- Heterodontosauridae
-
 Heterodontosauridae
HeterodontosauridaeRango fósil: Triásico — Cretácico 
Clasificación científica Reino: Animalia Filo: Chordata Clase: Sauropsida Superorden: Dinosauria Orden: Ornithischia Suborden: Cerapoda (sin clasif.): Heterodontosauriformes Familia: Heterodontosauridae
Romer, 1966Géneros - Abrictosaurus
- Echinodon?
- Fruitadens
- Geranosaurus (nomen dubium)
- Heterodontosaurus (tipo)
- Lanasaurus
posible sinónimo del Lycorhinus[1] - Lycorhinus
- Manidens
- Tianyulong
Heterodontosauridae ("lagartos de dientes diferentes") es una familia de dinosaurios ornitisquios primitivos, que de acuerdo a nuevos estudios, estuvieron más relacionados con los marginocéfalos, en vez de ser ornitópodos basales como se consideraba anteriormente. A pesar de que sus fósiles son raros, se sabe que habitaron alrededor del planeta comenzando a finales del período Triásico y pudiendo sobrevivir hasta principios del Cretácico.
Los heterodontosáuridos fueron pequeños dinosaurios, con menos de 2 metros en longitud. Se conocen principalmente por sus característicos dientes, incluyendo colmillos alargados parecidos a caninos y muelas adaptadas para masticar, análogo de los de hadrosáuridos del Cretácico. Su alimentación era herbívora y posiblemente omnívora.
Contenido
Descripción
De entre los heterodontosáuridos, sólo se conoce un esqueleto completo del Heterodontosaurus. Además se han encontrado restos fragmentarios del esqueleto del Abrictosaurus, pero no se han descrito totalmente, mientras que del resto de heterodontosáuridos se conocen simplemente de fragmentos de mandíbula y dientes. Por tanto, la mayoría de las sinapomorfías de los heterodontosáuridos (rasgos de definición) se han descrito a partir de los dientes.[2] [1]
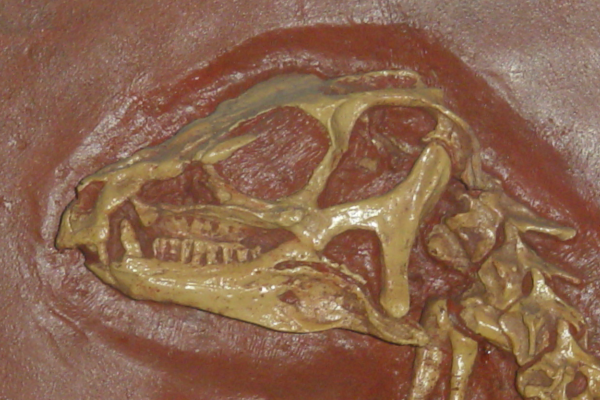
Cráneo
En ambos Abrictosaurus y Heterodontosaurus, los ojos eran muy largos, y debajo de los ojos, el hueso yugal (pómulo) se proyecta a los lados. Esta proyección, conocida como protuberancia, también se presenta en ceratopsianos. Como en las mandíbulas de muchos ornitisquios, el borde anterior del premaxilar no tenía dientes y probablemente en él se mantenía un pico queratinoso (rhamphoteca), aunque los heterodontosáuridos tuvieron dientes en la sección posterior del premaxilar. Un gran hueco, llamado diastema, separaba estos dientes premaxilares de los maxilares. La mandíbula inferior estuvo inclinada por el predental, un hueso único de los ornitisquios. Este hueso también apoyaba un pico similar al uno encontrado en el premaxilar. Todos los dientes mandibulares fueron hallados en el hueso dental.[2]
Dientes
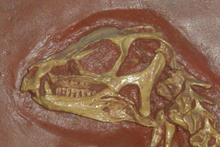
