la cromatina, estableciendo dominios de silenciamiento a los cuales se enlazan proteínas SIR que evitan la transcripción. En las histonas del ADN también se han detectado modificaciones como poliadenilación, y deacetilación, cuyo efecto será impedir la unión del ADN con proteínas enlazantes.[2]
El silenciamiento, no es un proceso localizado, pues a través de uniones celulares como los plasmodesmos vegetales, hay un flujo de los elementos ya mencionados, de modo que se ejerce un efecto sistematizado. Una vez iniciado este proceso, se mantiene el silenciamiento, y un mecanismo para ello es sintetizar secundariamente siRNA a través de ARN polimerasas ARN dependientes (ARNdARNp). De esta forma los cambios muchas veces se prolongan por un tiempo, y en algunos casos, como la modificación de la heterocromatina, son heredados.[1]
Contenido
Moléculas Involucradas
Desde su descubrimiento en 1995 por Guo y Kemphues, con el silenciamiento del gen par-1 de C.elegans se ha mantenido un estudio constante respecto a los mecanismos naturales que se encuentran en una gran cantidad de organismos y que están involucrados en la regulación de la expresión.[3]
Heterocromatina
Una gran cantidad de ADN que actúa en centrómeros y telómeros estimulando el silenciamiento de genes aledaños en la eucromatina. No es codificante, y cerca a sus histonas se dan procesos de metilación que mantienen el silenciamiento. Esa heterocromatina va a producir siRNA que va a ser el encargado del silenciamiento interactuando con otras proteínas también asociadas a ella y a histonas. Se da igualmente un acople entre la heterocromatina con RITS y con ARNpARNd, para dar una acumulación de siRNA. Su efecto silenciador, está potencializado en parte por la acción de ARN que se asocia para estimular cambios. Las variaciones existentes entre los diferentes organismos son principalmente en la cantidad y tipo de repeticiones.[4]
Proteínas DICER y Argonautas
Las proteínas DICER, son nucleasas que degradan ARN de doble cadena, formando pequeños fragmentos. En sus extremos tienen sitios de unión de ARN y de proteínas argonautas. Las proteínas argonautas (Ago) se acoplan con el sistema RISC y su función será cortar ARNm blanco y a su vez guiar al complejo hacia su objetivo con mayor eficiencia.[1] [5]
siRNA (Fragmentos pequeños de ARN interferente)
Estos pequeños fragmentos de ARN son sintetizados a partir de ARN de doble cadena. En esa conformación las proteínas DICER van a reconocerla y ejercerán su efecto endonucleasa. Luego los pequeños fragmentos se acoplan con el sistema RISC, que va a romper la doble hebra para activar la actividad de siRNA. Se movilizan hacia ARNm y se unirán a él, impulsando de nuevo la actividad RNAsa. Estos fragmentos pueden ser templados para que una ARNdARNp sintetice más ARN de doble cadena, mantiendo la presencia de siRNA. Tiene varios tipos y algunos son inducidos bajo condiciones de estrés como nt-siRNA y otros actúan sobre las repeticiones nucleotídicas como el ra-siRNA.[1] [4]
miRNA (micro fragmentos de ARN interferente)
Es ARN no traducido que va a silenciar de la misma forma que los siRNA, acoplándose a complejos efectores como RISC. La principal diferencia entre los dos tipos es el tamaño, siendo estos los más pequeños.[1]
Complejo RISC
Este complejo es una agrupación de varias proteínas, incluyendo su unidad más importante que es una endonucleasa. Actúa a nivel citoplasmático y se encarga de enlazar el siRNA de doble cadena activándolo, llevándolo a cortar esos pequeños fragmentos en hebras sensibles, para dirigirlos posteriormente hacia los blancos a silenciar, estimulando su clivaje.[5]
Complejo RIST
Funciona de una forma semejante a la del complejo RISC, su única diferencia es que actúa a nivel nuclear.[1]
Silenciamiento Post-Transcripcional y Transcripcional
Silenciamiento Post-Transcripcional (PTGS)
