- Succinil-CoA sintetasa
-
Succinil-CoA sintetasa (GDP) subunidad α Otros nombres Succinato-CoA ligasa (GDP) subunidad α HUGO 11449 Símbolo SUCLG1 Datos genéticos Código de gen SUCLG1 Tipo de gen Gen Codificante Locus Cr. 2 p11.3 Estructura/Función proteica Tamaño 346 (aminoácidos) Bases de datos Número EC 6.2.1.4 Entrez 8802 OMIM 611224 RefSeq NM_003849 UniProt P53597 La succinil-CoA sintetasa (SCS) o succinato-CoA ligasa es una enzima que cataliza la reacción reversible desde succinato a succinil-CoA. Para la realización de esta reacción consume un nucleótido-trifosfato (ATP o GTP). Por ello se distinguen dos enzimas diferentes:
- Formadora de GDP (EC 6.2.1.4): succinato + CoA + GTP
 succinil-CoA + fosfato + GDP
succinil-CoA + fosfato + GDP
- Formadora de ADP (EC 6.2.1.5): succinato + CoA + ATP succinil-CoA + fosfato + ATP
Esta enzima juega un papel importante como uno de los catalizadores que participan en el ciclo de Krebs, una ruta metabólica central en el metabolismo celular. La enzima se localiza en la matriz mitocondrial de la célula y su actividad óptima se consigue a una temperatura de 37 C y a un pH comprendido entre 7.0 y 8.0.
Contenido
Mecanismo enzimático
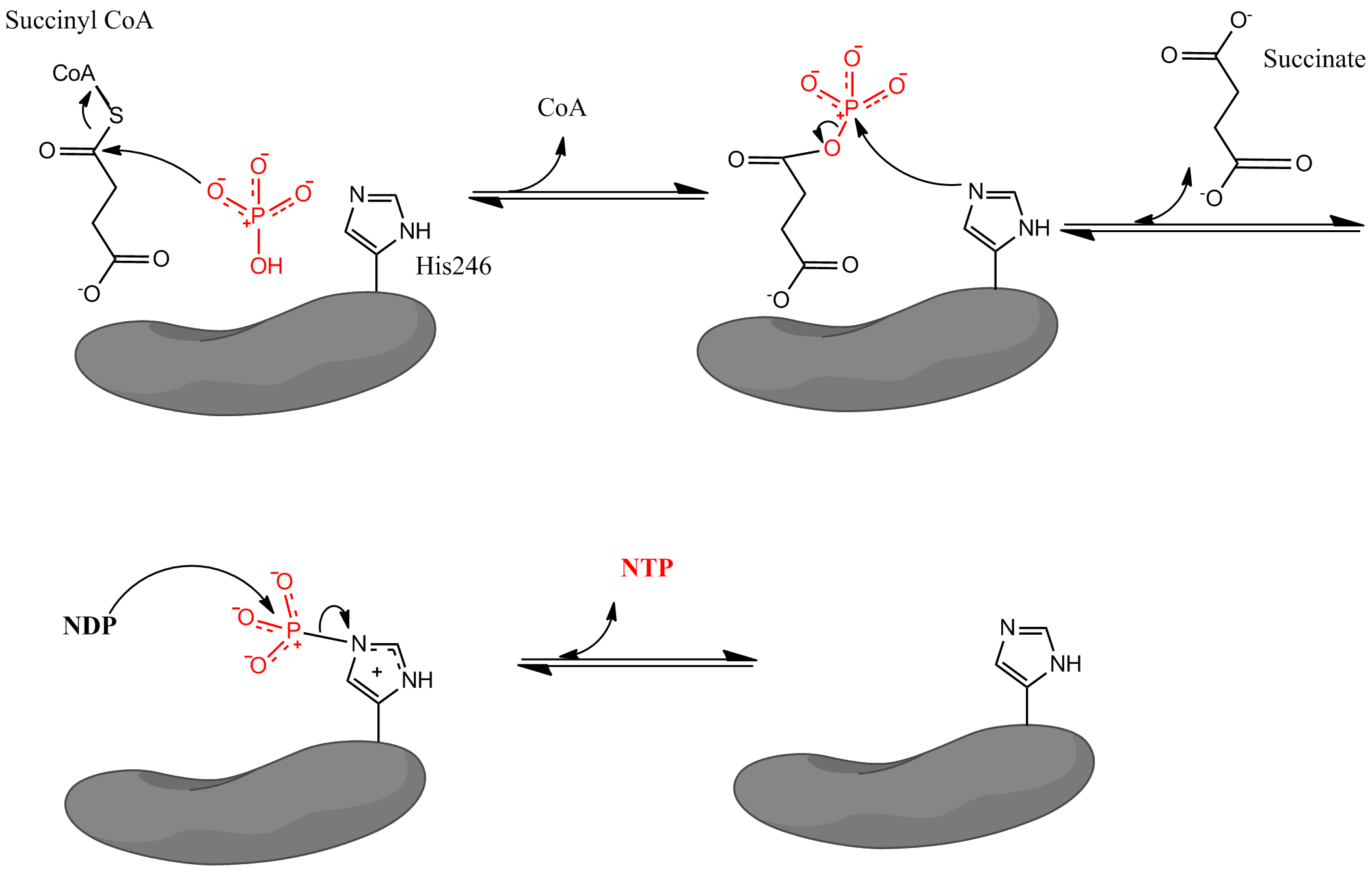
La reacción catalizada tiene lugar mediante un mecanismo de tres etapas. Se describe aquí el proceso desde succinil-CoA a succinato. Ver figura 1.
