- Haplogrupo E ADN-Y
-
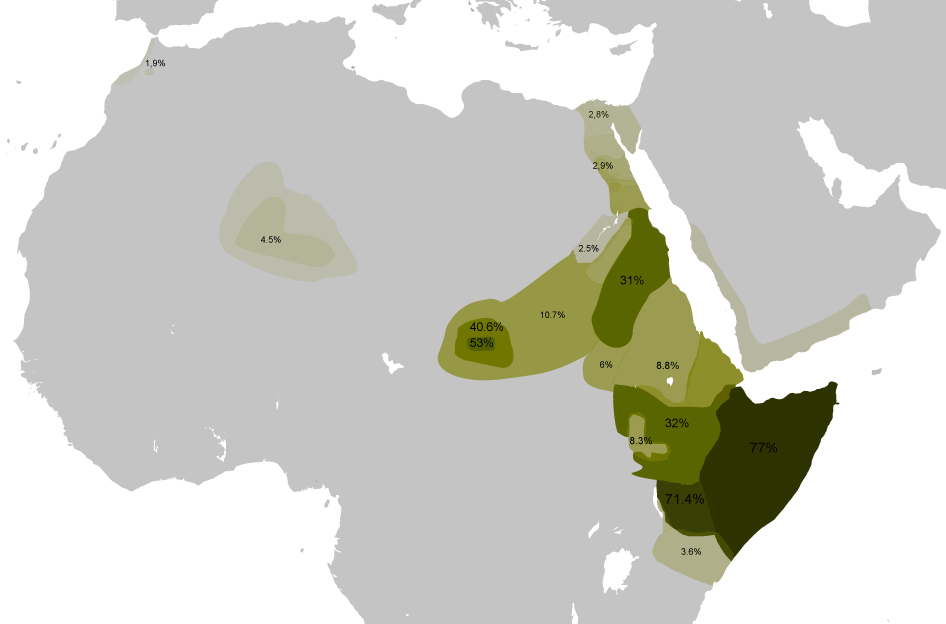
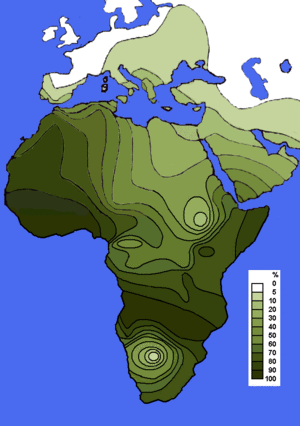 Distribución del Haplogrupo E-ADN-Y según Semino et al.2004[1] y otros autores.
Distribución del Haplogrupo E-ADN-Y según Semino et al.2004[1] y otros autores.
En genética humana, el haplogrupo E es un haplogrupo del cromosoma Y humano formado a partir de muchos haplotipos, el más conocido de los cuales es M96 y al igual que el haplogrupo D, desciende de DE. Este haplogrupo es el más característico de toda África y se encuentra también, aunque en menor proporción, en el Cercano Oriente y en Europa, especialmente en el área del Mediterráneo.
Contenido
Origen
Probablemente tiene un origen en África Oriental, de una población que generó la expansión fuera de África,[2] además en esta región está la mayor diversidad,[1] originándose en una época en que el Sahara era habitable, hace 50 o 55.000 años.[3]
Origen y antigüedad por subclados
Los subclados más frecuentes son E1b1a1 y E1b1b1. E1b1a1 se origina hace unos 20.000 años y se desarrolla en el África Occidental conjuntamente con las lenguas nigero-congoleñas, logrando una gran dispersión en todo África debido a la expansión bantú.
Por otro lado E1b1a1 se origina igualmente hace unos 20.000 años y se desarrolla en África Oriental, mostrando una relación parcial con las lenguas nilo-saharianas pero principalmente con las lenguas afroasiáticas, por lo que su presencia fuera de África podría estar relacionada al neolítico y por la interacción histórica del antiguo Egipto con otras culturas del Mediterráneo.
- E (L339, M40/SRY4064/SRY8299, M96, P29, P150, P152, P154, P155, P156, P162, P168, P169, P170, P171, P172, P173, P174, P175, P176): Con unos 50.000 - 55.000 años de antigüedad.[3] y el origen más probable en África Oriental[1]
- E1 (P147): 45.000-50.000 años en el África subsahariana.