El PTGS se caracteriza por estar mediado principalmente por siRNAs y miRNAs. En este tipo de silenciamiento además de encontrar esos dos elementos ya mencionados, se asocian los complejos RISC, mRNP, proteínas DICER, Argonautas y Drosha. El mecanismo general inicia con la síntesis de los pre-siRNAs y pre-miRNAs en el núcleo y su maduración. Este último proceso se da mediante proteínas DICER, cuyo centro catalítico dimérico, actúa de una forma similar a una RNAsa tipo III, clivando en pequeños fragmentos de doble cadena. De acuerdo al organismo del que estemos hablando actúan otras proteínas, pues en los vertebrados solo se produce un tipo de DICER, mientras que en animales como Drosophila hay más genes codificantes que dan DICERs más especializadas.[6]
Estos fragmentos son los que van a unirse al ARNm blanco, pero para ello se da un acople con el complejo RISC. Este complejo en su estructura posee proteínas argonautas Ago, que tienen dominios de unión (PAZ y Piwi) al ARN, y su actividad RNAsa H contribuye a que los siRNA de doble cadena sean procesados a cadena sencilla, volviéndolos funcionales. Existe una gran variedad de complejos RISC, de acuerdo al tamaño de iRNA que vaya a enlazar.[6]
Silenciamiento Transcripcional (TGS)
Aunque es más común encontrar procesos de PTGS en las células, los TGS son mecanismos de silenciamiento importantes, pues actúan antes de la transcripción. La forma más estudiada de TGS, ha sido la metilación del ADN, estimulada por una serie de enzimas como las proteínas Dicer, DNA metiltransferasas (DMTasas) y por la presencia de ciertos promotores que formarán conformaciones de ARN de doble cadena. A partir de esos promotores, las Dicers formarán fragmentos cortos de ARN de aproximadamente 21 nucleótidos, que serán útiles para unirse y ser reconocidos por DMTasas que se encargarán de metilar.[7]
El proceso de metilación se da a partir de proteínas específicas para los diferentes organismos, y se mantiene a partir de otro tipo de DMTasas que son conservadoras de la metilación, actuando en la replicación manteniendo la metilación en la hebra templado, y metilando la hebra sintetizada. Las proteínas que se han reconocido son varias, dentro de las cuales encontramos en los mamíferos Dnmt3a y Dnmt3b y en plantas sus ortólogos DRM1 y DRM2. Para la metilación es importante algunas veces la modificación estructural de la heterocromatina o de las histonas asociadas. El resultado de esto será modificar la expresión alterando promotores, represores, etc.[7]
Silenciamiento Génico en Animales
En los animales se ha reportado la presencia tanto de TGS como de PTGS, sin embargo en general a los mecanismos de silenciamiento en estos organismos se les conoce como ARNi o cosupresión. Su utilidad se basa en la protección de la integridad del genoma, evitando de esta forma los transposones, pero también estos mecanismos de silenciamiento establecen rutas de regulación durante el desarrollo que son indispensables para evitar anomalías. Particularmente para cada animal se han distinguido una variedad de rutas, sin embargo en términos generales los mecanismos encontrados son parecidos a los que ya se han mencionado.
Caenorhabditis elegans fue el animal que impulsó el estudio de estas rutas, y en este nemátodo se han detectado alrededor de 112 miRNA, de los cuales se conoce muy poco sobre su función. Algunos importantes han sido lin-4 y let-7, involucrados en el desarrollo del organismo, también lsy-6 y mir-273 que están involucrados en la formación de patrones del sistema nervioso. El mecanismo más común es el PTGS, usando siRNA producido por proteínas Dicer y utilizando los complejos RISC. Adicionalmente se ha encontrado una variedad de tipos de siRNA como por ejemplo el transitorio, producido desde el ARNm blanco mediante una ARNpARNd y algunos nucleares.[8]
Además de las funciones en el desarrollo, en el caso de los animales estas rutas de silenciamiento están involucradas en la protección de infecciones virales, aunque esto se ha estudiado más en plantas.
Silenciamiento Génico en Plantas
En las plantas es más común referirse a PTGS, TGS y a VIGS, este último hace referencia al silenciamiento inducido por virus. En las plantas se dan diferentes mecanismos, pero lo más interesante es que hay una conexión entre PTGS y TGS, ya que ambos pueden inducirse a partir de una misma molécula de ARN de doble cadena. El funcionamiento de PTGS es parecido al ya mencionado, en donde se acopla con un complejo efector para silenciar.
