- Parasaurolophus
-
Parasaurolophus
?Parasaurolophus
Rango fósil: Cretácico superior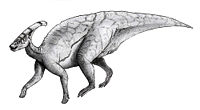
Estado de conservación Fósil Clasificación científica Reino: Animalia Filo: Chordata Clase: Sauropsida Superorden: Dinosauria Orden: Ornithischia Suborden: Neornithischia Infraorden: Ornithopoda Familia: Hadrosauridae Subfamilia: Lambeosaurinae Género: Parasaurolophus
Parks, 1922Especies Parasaurolophus (gr."cerca del lagarto crestado")[1] es un género de dinosaurios ornitópodos hadrosáuridos, que vivieron a finales del período Cretácico, hace aproximadamente 83 y 71 millones de años, en el Campaniano, en lo que hoy es Norteamérica. Su nombre hace referencia a un supuesto parentesco con el dinosaurio Saurolophus. Fue un herbívoro que caminaba tanto de manera bípeda como cuadrúpeda. Se conocen tres especies, P. walkeri, la especie tipo, P. tubicen y la de cresta corta P. cyrtocristatus. Sus restos se han encontrado en Canadá, en las formaciones Oldman y Dinosaur Park ambas parte del Grupo del Río Judith en Alberta (P. walkeri),[2] y en Estados Unidos, siendo encontrados en Nuevo México en la Formación Kirtland (P. tubicen),[3] en la Formación Fruitland (P. cyrtocristatus),[4] en Utah, en la Formación Kaiparowits (P. cyrtocristatus)[5] y por ultimo en Montana, en la Formación Hell Creek (P. walkeri).[2] Descrito por primera vez en 1922 por William Parks a partir de un cráneo hallado en Alberta.[6]
Parasaurolophus es un hadrosáurido, parte de la diversa familia de dinosaurios cretácicos conocidos por la variedad de extraños adornos en la cabeza. Este género en conocido por su gran y elaborada cresta craneal, con la forma de un tubo que se proyecta hacia atrás y arriba del cráneo. Charonosaurus de China, que puede ser su pariente más cercano, posee un cráneo muy parecido y una posible cresta similar. La cresta ha sido objeto de muchas discusiones entre los paleontólogos, siendo el consenso actual que las funciones importantes de esta incluyeron el reconocimiento visual de la especie y del sexo, resonancia acústica, y termorregulación. Es uno de los dinosaurios de pico de pato más raros, conocido solamente de un puñado de buenos especímenes
Contenido
Descripción
 Comparación de tamaños entre Parasaurolophus y Homo sapiens.
Comparación de tamaños entre Parasaurolophus y Homo sapiens.
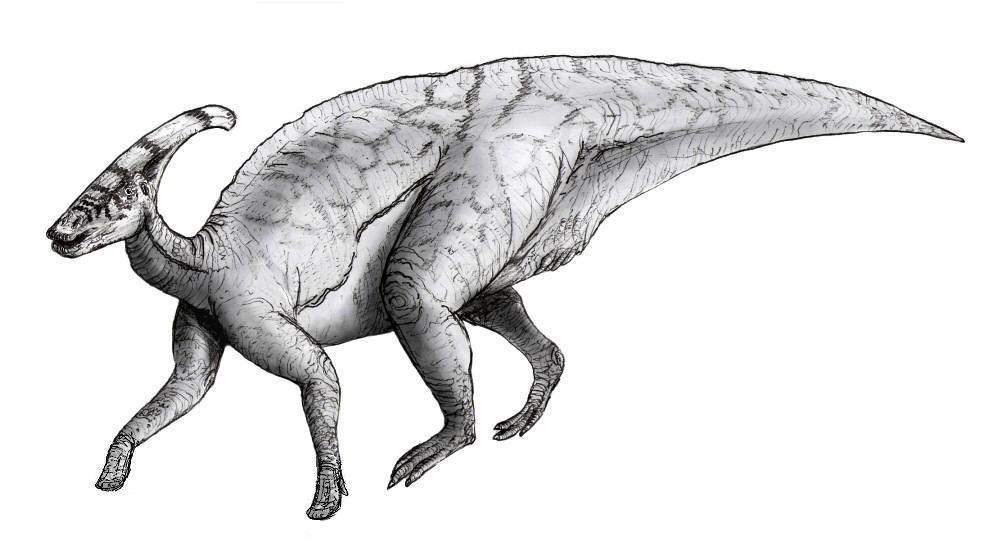
Como es el caso de la mayoría de los dinosaurios, el esqueleto de Parasaurolophus no es completamente conocido. El largo del cuerpo del espécimen tipo de P. walkeri se estima en 10 metros. El cráneo mide 1,6 metros de largo, incluyendo la cresta, sin embargo el cráneo del P. tubicen supera de largo, lo que indicaun amimal aun más grande.[7] El peso estimado es de 2,5 toneladas.[8] El unico miembro delanero conocido es relativamente corto para un hadrosáurido, con una corta pero ancha escapula. El fémur mide 103 centímetros en P. walkeri y es robusto para su longitud cuando es comparado al de los otros hadrosáuridos.[7] El humero y los huesos pelvicos estan fuertemente construidos.[9]
Como otros hadrosáuridos, es posible que caminara tanto en dos como en cuatro patas. Es probable que se parara en cuatro patas para forrajear, pero se desplazaría en dos.[10] El proceso espinoso de las vértebras era alto, algo común en los lambeosaurinos;[7] más alta sobre las caderas, aumentaron la altura de la parte posterior. Las impresiones de la piel que son conocidas en P. walkeri, muestran una estructura uniforme de escamas similares a tubérculos, pero sin ninguna particularidad en especial.[6]
 Parasaurolophus walkeri con detalles de la piel.