Los heterodontosáuridos son nombrados por su gran dentición heterodonte. Tuvieron tres dientes premaxilares. En el Abrictosaurus, Heterodontosaurus, y Lycorhinus del Jurásico inferior, los dos primeros dientes premaxilares fueron pequeños y cónicos, mientras los terceros mucho más largos se asemejan a dientes caninos de mamíferos carnívoros y son a menudo llamados caniniformes o colmillos. Un caniniforme inferior, más largo que los superiores, tomaba la primera posición en el dental y se acomodaba por el diastema de la mandíbula superior cuando la boca estaba cerrada.[2] Estos caniniformes estuvieron aserrados en ambos bordes anterior y posterior en el Heterodontosaurus y Lycorhinus, mientras los del Abrictosaurus presentaban dentaduras sólo en el borde anterior.[3] [4] En el Echinodon del Cretácico inferior, el caniniforme superior era aparentemente el primer diente del maxilar, y pudieron haber dos caniniformes inferiores en cada dentario,[5] y en Fruitadens del Jurásico Superior puede haber tenido dos dientes caniniformes inferiores en cada dentario.[6] [7]
Como los característicos colmillos, las muelas derivadas de los heterodontosáuridos son también únicos entre los ornitisquios primitivos. En los bordes de estos dientes hubo pequeñas elevaciones, o dentículos, los cuales alinearon el tercero de la corona del diente más cercana al tope en todos los heterodontosáuridos; en otros ornitisquios, los dentículos se extienden más abajo hacia la raíz. Formas basales como el Abrictosaurus tuvieron muelas en ambos maxilar y dental que fueron generalmente similares a los de otros ornitisquios: ampliamente espaciados, cada uno teniendo una baja corona y un cíngulo perfectamente desarrollado, una elevación la cual separaba la corona del diente de la raíz. En formas más derivadas como el Lycorhinus y Heterodontosaurus, los dientes tuvieron forma de cincel, con coronas mucho más altas y sin cíngulo, así que no hubo diferencia en el ancho entre las coronas y las raíces. Estas derivadas muelas también se podían recubrir una a otra, por lo que sus coronas formaban una superficie continua en la cual la comida pudo ser masticada. Las filas de dientes eran la inserción del lado de la boca, dejando un espacio fuera de los dientes el cual pudo haber sido cubierto por una mejilla muscular, la cual podría haber sido necesaria para masticar.[2] Los posteriores hadrosaurios y ceratopsianos del período Cretácico, tal como los mamíferos herbívoros, hubieran convergentemente evolucionado baterías dentales algo análogas. A diferencia de los hadrosaurios, los cuales podían tener miles de dientes constantemente siendo reemplazados, el reemplazo de dientes en heterodontosáuridos ocurría mucho más lentamente y varios especímenes han sido hallados sin ningún reemplazo de dientes en espera. Los heterodontosáuridos poseían una única artículación esferoide entre los dentarles y el predental, permitiendo a las mandíbulas inferiores rotar hacia fuera como la boca estaba cerrada, moliendo las muelas una contra otra. Debido al ritmo lento de reemplazo, este desgaste extremo producido del diente que comúnmente eliminó muchos de los dentículos en dientes viejos, aunque la altura aumentada de las coronas diera una vida larga a cada diente.[8]
Esqueleto