El proceso de TGS tiene varios mecanismos, y se han detectado proteínas como DDM1, MOM y MET1 que están involucradas en procesos de metilación, y en mutantes transgénicos de maíz se ha visto que su ausencia puede llevar a la activación de transposones y de secuencias sin función conocida, que alterarán el desarrollo de la planta, modificando su tamaño, volviéndolas estériles o afectando la floración. Por otra parte, su presencia en otros casos contribuye a evitar el silenciamiento de ciertos genes relacionados también con el desarrollo, como sup o ag.[9]
Existen dos tipos de acción en el mecanismo TGS, uno funciona en posición cis, y consiste en silenciar las copias múltiples en un mismo locus de un gen, formando estructuras en el ADN que estimulan la metilación. El otro tipo es el trans, donde se forma un intermediario de ARN corto de cadena sencilla a partir de uno de cadena doble (que puede inducir también PTGS), que se unirá al ADN blanco.[9]
La principal utilidad de estos mecanismos consiste en combatir las infecciones virales, formando fragmentos de ARN que son complementarios al ADN o al ARN viral, buscando su silenciamiento y de esta forma deteniendo el proceso de infección. No siempre es un mecanismo efectivo, pero se ha reportado para enfermedades como el virus del mosaico del coliflor y el virus X de la papa.[9]
Silenciamiento Génico y Desarrollo
En el desarrollo de todos los organismos el control de la expresión de genes es muy importante, pues solamente mediante un control estricto y sincronizado se desarrollan los patrones adecuados. Existe una gran cantidad de ejemplos en los que se ha reportado la importancia del silenciamiento en la formación de muchas estructuras corporales. A continuación se mencionan algunos casos.
Destino celular y Diferenciación celular
Una de las preguntas que antes se hacían con gran frecuencia era cómo se lograba una diferenciación celular en el desarrollo de los organismos, que permitiera a partir de unas cuantas células generar una gran cantidad de tejidos y órganos. Actualmente, la respuesta posiblemente más acertada a esta pregunta, está basada en el silenciamiento génico direccional, más que en una determinación definitiva de las células.
Dentro de los factores más estudiados está la distribución espacial de los genes, que se relaciona con mecanismos TGS, en donde de acuerdo a la posición de un gen respecto a dominios de heterocromatina, se dará un silenciamiento o expresión. Esta forma de represión, consiste en que entre más cercano se encuentre un gen a estos dominios, la probabilidad de ser silenciado es mayor, sin embargo, en Drosophila, se han descrito casos en los que por regulación trans, el gen brown en heterocigosis es afectado, y no se dará el fenotipo silvestre.[10]
Esas polarizaciones a lo largo de un eje se ha estudiado no solamente en la mosca de la fruta, también se ha visto en los fibroblastos de los mamíferos, en donde la diferenciación de la línea celular de los linfocitos T, está mediada por factores de heterocromatina que silencian otros genes como IL-4, permitiendo que se formen poblaciones de Th1 y Th2. También para las células B se ha visto algo parecido, hay genes silenciados que se ubican cerca a centrómeros de heterocromatina, permitiendo su diferenciación.[10]
De esta forma existen muchos mecanismos en los que se involucran varios genes, y el silenciamiento de algunos lleva a la diferenciación y formación de un tipo de células.
Embriogénesis
En la embriogénesis se dan varios cambios epigenéticos, que modifican el fenotipo manteniendo el genotipo, a través del silenciamiento génico. Existen varios mecanismos ya mencionados que se dan en las diferentes etapas de este proceso, con el objetivo de producir individuos multicelulares a partir de un cigoto.