Parasaurolophus walkeri con detalles de la piel.Estos animales tenían la cola larga y aplastada a los lados lo que lleva a pensar que Parasaurolophus podía nadar. Al final de las extremidades es un misterio, algunos paleontólogos dicen que son pezuñas y otros dicen que son garras desgastadas por el tiempo. Se han encontrado impresiones de piel de Parasaurolophus, así como varios esqueletos completos por lo que se tiene una muy buena idea de su aspecto.
La cresta
Lo más remarcable de este dinosaurio es la cresta, la cuál se despega de la parte posterior de la cabeza y estaba compuesta del hueso premaxilar y el nasal. El especímen tipo de P. walkeri tiene una muesca en las espinas dorsales de las vértebras de cerca donde la cresta golpearía la espalda, pero esto puede ser a patología peculiar a este individuo.[7] William Parks, quien nombrara el género, hipotizo que un ligamento se encontraba desde la cresta a la muesca para ayudar a soportar la cabeza.[6] Aunque esta idea parezca inverosímil,[8] Parasaurolophus es a veces restaurado con una vela de piel desde la cresta al cuello. Esta enorme cresta tubular con cuatro secciones huecas dos que aputaban hacia arriba y dos hacia abajo de largo que se conectaba con la nariz, la cual se creía que podía servir como tubo para respirar debajo del agua, suposición que fue desmentida por estudios posteriores, los cuales establecieron la ausencia de algún tipo de orificio en la parte superior. Se han propuesto muchas hipótesis para explicar la función de la cresta, aunque la mayoría de ellas han sido desacreditadas.[11] [8] Se han dado varias hipotesis acerca de su cresta como respirar debajo del agua teoría que se creyó durante mucho tiempo hasta que fue desmentida. Se cree que era para la atracción y distincion entre machos y hembras, para comunicarse en caso de algun peligro u orden y para termorregulación.[12] La más sobresaliente es la de la comunicación. Esta se realizó observando la estructura interna del tubo, para concluir que pudo funcionar como un resonador natural, y haber tenido una función sonora.[13]
A raíz de modelos hechos de la cresta, se ha sugerido que se empleaba para para emitir sonidos (parecidos a los de un trombón) con la finalidad de comunicarse con dinosaurios de su misma especie. Una función que también se cree posible es la atracción de machos a hembras utilizando la cresta, esto se cree debido a que los machos poseén una cresta más larga que las de las hembras.
La cresta era hueca, con tubos separados que van de cada ventana de la nariz al extremo de la cresta para dar vuelta en la dirección contraria y dirigirse atrás y abajo de la cresta y el cráneo. Los tubos eran más simples en P. walkeri, y más complejos en P. tubicen, donde estaban ocultos y otros tubos cruzados y separados.[14] Como P. walkeri y P. tubicen tenía crestas largas con solamente una curvatura leve, P. cyrtocristatus tenia una cresta corta con un perfil más circular.[15]
Historia
 El especímen tipo de Parasaurolophus walkeri en el Museo Real de Ontario.
El especímen tipo de Parasaurolophus walkeri en el Museo Real de Ontario.Fue descrito por primera vez en 1922 por William Parks, se basa ROM 768, un cráneo y esqueleto parcial al que le faltan la mayor parte de la cola, las patas traceras distal de la articulación fémoro-tibio-rotuliana, que fue encontrado por una expedición de la Universidad de Toronto en 1920 cerca de Sand Creek e las riveras del Río Red Deer en Alberta, Canadá.[6] Estas rocas hoy se las data en el Campaniano Cretácico Superior]] siendo parte de la Formación Dinosaur Park. Su nombre significa "cercano al lagarto crestado", el nombre Parasaurolophus deriva del griego para/παρα "junto" o "cerca", saurus/σαυρος "lagarto" y lophos/λοφος "cresta".[1] William Parks llamo al especímen P. walkeri en honor a Sir Byron Edmund Walker, Presidente de la junta de administración del Museo Real de Ontario.[6] Los restos de Parasaurolophus son raros en Alberta,[16] con solo otro cráneo de un individuo juvenil, posiblemente de esta misma formación o de la Formación Oldman,[17] y tres posibles especímenes sin cráneos de la Formación Dinosaur Park.[18] En ciertas listas de faunas, se hace mención de material de P. walkeri en la Formación Hell Creek de Montana, una unidad rocosa datada en finales de Maastrichtiano.[19] [20] Esta situación no fue tomada en cuenta por Sullivan y Williamson en su revisión del género en 1999,[14] y no se ha profundizado sobre el tema en otra parte.