- E1a (M33, M132): 40.000-45.000 años, origen africano.
- E1b (P177): 40.000 años, origen africano.[1]
- E1b1 (P2/PN2, P179, P180, P181, DYS391p): 35.000 en África Oriental.[4]
- E1b1a (L222.1, V38, V100): 20.000-30.000 años.
- E1b1a1 (M2, V43, V95): África Occidental.
- E1b1b (M215/Page40): 22,400 años[5] en el África Oriental,[1] probablemente Etiopía.
- E1b1b1 (M35)
- E1b1b1a (E-V68)
- E1b1b1a1 (E-M78) 18.600 años, Egipto.
- E1b1b1b (E-L19/V257)
- E1b1b1b1 (E-M81) 5.600 años, Norte de África.
- E1b1b1a (E-V68)
- E1b1b1 (M35)
- E1b1a (L222.1, V38, V100): 20.000-30.000 años.
- E1b1 (P2/PN2, P179, P180, P181, DYS391p): 35.000 en África Oriental.[4]
- E2 (M75, P68): 45.000-50.000 años, origen africano.
- E1 (P147): 45.000-50.000 años en el África subsahariana.
Distribución
El subclado más difundido es E1b1a, que es exclusivamente africano y tiene la mayor frecuencia en África occidental con un 80%,[6] fue ampliamente disperso debido primero a la expansión bantú y luego por la diáspora africana durante la Edad moderna.
El haplogrupo E se clasifica según ISOGG 2011 y se distribuye del siguiente modo:
E*
El paragrupo E* ha sido encontrado en pigmeos y bantúes de Gabón y Camerún (África Central).[7] Muy raramente en Sudáfrica y Arabia Saudita.
E1 (P147)
Propio de todo África y extendido hacia Arabia y el Mediterráneo.
E1a (M33, M132)
Muy extendido en África occidental, especialmente en Malí con 34%;[8] también en Burkina Faso, norte de Camerún y Senegal. En menor frecuencia en Norte África (Marruecos, Sudán, Egipto y saharauis) y en Europa (italianos y albaneses de Calabria).[1]
- E1a1 (M44) Característico de los fulani, especialmente en los de Camerún con 53%.[9] Presente en Malí.
- E1a2 (P110)
- E1a3 (L94)
- E1a4 (L133)
E1b (P177)