La regulación de la expresión, no empieza sino hasta determinado punto del desarrollo del embrión, pues la transcripción no está activa todo el tiempo. En el caso de los ratones por ejemplo, esto comienza en el embrión con 2 células, mientras que en otros mamíferos como el humano o las ovejas, empieza en el estadio de 5-8 células. Los principales mecanismos son los TGS, dando una gran cantidad de eventos de metilación y demetilación.[11]
Se ha reportado el inicio de los eventos de metilación desde los núcleos parentales, especialmente en el del padre donde se da antes de la cariogamia una metilación, que inducirá cambios en el núcleo de la madre con el fin de estimular la compatibilidad. Adicionalmente se han caracterizado cambios durante la embriogénesis de la metilación para el desarrollo apropiado, como en Xenopus laevis en donde se bajan los niveles de metilación para entrar en el estadio de blástula media.[11]
Finalmente, otros cambios importantes que se dan en la embriogénesis están relacionados con las histonas, que inducirán ciertas estructuras de heterocromatina con el objetivo de silenciar o favorecer la expresión de ciertos genes, involucrados por ejemplo en la plasticidad de células pluripotentes. Estos cambios en las histonas también son inducidos durante la fecundación y durante el inicio de la gastrulación en animales como las ranas.[11]
Desarrollo temprano del ojo en Xenopus laevis
Se ha detectado la presencia de una familia de miRNAs que está asociada al desarrollo del sistema nervioso central en los vertebrados. En el caso de X.laevis, mediante técnicas de hibridación in situ se vio que miR-124 a se expresaba en diferentes estadios de su desarrollo, localizándose en diferentes partes pero especialmente en regiones como el mesencéfalo y el rombencéfalo. Si se llegara a sobre expresar, los blancos de ARNm cambiarían evitando la profileración celular, alterando la morfología. Lo mismo sucedería en su ausencia. En el ojo se detectó su expresión a nivel de la retina. Su blanco no es Pax6, sino un gen conocido como Lhx2 que es regulador de la división durante la retinogénesis.[12]
Silenciamiento génico y Enfermedades
La mayoría de las asociaciones que se han establecido entre el silenciamiento génico y las enfermedades están relacionadas con el ciclo celular. Muchos silenciadores, cumplen un rol fundamental en esto, de modo que mutaciones en estos genes como el silenciador p53 de EZH, llevan al desarrollo de cáncer. En pacientes que padecen de esta patología en la próstata, se ha visto un nivel elevado de EZH, pues no tienen el silenciador.[13]
Las metiltransferasas nuevamente cumplen un papel esencial como SUV39h1 que se encarga de metilar K9H3, para que se dé la unión de un dominio de heterocromatina. Su ausencia tiene diferentes efectos en el desarrollo pero también en otras funciones como las reproductivas. Se desarrollan linfomas, se da un crecimiento lento, etc.[13]
Por último las proteínas de la heterocromatina enlazantes como H1 son esenciales para evitar enfermedades, impulsando el silenciamiento de ciertos genes. Sin ellas se afecta la segregación de cromosomas, además son supresores de tumores, de modo que también se afectaría el ciclo celular. Adicionalmente se encargan de eliminar la transcripción de material genético viral como en VIH, o herpes virus, por lo tanto sin ellas, habría mayor vulnerabilidad.[13]
Aplicaciones
Las aplicaciones del silenciamiento génico son muy amplias, pues además de tener fines terapéuticos, su uso en investigación ha aumentado, pues mediante iRNA y las moléculas ya mencionadas se ha podido estudiar la función de varios genes.
Una de las aplicaciones con mayor potencial, es el uso de transplantes celulares para evitar el desarrollo de enfermedades como el cáncer. Modificando células madre de blastocistos humanos, empleando iRNA, es posible formar líneas celulares no defectuosas desde las células del paciente en medios de cultivo, para posteriormente ser implantadas.[14]
Actualmente se están investigando terapias a base de ARN que inhiban la trascripción, esta nueva tecnología se la ha denominado triplex. Funciona a través de oligo que no se une al un ARN mensajero (como haría el ARNi) sino que se une al ADN inhibiendo la trasncripción. Se une un oligo a la doble cadena formando una triple hélice. En teoría sería útil para mutaciones dominantes. Solo ha funcionado en promotores de microorganismos donde no hay superenrollamiento ni histonas.
Otra nueva vía de desarrollo es la construcción de ARN catalíticos (diferentes a las ribozimas). Se ha propuesto unir al oligo que emparejaría con el ARN mensajero del gen que queremos silenciar una molécula de EDTA-Fe. Esto provoca la destrucción activa del ARN mensajero al que se une de forma que el ARN catalítico puede liberarse para degradar otro ARN mensajero.