En 1921, Charles H. Sternberg recobro un cráneo parcial (PMU.R1250) de la levemente más joven Formación Kirtland en el Condado de San Juan, Nuevo México. Este espécimen se encuentra en Uppsala, Suecia, donde Carl Wiman lo describio como una segunda especie, P. tubicen, en 1931.[21] El epíteto especifico probiene del latín tǔbǐcěn "trompetista".[22] Un segundo cráneo, casi completo P. tubicen (NMMNH P-25100) que fuera encontrado en Nuevo México en 1995. Usando tomografía computada en este cráneo, Robert Sullivan y Thomas Williamson le dieron a este género una monografía muy completa 1999, cubriendo aspectos de su anatomía y taxonomía, y las funciones de su cresta.[14] Williamson publicó posteriormente una revisión independiente de los restos, discrepando con las conclusiones taxonómicas.[23]
John Ostrom describio otro buen ejemplar (FMNH P27393) proveniente de Nuevo México P. cyrtocristatus ein 1961. Incluye un cráneo parcial con una cresta redondeada y corta, y mucho material postcraneal del esqueleto a excepción de los pies, cuello, y de piezas de la cola.[15] El nombre especifico deriva del latín curtus "corto" y cristatus "cresta".[22] Este especimen fue hallado en la Formación Fruitland o, más probable, la base que cubre la formación Kirtland.[14] El rango de distribución de esta especie se amplio en 1979, cuando David B. Weishampel y James A. Jensen describieran un cráneo parcial con una cresta del mismo tipo en el(BYU 2467) from the Campanian-age Kaiparowits Formation of GarfCondado Garfield, Utah.[5] Además de esta, otros cráneo ha sido encontrado en Utah con la cresta de morfología corta y redondeada de P. cyrtocristatus.[14]
Especies
La especie tipo P. walkeri, de Alberta, es conocida por un solo ejemplar bien establecido.[10] Difiere de P. tubicen by por tener tubos simples en la cresta,[14] y de P. cyrtocristatus por poseer una cresta más larga y no tan redondeada y un humero más largo que el radio.[15]
P. tubicen, de Nuevo México es conocido de por lo menos los restos de tres individuos.[10] Esta es la especie más grande, con pasajes de aires mas complejos en la cresta que P. walkeri y esta en más grande y menos curvada que P. cyrtocristatus.[14]
P. cyrtocristatus, de Nuevo México y Utah, es conocido por tres posibles especimen. Es la especie más pequeña, con una cresta redondeada y corta.[14] Su tamaño pequeño y la forma de su cresta han llevado a varios científicos a sugerir que representa jóvenes o a hembras de P. tubicen, el cuál vivio en el mismo tiempo y de la misma formación. Segun lo dicho por Thomas Williamson, el materia tipo P. cyrtocristatus iaproximadamente del 72% del tamaño de P. tubicen, cerca del tamaño en el cual sobre otros lambeosaurinos se interpreta para comenzar demostrar el dimorfismo sexual en sus crestas (~70% del tamaño del adulto).[23] Esta posición se ha rechazado en revisiones recientes de lambeosaurinos.[10] [24]
Galería de cráneos

Molde del cráneo de un Parasaurolophus cyrtocristatus, Museo Field.

Molde del Cráneo de unParasaurolophus walkeri skull cast, Museo de Historia Natural de Londres.

Modelo por ordenador de los pasajes dentro de la cresta del cráneo de P. tubicen
Clasificación
Como su nombre lo indica, Parasaurolophus fue inicialmente vinculado a Saurolophus debido a las similitudes en la cresta.[6] Sin embargo, rápidamente fue asignado a la subfamilia de hadrosáuridos Lambeosaurinae mientras que Saurolophus es considerado un Hadrosaurinae.[25] Es usualmente interpretado de forma separada a otros lambeosaurinos, distinguiéndose de los hadrosáuridos de cresta en forma de casco como Corythosaurus, Hypacrosaurus y Lambeosaurus.[10] [26] [24] Su pariente más cercano parece ser Charonosaurus, un lambeosaurino con un cráneo simila, pero del que no se conoce la cresta completa, de la región del Río Amur en el noreste de China,[27] y los dos son colocado en la tribu Parasaurolophini.[24] P. cyrtocristatus, con su corta y redondeada cresta, es considerado el más filogeneticamente basal de las tres especies conocidas de Parasaurolophus,[24] o podría ser un juvenil o hembra de la especie P. tubicen.[23]
Paleoecología
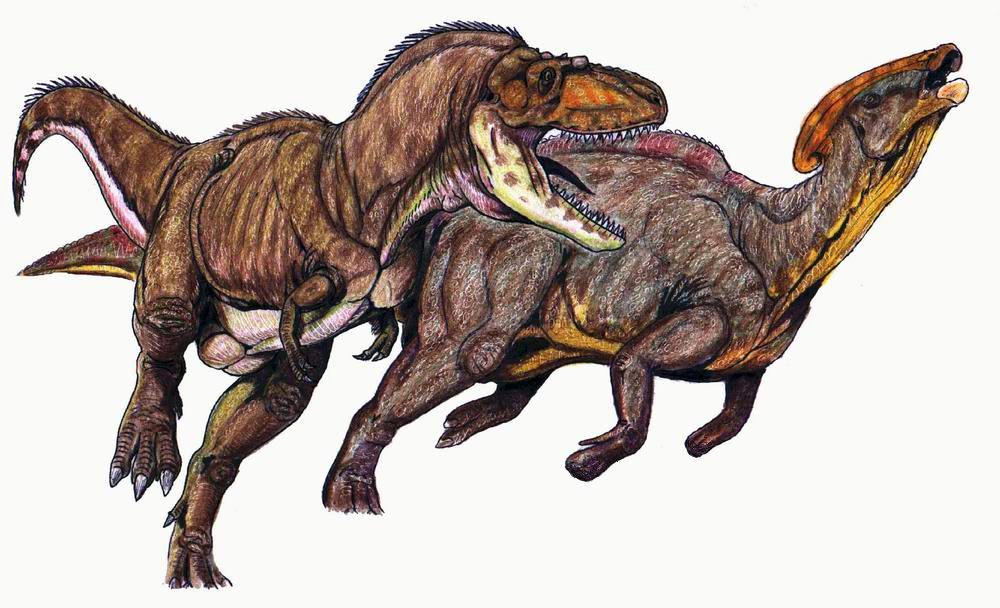
 Gorgosaurus atacando a P. cyrtocristatus.