1.- La primera etapa es el desplazamiento de la CoA desde la succinil-CoA por una molécula nucleofílica de fosfato inorgánico para formar succinil fosfato.
2.- La enzima entonces utiliza un residuo de histidina para eliminar el grupo fosfato de la succinil-CoA y generar succinato.
3.- Finalmente, la histidina fosforilada transfiere el grupo fosfato a un nucleótido difosfato, generándose el nucleótido trifosfato.
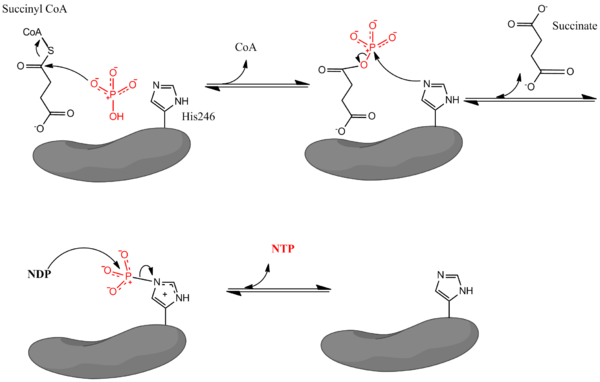 Figura 1. Mecanismo catalítico de la succinil-CoA sintetasa.
Figura 1. Mecanismo catalítico de la succinil-CoA sintetasa.
Estructura
Subunidades
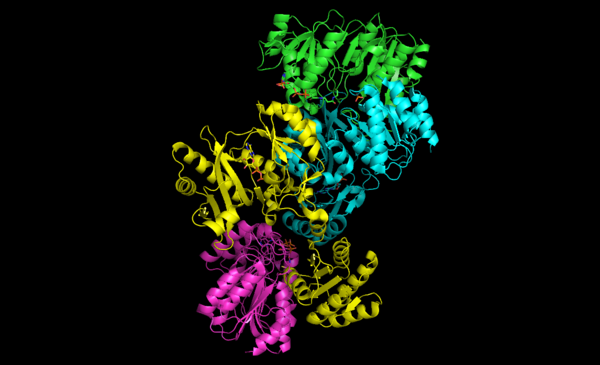 Figura 2. Heterotetrámero de la succinil-CoA sintetasa. En rosa y verde subunidades α, y en amarillo y azul subunidades β. Las subunidades rosa y amarillo forman un dímero. Las subunidades verde y azul forman el otro dímero.
Figura 2. Heterotetrámero de la succinil-CoA sintetasa. En rosa y verde subunidades α, y en amarillo y azul subunidades β. Las subunidades rosa y amarillo forman un dímero. Las subunidades verde y azul forman el otro dímero.Succinil-CoA sintetasa (GDP) subunidad β Otros nombres Succinato-CoA ligasa (GDP) subunidad β HUGO 11450 Símbolo SUCLG2 Datos genéticos Código de gen SUCLG2 Tipo de gen Gen Codificante Locus Cr. 3 p14.3 Estructura/Función proteica Tamaño 432 (aminoácidos) Bases de datos Número EC 6.2.1.4 Entrez 8801 OMIM 603922 RefSeq NM_001177599 UniProt Q96I99 Succinil-CoA sintetasa (ADP) subunidad β Otros nombres Succinato-CoA ligasa (ADP) subunidad β HUGO 11448 Símbolo SUCLA2 Datos genéticos Código de gen SUCLA2 Tipo de gen Gen Codificante Locus Cr. 13 q12.2-q13.3 Estructura/Función proteica Tamaño 463 (aminoácidos) Bases de datos Número EC 6.2.1.5 Entrez 8803 OMIM 603921 RefSeq NM_003850 UniProt Q9P2R7 Las succinil-CoA sintetasas (SCS) de los mamíferos y bacterianas están formadas por subunidades α y β. En la Escherichia coli dos heterodímeros αβ se unen para formar una estructura heterotetramérica α2β2. Al contrario, las SCS mitocondriales de los mamíferos son activas como dímeros αβ y por tanto no forman un heterotetrámero.