La anatomía postcranial del Heterodontosaurus tucki se la ha analizado bien, sin embargo H. tucki es generalmente considerado el más derivado de los heterodontosáuridos del Jurásico inferior, así que es imposible saber cuando características compartió con otras especies.[2] Los miembros delanteros fueron largos para un dinosaurio, más del 70% de la longitud de los miembros traseros. La cresta deltopectoral (una elevación para la atadura de los músculos del brazo y del pecho) bien desarrollada del húmero y el prominente proceso del olécranon del cúbito indican que el miembro delantero fue de gran alcance también. Tuvieron 5 dedos en las "manos". El primero, el pulgar, fue grande y provisto de una garra afiladamente curvada. Cuando lo doblaba, este primer dedo haya rotado hacia dentro; se lo ha llamado el "pulgar-retorcido".[9] El segundo dedo fue el más largo, levemente más largo que el tercero, y ambos también poseían garras. Los dedos cuarto y quinto no tenían garras, eran muy pequeños y simples en comparación. En el miembro trasero, la tibia fue 30% más larga que el fémur, el cual es generalmente considerado una adaptación para velocidad. La tibia y el peroné estuvieron fusionados con el astrágalo y el calcáneo del tobillo, formando un tibiofibiotarso convergentemente con aves modernas. También similarmente a las aves, los huesos bajos del tarso y los metatarsos estaban fusionados para formar un tarsometatarso. Hay cuatro dedos en la pata, con sólo el segundo, el tercero y el cuarto contactando el suelo. La cola, a diferencia de las de muchos otros ornitisquios, no tenía tendones osificados para mantener una postura rígida y fue probablemente flexible.[10] El esqueleto fragmentario que se conoce del Abrictosaurus nunca ha sido totalmente descrito, aunque el miembro delantero y las manos son mucho más pequeñas. El cuarto y quinto dedo del miembro delantero soportan un hueso falange menos que los del Heterodontosaurus.[11]
Protoplumas
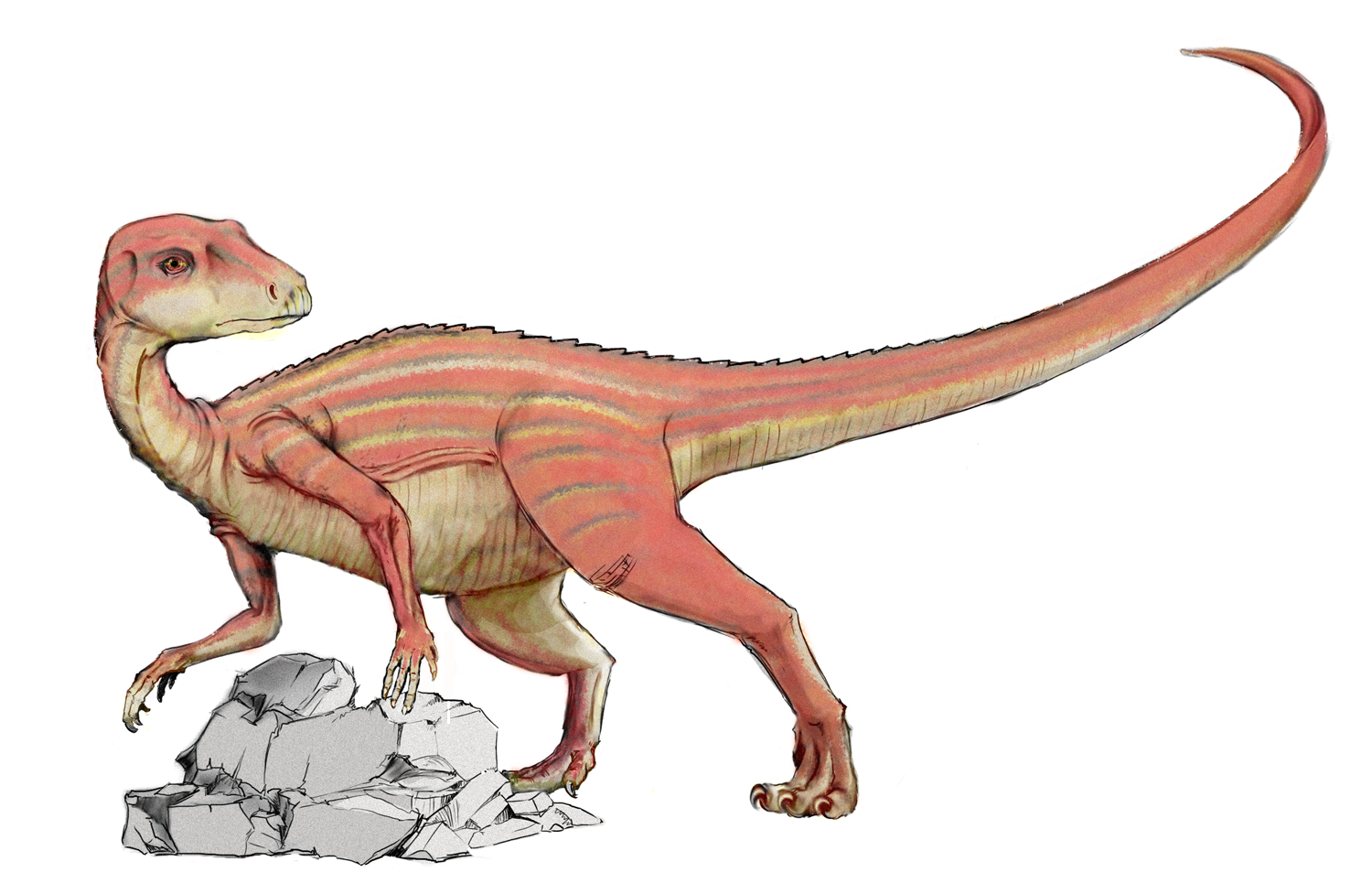
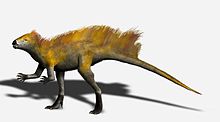 Reconstrucción de Tianyulong
Reconstrucción de Tianyulong
El Tianyulong de China parece preservar un tejido filamentosos el cual ha sido interpretado como una variante de las protoplumas halladas en algunos terópodos. Estos filamentos incluyen una especie de cresta a lo largo de su cola. La presencia de este tejido filamentoso ha sido usada para sugerir que tanto los ornitisquios como los saurisquios eran animales endotérmicos.[12]
Taxonomía y clasificación
Cladogramas de Heterodontosauridae
Butler et al., 2011[13] Heterodontosauridae NHM RU A100
Butler et al. 2010[7] Heterodontosauridae void void void NHM RU A100
En 1966, Alfred Romer nombró a esta familia Heterodontosauridae en referencia al género tipo Heterodontosaurus.[14] Tales géneros como el Echinodon y el Pisanosaurus han sido incluidos. Sin embargo, el Pisanosaurus ahora no es considerado un heterodontosáurido, sino un ornitisquio más basal.[15]
Antes visto como un clado hermano de Euornithopoda dentro de Ornithopoda, los heterodontosáuridos pudieron haber sido en cambio el grupo hermano de Marginocephalia, como lo indicó el reciente descubrimiento del Yinlong.[16]
El clado fue definido en 1998 por Paul Sereno y redefinido por él mismo en 2005 como un clado raíz consistiendo del Heterodontosaurus tucki y todas las especies más cercanamente relacionadas al Heterodontosaurus que el Parasaurolophus walkeri, Pachycephalosaurus wyomingensis, Triceratops horridus o Ankylosaurus magniventris.[17] [18]