Gorgosaurus atacando a P. cyrtocristatus.En la Formación Dinosaur Park, P. walkeri compartía el territorio con una bien documentada fauna de diinosaurios. Los más conocidos de estos son los herbívoros Centrosaurus, Styracosaurus y Chasmosaurus todos ellos dinosaurios con cuernos, Prosaurolophus, Gryposaurus, Corythosaurus, Lambeosaurus que como el Parasaurolophus eran dinosaurios de pico de pato y los dinosaurios acorazados Edmontonia y Euplocephalus. El principal predador fue el tiranosáurido Gorgosaurus, adémas se encontrava el raptor Dromaeosaurus y varios pequeños predadores más.[20] La Formación Dinosaur Park es interpretada como la sección baja de un [río] en una llanura inundable con la presencia de pantanos en influencia marina debido a la Mar interior occidental que invadia las tierras bajas en periodicas ingresiones marinas.[28] El clima era predominantemente cálido, sin estación fría pero si la presencia de temporadas secas y humedas. Las coníferas eran las plantas predominates en la región, formando la mayor parte de la dosel vegetal. Debajo de este existia un sotobosque de helechos, helechos arbóreos y plantas con flores.[29]
Las especies, P. cyrtocristatus y P. tubicen de Nuevo México compartía territorio con el gran saurópodo Alamosaurus, el hadrosáurido Kritosaurus, el ceratopsiano Pentaceratops, el anquilosáurido Nodocephalosaurus todos ellos hebívoros. Entre los principales carnívoros se encuentra a Saurornitholestes y un gran tiranosáurido que no ha sido nombrado todavia.[20] La Formación Kirtland es interpretada como una planicie ribereña, aparecida despues de que se retirara el Mar Niobraran. Coníferas eran las plantas dominates, y aparentemente los dinosaurios con cuernos erán más comunes que los de pico de pato.[30]
Paleobiología
Alimentación
Como todo hadrosáurido Parasaurolophus era un animal con la capacidad de desplazarce en forma bípeda o cuadúpeda, de alimentación hervíbora. Tenía cientos de dientes ordenados en columnas que los iba remplazando a medida que se desgastaban. Este desgaste se debía a tenía un sofisticado aparato masticatorio que le permitia moler el alimento antes de tragarlo, a diferencia de otros herbívoros de su época. El aliemnte lo tomaba con un oirgano similar a un pico y era mantenido dentro de la boca por unas estructuras similares a las mejillas de los mamiferos, que impedia que el forraje se le cayera. Este animal comía hasto los cuatro metros del suelo.[10] Segun lo notado Robert Bakker, los lambeosaurinos tenián picos mas estrechos que sus primos los hadrosaurinos, lo que podria implicar que Parasaurolophus y sus parientes pudieron ser más selectivos a la hora de tomar el alimento.[31]
Cresta craneal
Muchas hipotesis han sido propuestas para explicar las funciones de la cresta craneal del Parasaurolophus, pero muchas han sido refutadas.[8] [11] Actualmente se considera que lo más probale es que haya tenido varias funciones simultaneamente, como parte de un sistema de individualización visualde la especie y sexo, amplificación del sonido para la comunicación y termorregulación.Lo que no esta claro es cual fue el principal motor para la evolución de la cresta y de los canales internos.[13]
Hipótesis rechazadas sobre la función de la cresta
Muchas de las sugerencias tempranas se centraron en las adaptaciones para una forma de vida acuática, siguiendo la hipótesis que los hadrosáuridos eran anfibios, una línea de pensamiento común hasta los años de 1960. Así, Alfred Sherwood Romer propuso que le servia como a tubo respirador,[32] Martin Wilfarth, que era el soporte para una probóscide móvil usada como tubo para respirar o para tomar el alimento,[33] Charles M. Sternberg, que habria servido como trampa de aire para evitar que el agua llegara a los pulmones,[34] y Ned Colbert que habria sido un reservoriao de aire para prolongar su estadía debajo del agua.[35]
 Fósil de Parasaurolophus walkeri.