Otras de las aplicaciones terapéuticas radican en el control de infecciones virales, bacterianas y parasitarias. En el caso de las virales es mucho más sencillo, y se ha probado mediante el silenciamiento de genes involucrados en su replicación, o en receptores celulares importantes, esto se ha hecho con VIH, Hepatitis C y VSR (Virus Sincitial Respiratorio). Para el caso de las bacterias, no ha sido lo más utilizado, dado que no existen tantos blancos, sin embargo, se han planteado alternativas para silenciar la respuesta proinflamatoria inducida por muchos factores de virulencia bacterianos como toxinas. Finalmente, para las infecciones parasitarias, se han planteado alternativas por ejemplo para la malaria, proponiendo la creación de vectores artrópodos transgénicos que sean resistentes a la colonización y establecimiento del parásito.[15]
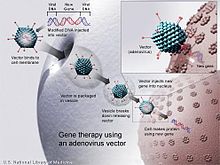 Uso de vectores virales para la terapia génica
Uso de vectores virales para la terapia génica
Existen dificultades metodológicas que se deben superar todavía para que estas alternativas sean funcionales y aplicadas. Los problemas principales están relacionados con la selección de secuencias adecuadas para diseñar el siRNA, la estabilidad de lo producido, es decir, la resistencia luego de ser inyectado, a las condiciones celulares (presencia de endonucleasas por ejemplo), la liberación (selección del vector adecuado) y lo más importante, la especificidad que garantice que no habrán efectos secundarios ni uniones a otros fragmentos que no son los blancos determinados. Se han ideado varios posibles vectores, algunos plasmídicos que mantengan la replicación de la secuencia, algunos virales con una eficiencia más grande, y por último la lipofección usando lípidos con el ADN en cultivos celulares. Para resolver estos problemas, falta mucho más estudio, pero adicionalmente debe reforzarse el trabajo con líneas celulares en medios, pues son modelos in vivo precisos, que podrían ser bastante útiles en el momento de diseñar una alternativa terapéutica basada en el silenciamiento de genes.[16]
Referencias
- ↑ a b c d e f g Martínez, W, & Peña, L. (2007). «El silenciamiento génico transcripcional y postranscripcional y la produccion de anticuerpos en plantas.». Revista Electrónica de Portales Médicos 2.
- ↑ Moazed, D. (2001). «Common themes in mechanisms of Gene Silencing.». Molecular Cell 8: pp. 489 – 498.
- ↑ Rahman, M., Ali, I., Husnain, T., Riazzudin, S (2008). «RNA interference: The study of gene silencing on plants and humans.». Biotechnology Advances 26: pp. 202 – 209.
- ↑ a b Zaratiegui, M., Irvin, D., Martienssen, R. (2007). «Non coding RNAs and Gene Silencing.». Cell 128: pp. 763 – 776.
- ↑ a b Aigner, A (2006). «silencing through RNA interference (in vivo): Strategies based on the direct application of siRNAs.». Journal of Biotechnology 124: pp. 12 – 25.
- ↑ a b Filipowicz, W., Jaskiewicz, L., Kolb, F., Pillai, R. (2005). «Post-transcriptional gene silencing by siRNAs and miRNAs.». Current Opinion in Structural Biology 15: pp. 331 – 341.
- ↑ a b Matzke, M., Aufsatz, W., Kanno, T., Daxinger, L., Papp, I., Mette, M., Matzke, A. (2004). «Genetic analysis of RNA-mediated transcriptional gene silencing.». Biochimica et Biophysica Acta 1677: pp. 129 – 141.
- ↑ Fisher, S. (2010). «Small- RNA mediated gene silencing pathways in C.elegans.». The International Journal of Biochemistry and Cell Biology, In press.
- ↑ a b c Vaucheret, H. & Fagard, M. (2001). «Transcriptional Gene Silencing in Plants: targets, inducers and regulators.». TRENDS in Genetics 17: pp. 29 – 36.
- ↑ a b Fisher, A. & Merkenshlager, M. (2002). «Gene silencing, cell fate and nuclear organization.». Current opinion in Genetics and Development 12: pp. 193 – 197.
- ↑ a b c Meehan, R., Dunican, D., Ruzov, A., Pennings, S. (2005). «Epigenetic silencing in embryogenesis.». Experimental Cell Research 309: pp. 241 – 249.
- ↑ Qiu, R., Liu, K., Mo, W., Flynt, A., Patton, J., Kar, A., Wu, J., He, R. (2009). «The role of miR-124 a in early development of the Xenopus eye.». Mechanisms of Development 126: pp. 804 - 816.
- ↑ a b c Moss, T. & Wallrath, L. (2007). «Connections between epigenetic gene silencing and human disease.». Mutation Research 618: pp. 163 – 174.
- ↑ Rassouli, F. & Matin, M. (2009). «Gene Silencing in human embryonic stem cells by RNA interference.». Biochemical and Biophysical Research Communications 390: pp. 1106 – 1110.
- ↑ Leung, R. & Whittaker, P. (2005). «RNA interference: from gene silencing to gene-specific therapeutics.». Pharmacology & Therapeutics 107: pp. 222 – 239.
- ↑ Takahashi, Y., Nishikawa, M., Takakura, N. (2009). «Nonviral vector-mediated RNA interference: Its gene silencing characteristics and important factors to achieve RNAi-based gene therapy.». Advance Drug Delivery Reviews 61: pp. 760 – 766.
Wikimedia foundation. 2010.