Distribución
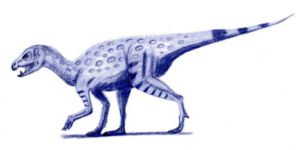 Heterodontosaurus.
Heterodontosaurus.Aunque originalmente eran sólo encontrados al sur de África de comienzos del Jurásico, ahora restos de heterodontosáurido se conocen en cuatro continentes. Los restos más primitivos que se conocen son un fragmento de mandíbula y dientes aislados de la Formación de Laguna Colorada de Argentina, los cuales datan del Triásico superior. Estos restos tienen uno morfología derivada similar a la del Heterodontosaurus, incluyendo un caniniforme con dentaduras en ambos bordes anterior y posterior, así como los dientes maxilares altamente coronados que carecen de un cíngulo.[19] La fauna heterodontosáurida más diversa se encuentra a comienzos del Jurásico en el sur de África, donde los fósiles del Heterodontosaurus, Abrictosaurus, y Lycorhinus han sido hallados. Otro taxón de estos mismos yacimientos, Geranosaurus, es a veces considerado un nomen dubium ya que todas las coronas de los dientes faltan en el espécimen tipo, siendo imposible de identificar si pertenece a una de las otras especies existentes.[2] Aunque, la aparente falta de un diastema puede indicar que es único.[20] El Lanasaurus, conocido solamente de restos de mandíbula,[21] se lo creyó ser particularmente basal,[2] pero simplemente puede ser el mismo que Lycorhinus.[1]
Heterodontosáuridos del Jurásico inferior, aún sin describir, también se conocen de los Estados Unidos y México, respectivamente.[22] [23] En adición, abundante material fósil ha sido descubierto en las capas de finales del Jurásico de la Formación de Morrison cerca de Fruita, Colorado, en los Estados Unidos. Descrito oficialmente en 2009, este mateial antes conocido como el "heterodontosaurio de Fruita" fue situado en el género Fruitadens.[7] [24] Los restos del Echinodon han sido recientemente redescritos y pueden representar un último heterodontosáurido sobreviviente de la época Berriasiana del Cretácico inferior al sur de Inglaterra.[5] Dientes de heterodontosáuridos que carecen del cíngulo dental ambién se han descrito de formaciones geológicas del Jurásico Tardío y el Cretácico Inferior en España y Portugal.[25] Dianchungosaurus del Jurásico Temprano de China ya no es más considerado un heterodontosáurido; en cambio se ha descubierto una forma relicta de principios del Cretácico Inferior de ese país, nombrada como Tianyulong.[12]
Paleobiología
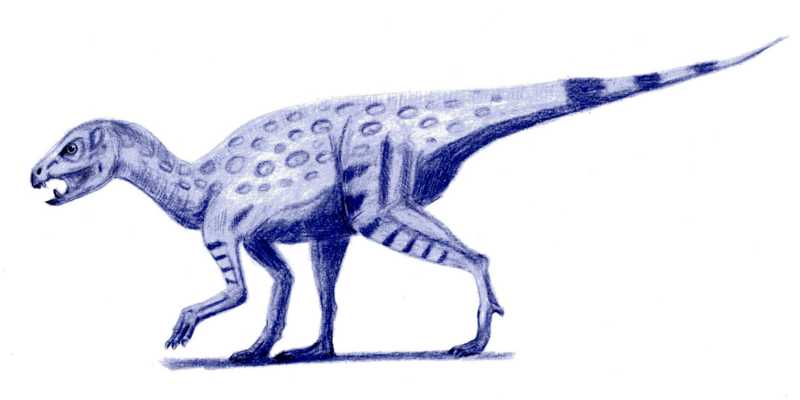
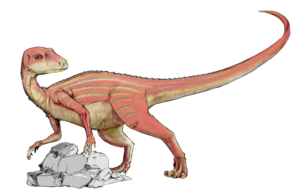 Abrictosaurus.