Fósil de Parasaurolophus walkeri.Otras sugerencias eran más físicas en naturaleza. Según lo mencionado anteriormente, W. Parks sugirió que se encontraba unida a las vértebras con ligamentos o músculos, y ayudada con el movimiento y soporte de la cabeza.[6] Othenio Abel propuso que era usada como arma para combates con otros de su especie,[36] y Andrew Milner que era usado para abrirse paso entre el follaje, como lo hace el casuario hoy en día.[11] Por otro lado también se propuso que alojaría órganos especiales. Halszka Osmólska sugiere que alojaba una glandula de sal,[37] y John Ostrom que estaba recubierta de tejido sensitivo olfatorio y mejorado sentido del olfato de los lambeosaurinos, que no tenían ninguna capacidad defensiva obvia.[38] Una hipótesis inusual, fue dada por el creacionista Duane Gish, en la que la cresta alojaria un glandulas capases de arrojar un "fuego quimico" a sus enemigos, similar a la de los escarabajos bombarderos de hoy en día.[39]
La mayor parte de estas hipótesis se han desacreditado o rechazado.[8] Por ejemplo, no hay agujero en el extremo de la cresta para una función de esnórquel. No hay cicatrices de inserciones musculares para una probóscide y es dudoso que un animal con un pico necesitara una. Como trampa de aire, no habría impedido que entrara el agua. El propuesto depósito de aire habría sido escaso para un animal el tamaño de Parasaurolophus. Otros hadrosáuridos tenían cabezas grandes sin la necesidad de crestas huecas grandes para servir como puntos inserción para los ligamentos de soporte.[38] Ningunas de las propuestas explican porqué la cresta tiene tal forma, porqué otros lambeosaurinos tienen crestas que parecen muy diferentes pero realizan una función similar o cómo los hadrosáuridos sin crestas o con una sólida consiguieron perdurar sin tales capacidades, o porqué algunos hadrosáuridos tenían crestas sólidas. Estas consideraciones afectan particularmente las hipótesis basadas en el aumento de las capacidades de los sistemas ya presentes en el animal, tal como las hipótesis de la glándula de sal y del olfato,[11] e indican que éstas no eran las funciones primarias de la cresta. Además, el trabajo sobre la cavidad nasal de los lambeosaurinos demuestra que nervio olfativo y el tejido sensorial correspondiente estaban en gran parte fuera de la porción de los pasos nasales en la cresta, así que la extensión de la cresta tenía poco que ver con el sentido del olfato.[13]
Diferencias entre especies, sexo y edad
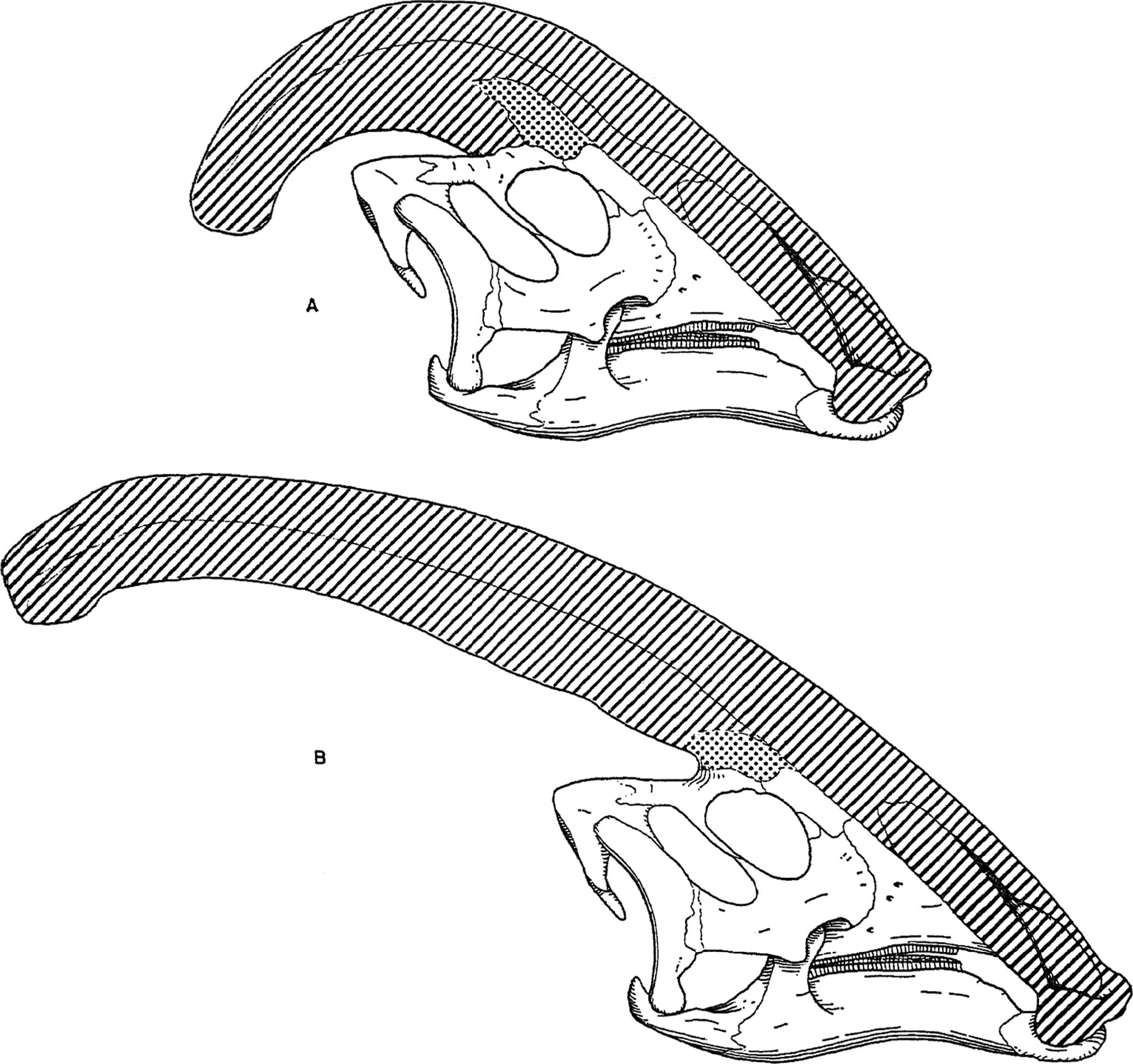
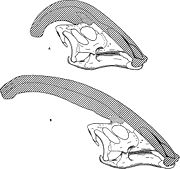 Diagrama comparativos de las de las crestas craneales de Parasaurolophus cyrtocristatus (a) y Parasaurolophus walkeri (b)
Diagrama comparativos de las de las crestas craneales de Parasaurolophus cyrtocristatus (a) y Parasaurolophus walkeri (b)Como en otros lambeosaurinos, se cree que la cresta craneal de Parasaurolophus cambiaba con la edad y en el adulto eran diferentes entre los sexos. James Hopson, uno de los primeros investigadores en describir la crestas de los lambeosaurinos en terminos de sus diferencias, sugiere que P. cyrtocristatus, con su pequeña cresta es una hembra de P. tubicen.[40] Thomas Williamson en cambio a sugerido de que se trata de una forma juvenil de este.[23] Ninguna de estas hipotesis ha sido ampliamente aceptada. Con solo seis buenos cráneos y uno juvenil, material adicional es necesario para clarificar esta situación. Williamson noto que en cualquier caso, los jovenes Parasaurolophus probablemente tuvieran, crestas redondeadas como P. cyrtocristatus, que probablemente crecieran rapidamente cuando el animal se hacercara a la madurez sexual.[23] Recientes estudios de cráneos juveniles asignados previamente a Lambeosaurus y hoy considerados Parasaurolophus, provee evidencia de una pequeña cresta tubular presente en los jovenes. This Este espécimen preserva una pequeña marca ascendente en el medio de cada hueso frontal que es similar pero menos notable que las vistas en los adultos. En estos los frontales forman una platforma que soporta la base de la cresta. Este espécimen también indica que el crecimiento de la cresta en Parasaurolophus y el perfil facial de los individuos juveniles se diferenciaba del modelo Corythosaurus-Hypacrosaurus-Lambeosaurus, en parte debido a la cresta de Parasaurolophus careciendo del hueso delgado en forma de peineta que compone la porción superior de la cresta de los otros tres lambeosaurinos.[17]