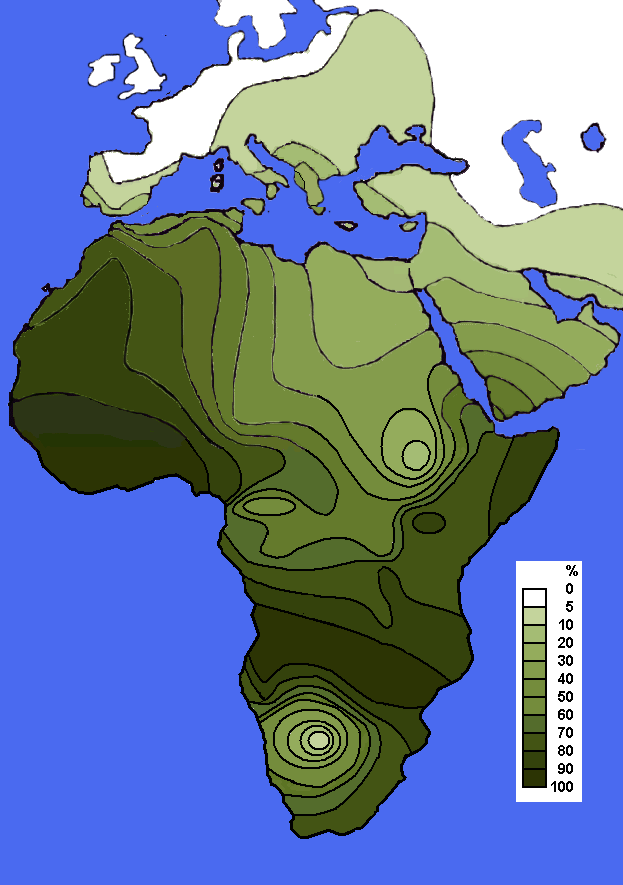
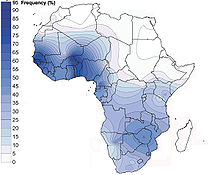 Mapa de distribución del haplogrupo E1b1a1-M2 (E3a) en Africa (Rosa et al. (2007)
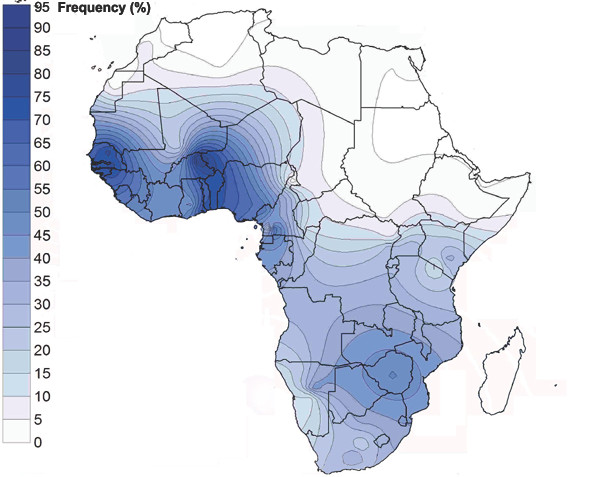
Mapa de distribución del haplogrupo E1b1a1-M2 (E3a) en Africa (Rosa et al. (2007)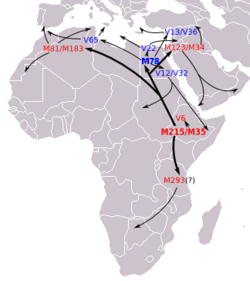 Mapa de dispersión del haplogrupo E1b1b (E3a) en Africa.
Mapa de dispersión del haplogrupo E1b1b (E3a) en Africa.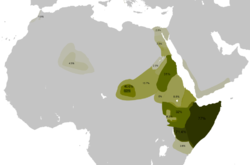 Distribución de E1b1b1a1a2 (E-V32) según Cruciani et al.2007.
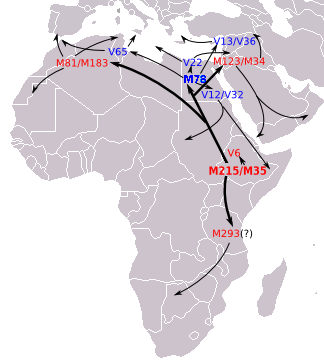
Distribución de E1b1b1a1a2 (E-V32) según Cruciani et al.2007.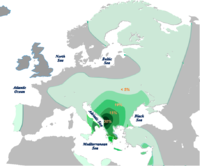 Distribución de E1b1b1a1b (E-V13) según Cruciani et al.2007.
Distribución de E1b1b1a1b (E-V13) según Cruciani et al.2007.- E1b1 (P2/PN2, P179, P180, P181, DYS391p)
- E1b1a (L222.1, V38, V100)
- E1b1a1 (DYS271/M2/SY81, V95, V43, M180/P88, P1/PN1, P46, P182, P189, P211, P293, L88.3) (antes E3a) es casi exclusivo y mayoritario en el África subsahariana, además es predominante en hablantes de lenguas nigero-congoleñas, tiene la mayor diversidad y alta frecuencia en África Occidental (80%).[6] Es también mayoritario en la población negra de América, en afroamericanos de EEUU 62%[10] y tiene menores frecuencias en Norte África, Cuerno de África y Cercano oriente. Alta frecuencia en los bamilekes con 100%,[11] mandingas 87%,[12] Senegal 81%,[13] hereros 71%[14] y mayoritario en Madagascar con 35%[15]
- E1b1a1a (M58): En hutus de Ruanda (15%),[16] Burkina faso y Sudáfrica.
- E1b1a1b (M116.2) Raro en Mali.
- E1b1a1c (M149) Raro en Sudáfrica.
- E1b1a1d (M155) Raro en Mali.
- E1b1a1e (M10, M66, M156, M195) Raro en bantúes de Tanzania y Centroáfrica. En Adamawa Camerún.