Abrictosaurus.La mayoría de fósiles de heterodontosáuridos se encuentran en formaciones geológicas que representa ambientes áridos y semiáridos, incluyendo la Formación Superior de Elliot del sur de África y las capas de Purbeck del sur de Inglaterra.[1] Se ha sugerido que los heterodontosáuridos experimentaban veraneo o hibernación estacional durante los tiempos más secos del año. Debido a la falta de reemplazo de dientes en muchos heterodontosáuridos, fue propuesto que el conjunto entero de dientes era reemplazado durante este período inactivo, mientras que parecía que el reemplazo continuo y esporádico de dientes interrumpiría la función de la fila de dientes como sola superficie de masticación.[11] Sin embargo, esto está basado en un malentendido de las mecánicas de mandíbula de heterodontosáurido, y también se ha demostrado que los heterodontosáuridos sustituían sus dientes continuamente, aunque más lentamente que en otros reptiles.[26] Recientemente no hay evidencia que aporte la hipótesis de veraneo en heterodontosáuridos.[2]
Mientras que las muelas de los heterodontosáuridos estuvieron claramente adaptados para triturar plantas, su dieta pudo haber sido omnívora. Los dientes premaxilares puntiagudos y las garras curvadas y afiladas en los miembros delanteros demuestran algún grado de comportamiento rapaz. Se ha sugerido que los miembros delanteros del Heterodontosaurus pudieron ser útiles para desgarrar en nidos de insectos, similar a los osos hormigueros modernos. Estos miembros delanteros también pudieron funcionar como unas herramientas de excavación, quizás por raíces y tubérculos.[2]
La longitud del miembro delantero comparado a la del miembro trasero sugiere que el Heterodontosaurus pudo haber sido parcialmente cuadrúpedo, y el proceso prominente del olécranon y los dedos hiperextendibles del miembro delantero se encuentran en muchos cuadrúpedos. Sin embargo, las manos están perfectamente diseñadas para agarrar, no como soporte de peso. Muchos rasgos del miembro trasero, incluyendo las largas tibia y pata, así como la fusión de los tibiofibiotarsos y tarsometatarsos, indican que los heterodontosáuridos estuvieron adaptados para correr rápidamente sobre sus miembros traseros, por lo tanto es improbable que el Heterodontosaurus se haya movido sobre sus cuatro miembros.[10]
Los cortos colmillos encontrados en todos los heterodontosáuridos conocidos impresionantemente recuerdan los colmillos encontrados en ciervos almizcleros, pecaríes y cerdos. En estos animales (y los de colmillos más largos como la morsa y los elefantes asiáticos), hay un peculiar dimorfismo sexual, con los colmillos solamente hallados en machos. El espécimen tipo del Abrictosaurus carece de colmillos y fue originalmente descrito como una hembra.[11] Aunque las vértebras sacras no fucionadas y la cara corta indican que este espécimen representa un animal juvenil, mientras un espécimen más grande de Abrictosaurus claramente posee colmillos. Por lo tanto, es posible que los colmillos se encuentren sólo wen adultos, en vez de ser una característica sexual secundaria de machos. Estos colmillos pudieron haberse usado en combates o despliegues con miembros de la misma especie o con otras especies.[2]
Referencias
- ↑ a b c d Norman, David B.; Sues, Hans-Dieter; Witmer, Lawrence M.; and Coria, Rodolfo A. (2004). «Basal Ornithopoda». En Weishampel, David B.; Osmólska, Halszka.; and Dodson, Peter. (eds.).. The Dinosauria (2nd edition edición). Berkeley: University of California Press. pp. 393-412. ISBN 0-520-24209-2.