Función social
Tomando en cuenta el punto de vista, social y fisiológico se han propuesto las funciones de la cresta, tienen que ver más con la identificación visual y la comunicación sonora. Como un objeto grande, la cresta tiene un claro valor como señal visual y distingue a este animal de sus contemporáneos. El gran tamaño de la órbitas oculares y la presencia de un anillo esclerótico en los ojos implican una visión aguda y hábitos diurnos, evidencia que la vista era importante para estos animales. Si, como se ilustra comúnmente, una vela de piel se extendía desde la cresta al cuello o a la espalda, el efecto de resaltar sobre los demás habría sido incluso más llamativo.[40] Como se ha sugerido por los cráneos de los lambeosaurinos, la cresta de Parasaurolophus le permitiría identificarse de otros dinosaurios similares como Corythosaurus o Lambeosaurus y reconocer el sexo y la edad de otro de su especie por el tamaño y la forma.[13]
Función en la emisión de sonido
La apariencia externa de la cresta, bastante simple, sin embargo no corresponde a la compleja anatomía interna de los pasajes nasales, lo que sugiere que estos tendrián alguna otra función.[13] Carl Wiman fue el primero en proponer, ya en 1931, que los pasajes habrían servido para emitir un sonido identificatorio, similar a el producido por un cromorno.[21] Hopson & David B. Weishampel revisarón esat idea entre las decadas de 1970 y 1980.[40] [41] [42] Hopson encontro que hay evidencia anatómica que los hadrosáuridos tenían un oído agudo. Hay por lo menos un ejemplo, en el relacionado Corythosaurus, de un delgado estribo (hueso reptil del oído) en el lugar, que combinado con un espacio grande para el tímpano implica un sensible oído medio. Además, en los hadrosáuridos el extremo del ducto coclear es alargado como en los cocodrilianos, indicando que la porción auditiva dle oído interno estaba bien desarrollada.[40] Weishampel suguiere que P. walkeri es capaz de producir frecuencias de sonido de entre 48 a 240 Hz, y P. cyrtocristatus (interpretada como una cresta de forma juvenil) de entre 75 a 375 Hz. Basados en la similitud de los oídos internos de los hadrosáuridos con los de los cocodrilos, propuso que los adultos eran sensibles a las frecuencias altas, como las que producirían las crías. According to Weishampel, this is consistent with parents and offspring communicating.[41]
 P. cyrtocristatus en el Museo Field
P. cyrtocristatus en el Museo FieldUn modelo de ordenador de la cresta bien preservada de P. tubicen, que poseía pasajes de aires más complejos que P. walkeri, llevaron a la reconstrucción de los posibles sonidos que produciría.[43] La trayectoria principal resuena aproximadamente 30 Hz, pero la anatomía complicada de los senos causa picos y los valles en el sonido .[44]
Función en la termorregulación
El gran areas expuesta y la vascularización de la cresta ha llevado a pensar que también tendria función en la termorregulación.[12] P.E. Wheeler fue el primero en proponer este uso en 1978 como una manera de enfriar el cerebro.[45] Teresa Maryańska y Osmólska propusieron la función termorregulatoria casi al mismo tiempo,[46] y Sullivan & Williamson lo tomaron como objeto de estudio. La disertación de David Evans de 2006 sobre la función de la cresta de los lambeosaurinos, por lo menos como factor inicial para la evolución de la extensión de la cresta.[13]
Referencias
- ↑ a b Liddell, Henry George and Scott, Robert (1980). A Greek-English Lexicon (Abridged Edition). United Kingdom: Oxford University Press. ISBN 0-19-910207-4.
- ↑ a b P. walkeri en Dinodata
- ↑ P. tubicen en Dinodata
- ↑ P. cyrtocristatus en Dinodata
- ↑ a b Weishampel, David B.; and Jensen, James A. (1979). «Parasaurolophus (Reptilia: Hadrosauridae) from Utah» Journal of Paleontology. Vol. 53. n.º 6. pp. 1422–1427.