- E1b1a1f (M191, U186, U247) es el subclado de E1b1a más común. típico en Camerún, Burkina Faso, Nigeria, en pueblos bantúes, gur, igbo, joisán, afroamericanos, etc.
- E1b1a1g (U175) extendido en el África Occidental.
- E1b1a1g1a (U290): en Costa de Marfil, Benín y afroamericanos
- E1b1a1g1c (54): en los Bamileke y bakaka (Camerún)
- E1b1a1h (P268, P269) raro en Gambia
- E1b1a2 (M329): en Etiopía y Qatar
- E1b1a1 (DYS271/M2/SY81, V95, V43, M180/P88, P1/PN1, P46, P182, P189, P211, P293, L88.3) (antes E3a) es casi exclusivo y mayoritario en el África subsahariana, además es predominante en hablantes de lenguas nigero-congoleñas, tiene la mayor diversidad y alta frecuencia en África Occidental (80%).[6] Es también mayoritario en la población negra de América, en afroamericanos de EEUU 62%[10] y tiene menores frecuencias en Norte África, Cuerno de África y Cercano oriente. Alta frecuencia en los bamilekes con 100%,[11] mandingas 87%,[12] Senegal 81%,[13] hereros 71%[14] y mayoritario en Madagascar con 35%[15]
- E1b1b (M215) (antes E3b) Es el haplogrupo más común en hablantes de lenguas afroasiáticas africanas[17] (camito-semíticas), así como en muchos hablantes de lenguas nilo-saharianas y su mayor diversidad se encuentra en Etiopía. Está muy extendido en el cuerno de África, Norte de África, Sudán, bereberes, árabes y judíos askenazíes y sefardíes; también en el Sur de Europa, los Balcanes y gran parte de África subsahariana.
- E1b1b* poco en amharas de Etiopía y en Yemen
- E1b1b1 (M35)
- E1b1b1a (V68)
- E1b1b1a* encontrado en Cerdeña.
- E1b1b1a1 (L18, M78) (antes E3b1a) común en el cuerno de África, especialmente en Somalia con 78%;[18] presente también en Norte África, Cercano Oriente y sureste de Europa.
- E1b1b1a1a (V12) principalmente al sur de Egipto (44%),[5] menor frecuencia en Sudán, coptos y nubios
- E1b1b1a1a1 (M224) poco en Yemenníes
- E1b1b1a1a2 (V32) típico de hablantes de lenguas cushitas como los borana de Kenya con 71%,[19] en Somalia (52%), en oromos de Etiopía (32%) y en beyas de Sudán; también en algunos nilosaharianos como los masalit y fur de Darfur (Sudán), en menor frecuencia en otras zonas de Norte África
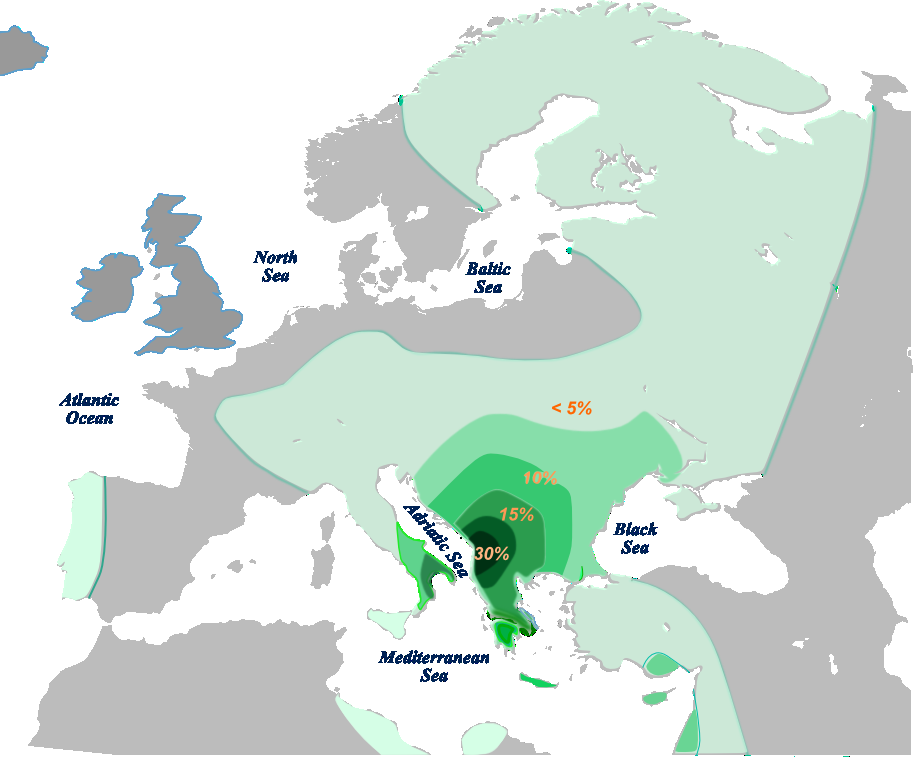
- E1b1b1a1b (V13, V36) es el único haplogrupo E que predomina en Europa en lugar de África; es típico de los Balcanes, especialmente en el Peloponeso griego (47%),[1] en albaneses de Kosovo (46%),[20] albaneses de República de Macedonia (34%), también en Grecia, Bulgaria, Rumania, Serbia y Kursk; poco en el sur de Italia, en turcochipriotas y druzos. Se estima que bajas frecuencias encontradas en Pakistán provienen de antiguos soldados griegos y otras en Gran Bretaña provienen de soldados romanos de origen balcánico.