- ↑ a b c d e f g h i j Weishampel, David B.; and Witmer, Lawrence M. (1990). «Heterodontosauridae». En Weishampel, David B.; Osmólska, Halszka.; and Dodson, Peter. (eds.).. The Dinosauria. Berkeley: University of California Press. pp. 486-497. ISBN 0-520-06727-4.
- ↑ Thulborn, Richard A. (1970). «The systematic position of the Triassic ornithischian dinosaur Lycorhinus angustidens». Zoological Journal of the Linnean Society 49: pp. 235-245.
- ↑ Hopson, James A. (1975). «On the generic separation of the ornithischian dinosaurs Lycorhinus and Heterodontosaurus from the Stormberg Series (Upper Triassic) of South Africa». South African Journal of Science 71: pp. 302-305.
- ↑ a b Norman, David B. (2002). «Ornithischian dinosaurs from the Lower Cretaceous (Berriasian) of England». En Milner, Andrew; and Batten, David J. (eds.). Life and Environments in Purbeck Times. Special Papers in Palaeontology 68. London: Palaeontological Association. ISBN 0901702730.
- ↑ Galton, Peter M. (2007). «Teeth of ornithischian dinosaurs (mostly Ornithopoda) from the Morrison Formation (Upper Jurassic) of the western United States.». En Kenneth Carpenter (ed.). Horns and Beaks: Ceratopsian and Ornithopod Dinosaurs. Bloomington: Indiana University Press. pp. 17–47. ISBN 0-253-34817-X.
- ↑ a b c Butler, Richard J.; Galton, Peter M.; Porro, Laura B.; Chiappe, Luis M.; Henderson, D. M.; and Erickson, Gregory M. (2010). «Lower limits of ornithischian dinosaur body size inferred from a new Upper Jurassic heterodontosaurid from North America» (PDF). Proceedings of the Royal Society B 277 (1680): pp. 375–381. doi:. PMID 19846460. PMC 2842649. http://rspb.royalsocietypublishing.org/content/early/2009/10/20/rspb.2009.1494.full.pdf.
- ↑ Weishampel, David B. (1984). Evolution in jaw mechanics in ornithopod dinosaurs. Advances in Anatomy, Embryology, and Cell Biology, 87. Berlin; New York: Springer-Verlag. ISBN 0387131140.
- ↑ Bakker, Robert T. (1986). The Dinosaur Heresies: New Theories Unlocking The Mystery of the Dinosaurs and Their Extinction. New York: William Morrow. pp. 453. ISBN 0140100555.
- ↑ a b Santa Luca, Albert P. (1980). «The postcranial skeleton of Heterodontosaurus tucki (Reptilia, Ornithischia) from the Stormberg of South Africa». Annals of the South African Museum 79 (7): pp. 159-211.
- ↑ a b c Thulborn, Richard A. (1974). «A new heterodontosaurid dinosaur (Reptilia: Ornithischia) from the Upper Triassic Red Beds of Lesotho». Zoological Journal of the Linnean Society 55: pp. 151-175.
- ↑ a b Zheng, Xiao-Ting; You, Hai-Lu; Xu, Xing; Dong, Zhi-Ming (19 de marzo de 2009). «An Early Cretaceous heterodontosaurid dinosaur with filamentous integumentary structures». Nature 458 (7236): pp. 333–336. doi:. PMID 19295609.