- ↑ a b c d e f g Parks, William A. (1922). «Parasaurolophus walkeri, a new genus and species of crested trachodont dinosaur» University of Toronto Studies, Geology Series. Vol. 13. pp. 1–32.
- ↑ a b c d Lull, Richard Swann; & Wright, N. E. (1942). Hadrosaurian Dinosaurs of North America. Geological Society of America, p. 209-213.
- ↑ a b c d e Glut, Donald F. (1997). «Parasaurolophus», Dinosaurs: The Encyclopedia. Jefferson, North Carolina: McFarland & Co, pp. 678–684. ISBN 0-89950-917-7.
- ↑ Brett-Surman, Michael K.; and Wagner, Jonathan R. (2006). «Appendicular anatomy in Campanian and Maastrichtian North American hadrosaurids», Carpenter, Kenneth (ed.) (ed.). Horns and Beaks: Ceratopsian and Ornithopod Dinosaurs. Bloomington and Indianapolis: Indiana University Press, pp. 135–169. ISBN 0-253-34817-X.
- ↑ a b c d e f Horner, John R.; Weishampel, David B.; and Forster, Catherine A (2004). «Hadrosauridae», Weishampel, David B.; Osmólska, Halszka; and Dodson, Peter (eds.) (ed.). The Dinosauria, 2nd edición, Berkeley: University of California Press, pp. 438–463. ISBN 0-520-24209-2.
- ↑ a b c d Norman, David B. (1985). «Hadrosaurids II», The Illustrated Encyclopedia of Dinosaurs: An Original and Compelling Insight into Life in the Dinosaur Kingdom. New York: Crescent Books, pp. 122-127. ISBN 0-517-468905.
- ↑ a b Sullivan, Robert M.; and Williamson, Thomas E. (1996). «A new skull of Parasaurolophus (long-crested form) from New Mexico: external and internal (CT scans) features and their functional implications» Journal of Vertebrate Paleontology. Vol. 16. n.º 3, Suppl.. pp. 68A.
- ↑ a b c d e f Evans, David C. (2006). «Nasal cavity homologies and cranial crest function in lambeosaurine dinosaurs» Paleobiology. Vol. 32. n.º 1. pp. 109-125.
- ↑ a b c d e f g h Sullivan, Robert M.; and Williamson, Thomas E. (1999). A new skull of Parasaurolophus (Dinosauria: Hadrosauridae) from the Kirtland Formation of New Mexico and a revision of the genus. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science, pp. 1–52.
- ↑ a b c Ostrom, John H. (1961). «A new species of hadrosaurian dinosaur from the Cretaceous of New Mexico» Journal of Paleontology. Vol. 35. n.º 3. pp. 575–577.
- ↑ Ryan, Michael J.; and Evans, David C. (2005). «Ornithischian Dinosaurs», Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Bloomington: Indiana University Press, pp. 312–348. ISBN 0-253-34595-2.
- ↑ a b Evans, David C.; Reisz, Robert R.; and Dupuis, Kevin (2007). «A juvenile Parasaurolophus braincase from Dinosaur Provincial Park, Alberta, with comments on crest ontogeny in the genus» Journal of Vertebrate Paleontology. Vol. 27. n.º 3. pp. 642–650. DOI [642:AJPOHB2.0.CO;2 10.1671/0272-4634(2007)27[642:AJPOHB]2.0.CO;2].
- ↑ Currie, Phillip J; and Russell, Dale A. "Geographic and stratigraphic distribution of dinosaur remains" in Dinosaur Provincial Park, p. 553.
- ↑ Weishampel, David B. (1990). "Dinosaur Distribution", in The Dinosauria (1st), pp. 63–139.
- ↑ a b c Weishampel, David B.; Barrett, Paul M.; Coria, Rodolfo A.; Le Loeuff, Jean; Xu Xing; Zhao Xijin; Sahni, Ashok; Gomani, Elizabeth, M.P.; and Noto, Christopher R. (2004). "Dinosaur Distribution", in The Dinosauria (2nd), pp. 517–606.
- ↑ a b Wiman, Carl (1931). «Parasaurolophus tubicen, n. sp. aus der Kreide in New Mexico» (en German). Nova Acta Regia Societas Scientarum Upsaliensis, series 4. Vol. 7. n.º 5. pp. 1–11.
- ↑ a b Simpson, D.P. (1979). Cassell's Latin Dictionary, 5 edición, London: Cassell Ltd., p. 883. ISBN 0-304-52257-0.
- ↑ a b c d e Williamson, Thomas E. (2000). «Review of Hadrosauridae (Dinosauria, Ornithischia) from the San Juan Basin, New Mexico», Lucas, S.G.; and Heckert, A.B. (eds.) (ed.). Dinosaurs of New Mexico. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science, pp. 191–213.
- ↑ a b c d Evans, David C.; and Reisz, Robert R. (2007). «Anatomy and relationships of Lambeosaurus magnicristatus, a crested hadrosaurid dinosaur (Ornithischia) from the Dinosaur Park Formation, Alberta» Journal of Vertebrate Paleontology. Vol. 27. n.º 2. pp. 373–393. DOI [373:AAROLM2.0.CO;2 10.1671/0272-4634(2007)27[373:AAROLM]2.0.CO;2].
- ↑ Gilmore, Charles W. (1924). «On the genus Stephanosaurus, with a description of the type specimen of Lambeosaurus lambei, Parks» Canada Department of Mines Geological Survey Bulletin (Geological Series). Vol. 38. n.º 43. pp. 29–48.