Tal y como se observa en la figura 2, las dos subunidades α (rosa y verde) se situan en lados opuestos de la estructura y las dos subunidades β (amarillo y azul) interactúan en el centro de la proteína. Las dos subunidades α solamente interactúan con una unidad β, mientras que las unidades β interactúan con una sola unidad α (para formar un dímero αβ) y la unidad β del otro dímero αβ. Una cadena corta de aminoácidos une las dos subunidades β para formar la estructura tetramérica.
Residuos catalíticos
Las estructuras cristalinas de la succinil-CoA sintasa de la Escherichia coli proporcionan la evidencia de que la coenzima A se une con cada subunidad α (en un plegamiento de Rossmann) en proximidad con un residuo de histidina (His-246-α). Este residuo de histidina es fosforilado durante la etapa de formación del succinato. La localización exacta de la unión del succinato no está bien definida. La formación del nucleótido trifosfato ocurre en un dominio localizado cerca del N-terminal de cada subunidad β. Este dominio está localizado a unos 35 amstrong del residuo fosforilado de histidina. Este hecho hace creer a los investigadores que la enzima debe realizar un cambio conformacional importante para llevar el residuo histidina al dominio de formación del nucleótido trifosfato para facilitar la formación de éste. Experimentos de mutagénesis han determinado que dos residuos de glutamato (uno cerca de la histidina catalítica, Glu-208-α, y otro cerca del dominio de formación del nucleótido trifosfato, Glu-179-β) juegan un papel en la fosforilación y defosforilación de la histidina, pero el mecanismo exacto por el que la enzima cambia de conformación no se conocen completamente.
Isoformas
Existen dos isoformas de succinil-CoA sintetasa en los mamíferos, una que usa ATP y otra que usa GDP. La forma GTP es la más usada en el ciclo de Krebs humano.
Funciones biológicas
Generación de nucleótidos trifosfato
La succinil-CoA sintetasa es la única enzima del ciclo de Krebs que cataliza la reacción en que un nucleótido trifosfato es formado por una fosforilación a nivel de sustrato. Estudios de investigación han mostrado que la SCS de la Escherichia coli puede catalizar tanto la formación de ATP y de GTP. En cambio, los mamíferos poseen diferentes tipos de SCS que son específicos para GTP y ATP, y son nativos en diferentes tejidos en el organismo. Un estudio interesante que utiliza células de paloma muestra que la SCS específica para el GTP está localizada en las células hepáticas, y que la SCS específica para el ATP está localizada en las células del músculo del pecho. Investigaciones posteriores revelaron un fenómeno similar en tejidos de ratas y humanos. Los tejidos que participan en el metabolismo anabólico (higado, riñones) expresan SCS-GTP, mientras que los tejidos que participan en el metabolismo catabólico (cerebro, corazón, músculo) expresan SCS-ATP.
Formación de intermedios metabólicos
La succinil-CoA sintetasa facilita el flujo de moléculas en otras rutas metabólicas controlando la interconversión entre succinil-CoA y succinato. Esto es importante ya que la succinil-CoA es un intermedio necesario para la síntesis de porfirina, grupo hemo y cuerpos cetónicos.
Regulación e inhibición
La investigación en la regulación de la succinil-CoA sintetasa en la Escherichia coli ha mostrado que la enzima es regulada a nivel transcripcional. Se ha desmostrado que el gen para la SCS es transcribida con el gen de la α-cetoglutarato deshidrogenasa bajo el control de un promotor llamado sdhC, que es parte del operón de la succinato deshidrogenasa. Este operón es activado por la presencia de oxígeno y responde a una variedad de fuentes de carbono. Los fármacos antibacterianos que previenen la fosforilación de la histidina, como la molécula Y26650, son inhibidores potentes de las SCS bacteriana.
Enfermedades relacionadas
El defecto en succinil-CoA sintetasa es causa de la acidosis láctica fatal infantil. Ésta es una enfermedad que se caracteriza por el crecimiento hasta niveles tóxicos del ácido láctico. Cuando es muy severa provoca la muerte entre 2 y 4 días despues del nacimiento. Se ha determinado que los pacientes con esta condición muestran el borrado de un par de bases en el gen SUCLG1 que codifica la subunidad α de la SCS. Como resultado, no existe SCS funcional en el metabolismo causando que las células no tengan un ciclo de Krebs funcional y la acidosis ocurra porque las células están obligadas a elegir la producción de ácido láctico para producir ATP.
Enlaces externos
Categorías:- Genes del cromosoma 2
- Genes del cromosoma 3
- Genes del cromosoma 13
- Ligasas
- Formadora de GDP (EC 6.2.1.4): succinato + CoA + GTP

Wikimedia foundation. 2010.