- ↑ Richard J. Butler, Jin Liyong, Chen Jun, Pascal Godefroit (2011). «The postcranial osteology and phylogenetic position of the small ornithischian dinosaur Changchunsaurus parvus from the Quantou Formation (Cretaceous: Aptian–Cenomanian) of Jilin Province, north-eastern China». Palaeontology 54 (3): pp. 667–683. doi:. http://onlinelibrary.wiley.com/doi/10.1111/j.1475-4983.2011.01046.x/abstract.
- ↑ Romer, Alfred Sherwood (1966). Vertebrate Paleontology, 3rd edition. Chicago: University of Chicago Press. pp. 468 pp.. ISBN 0-7167-1822-7.
- ↑ Weishampel, David B.; and Witmer, Lawrence M. (1990). «Lesothosaurus, Pisanosaurus, and Technosaurus». En Weishampel, David B.; Osmólska, Halszka.; and Dodson, Peter. (eds.).. The Dinosauria. Berkeley: University of California Press. pp. 416-425. ISBN 0-520-06727-4.
- ↑ Xu X.; Forster, C.A.; Clark, J.M.; and Mo J. (2006). «A basal ceratopsian with transitional features from the Late Jurassic of northwestern China». Proceedings of the Royal Society B: Biological Sciences 273: pp. 2135-2140. doi:.
- ↑ Sereno, Paul C. (1998). «A rationale for phylogenetic definitions, with application to the higher-level taxonomy of Dinosauria». Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 210 (1): pp. 41-83.
- ↑ Sereno, Paul C. (07-11-2005). «Stem Archosauria—TaxonSearch». Consultado el 24-02-2007.
- ↑ Baez, A.; and Marsicano, C.A. (2001). «A heterodontosaurid ornithischian dinosaur from the Upper Triassic of Patagonia.». Ameghiniana 38: pp. 271-279.
- ↑ Crompton, A.W.; and Charig, Alan (1962). «A new ornithischian from the Upper Triassic of South Africa». Nature 196: pp. 1074-1077.
- ↑ Gow, Christopher E. (1975). «A new heterodontosaurid from the Redbeds of South Africa showing clear evidence of tooth replacement». Zoological Journal of the Linnean Society 57: pp. 335-339.
- ↑ Sues, Hans-Dieter; Clark, J.M.; and Jenkins, F.A. (1994). «A review of the Early Jurassic tetrapods from the Glen Canyon Group of the American Southwest». En Fraser, N.C.; and Sues, H.-D. (Eds.). In The Shadow of the Dinosaurs: Early Mesozoic Tetrapods. Cambridge: Cambridge University Press. pp. 285-294. ISBN 0521458994.
- ↑ Clark, J.M.; Hernandez, R.R.; Montellano, M.; Hopson, James A.; and Fastovsky, David E. (1994). "An Early or Middle Jurassic tetrapod assemblage from the La Boca Formation, northeastern Mexico." In The Shadow of the Dinosaurs: Early Mesozoic Tetrapods, 295-302.
- ↑ Error en la cita: El elemento
<ref>no es válido; pues no hay una referencia con texto llamadaPMG06 - ↑ Sánchez-Hernández, Barbara; Benton, Michael J.; & Naish, Darren. (2007). «Dinosaurs and other fossil vertebrates from the Late Jurassic and Early Cretaceous of the Galve area, NE Spain.». Palaeogeography, Palaeoclimatology, Palaeoecology 249 (1-2): pp. 180–215. doi:.
- ↑ Hopson, James A. (1980). «Tooth function and replacement in early Mesozoic ornithischian dinosaurs: implications for aestivation». Lethaia 1: pp. 93-105.
Enlaces externos
 Portal:Dinosaurios. Contenido relacionado con Dinosaurios.
Portal:Dinosaurios. Contenido relacionado con Dinosaurios. Wikiespecies tiene un artículo sobre Heterodontosauridae. Wikispecies
Wikiespecies tiene un artículo sobre Heterodontosauridae. Wikispecies Wikimedia Commons alberga contenido multimedia sobre Heterodontosauridae. Commons
Wikimedia Commons alberga contenido multimedia sobre Heterodontosauridae. Commons- Datos científicos de Heterodontosauridae en The Paleobiology Database (en inglés)
- Detalles sobre Heterodontosauridae en Taxon Search (en inglés)
Categorías:- Fósiles
- Cerapoda


Wikimedia foundation. 2010.