- ↑ Weishampel, David B.; and Horner, Jack R. (1990). «Hadrosauridae», Weishampel, David B.; Osmólska, Halszka; and Dodson, Peter (eds.) (ed.). The Dinosauria, 1st edición, Berkeley: University of California Press, pp. 534–561. ISBN 0-520-06727-4.
- ↑ Godefroit, Pascal; Shuqin Zan; and Liyong Jin (2000). «Charonosaurus jiayinensis n. g., n. sp., a lambeosaurine dinosaur from the Late Maastrichtian of northeastern China» Compte Rendus de l'Academie des Sciences, Paris, Sciences de la Terre et des planètes. Vol. 330. pp. 875–882. DOI 10.1016/S1251-8050(00)00214-7.
- ↑ Eberth, David A. 2005. "The geology", in Dinosaur Provincial Park, pp. 54-82.
- ↑ Braman, Dennis R., and Koppelhus, Eva B. 2005. "Campanian palynomorphs", in Dinosaur Provincial Park, pp. 101-130.
- ↑ Russell, Dale A. (1989). An Odyssey in Time: Dinosaurs of North America. Minocqua, Wisconsin: NorthWord Press, pp. 160–164. ISBN 1-55971-038-1.
- ↑ Bakker, Robert T. (1986). The Dinosaur Heresies: New Theories Unlocking the Mystery of the Dinosaurs and their Extinction. New York: William Morrow, p. 194. ISBN 0-8217-2859-8.
- ↑ Romer, Alfred Sherwood (1933). Vertebrate Paleontology. University of Chicago Press, p. 491.
- ↑ Wilfarth, Martin (1947). «Russeltragende Dinosaurier» (en German). Orion (Munich). Vol. 2. pp. 525–532.
- ↑ Sternberg, Charles M. (1935). «Hooded hadrosaurs of the Belly River Series of the Upper Cretaceous» Canada Department of Mines Bulletin (Geological Series). Vol. 77. n.º 52. pp. 1–37.
- ↑ Colbert, Edwin H. (1945). The Dinosaur Book: The Ruling Reptiles and their Relatives. New York: American Museum of Natural History, Man and Nature Publications, 14, p. 156.
- ↑ Abel, Othenio (1924). «Die neuen Dinosaurierfunde in der Oberkreide Canadas» (en German). Jarbuch Naturwissenschaften. Vol. 12. n.º 36. pp. 709–716. DOI 10.1007/BF01504818.
- ↑ Osmólska, Halszka (1979). «Nasal salt glands in dinosaurs» Acta Palaeontologica Polonica. Vol. 24. pp. 205–215.
- ↑ a b Ostrom, John H. (1962). «The cranial crests of hadrosaurian dinosaurs» Postilla. Vol. 62. pp. 1–29.
- ↑ Gish, Duane T. (1992). Dinosaurs by Design. Green Forest: Master Books, p. 82. ISBN 0-89051-165-9.
- ↑ a b c d Hopson, James A. (1975). «The evolution of cranial display structures in hadrosaurian dinosaurs» Paleobiology. Vol. 1. n.º 1. pp. 21–43.
- ↑ a b Weishampel, David B. (1981). «Acoustic analyses of potential vocalization in lambeosaurine dinosaurs (Reptilia:Ornithischia)» Paleobiology. Vol. 7. n.º 2. pp. 252–261.
- ↑ Weishampel, David B. (1981). «The nasal cavity of lambeosaurine hadrosaurids (Reptilia:Ornithischia): comparative anatomy and homologies» Journal of Paleontology. Vol. 55. n.º 5. pp. 1046–1057.
- ↑ Sandia National Laboratories (1997-12-05). «Scientists Use Digital Paleontology to Produce Voice of Parasaurolophus Dinosaur». Consultado el 2007-06-30.
- ↑ Diegert, Carl F.; and Williamson, Thomas E. (1998). «A digital acoustic model of the lambeosaurine hadrosaur Parasaurolophus tubicen» Journal of Vertebrate Paleontology. Vol. 18. n.º 3, Suppl.. pp. 38A.
- ↑ Wheeler, P.E. (1978). «Elaborate CNS cooling structure in large dinosaurs» Nature. Vol. 275. pp. 441–443. DOI 10.1038/275441a0.
- ↑ Maryańska, Teresa; and Osmólska, Halszka (1979). «Aspects of hadrosaurian cranial anatomy» Lethaia. Vol. 12. pp. 265–273. DOI 10.1111/j.1502-3931.1979.tb01006.x.
Véase también
- Anexo: Géneros válidos de dinosaurios
- Saurolophus
- Parque Jurásico
- Cretácico
- Hadrosauridae
Enlaces externos
 Portal:Dinosaurios Contenido relacionado con Dinosaurios.
Portal:Dinosaurios Contenido relacionado con Dinosaurios.
 Wikiespecies tiene un artículo sobre Parasaurolophus.
Wikiespecies tiene un artículo sobre Parasaurolophus.
 Wikimedia Commons alberga contenido multimedia sobre Parasaurolophus.
Wikimedia Commons alberga contenido multimedia sobre Parasaurolophus.- Los científicos usan la Paleontología Digital para producir voces de Parasaurolophus (Inglés)
- Restauraciones de P. walkeri y P. cyrtocristatus (Inglés)
- Parasaurolophus en duiops.net
- Dinosauriomanía
Categorías: Fósiles | Hadrosauroidea | Dinosaurios del Cretácico | Dinosaurios norteamericanos








{kind=link}
Wikimedia foundation. 2010.