- E1b1b1a1c (V22) principalmente en el cuerno de África y Egipto, importantes frecuencias en los fulani (31%)[12] y en los pueblos nilóticos chollo (13%) y dinka (8%); menores frecuencias al sur de Europa, Norte África y Cercano Oriente.
- E1b1b1a1d (V65) común en el Magreb,[5] principalmente en árabes de Marruecos (30%) y Libia (20%); menor frecuencia en bereberes y al sur de Europa.
- E1b1b1a1e (M521) poco en Grecia[21]
- E1b1b1a1a (V12) principalmente al sur de Egipto (44%),[5] menor frecuencia en Sudán, coptos y nubios
- E1b1b1b (L19/V257)
- E1b1b1b* encontrado en bereberes, Sudoeste de Europa y Kenia.
- E1b1b1b1 (M81) predominante en la región del Magreb (80%), especialmente en bereberes como los cabilios y otros; también en semitas como árabes y saharauis (76%);[22] común en Iberia principalmente entre pasiegos (40%)[19] y algo menos en Galicia y Portugal (12%), Andalucía (9%), Castilla y León (10%), Cantabria (16%), en Latinoamérica, y baja frecuencia en otras zonas del sur de Europa (6%, Sicilia) y Cercano Oriente (4%, Líbano y Turquía).
- E1b1b1b1a (M107)
- E1b1b1b1b (M183)
- E1b1b1c (M123) común en el Cercano Oriente y en el Mediterráneo, extendiéndose hasta el Cuerno de África.
- E1b1b1c* se ha encontrado en Omán, en israelíes de origen libio, en albaneses de Calabria (Italia) y menores frecuencias en Iberia, Egipto y Cercano Oriente.
- E1b1b1c1 (M34) importante en semitas, alta frecuencia en Etiopía y Cercano Oriente, en el área jordana del Mar Muerto (31%),[23] en Omán, Turquía, Israel (10%), Yemen (8%), en el sur de Europa (Galicia 10%, Sicilia 7%, Portugal) y en Norte África.
- E1b1b1d (V6) en los welayta 17%, en amharas 15% y en otras etnias de Etiopía y Somalia
- E1b1b1e (M293) según Henn et al.2008[24] se encuentra en etnias de Tanzania como los datooga (43%) y burunge (28%), y en pueblos khoisan como los kxoe (31%) y sandawe (24%).
- E1b1b1f (V42) en Etiopía.
- E1b1b1g (V92) en Etiopía.
- E1b1b1a (V68)
- E1b1b2 (M281) poco en oromos de Etiopía
- E1b1a (L222.1, V38, V100)
- E1b2 (P75)
E2 (M75, P68)
Es típico del África subsahariana, especialmente en Sudáfrica y en bantúes kenianos; también se encuentra en Burkina Faso, en hutus, tutsis, malgaches de Madagascar, khoisan, fon de Benín, iraqw de Tanzania, en Sudán, norte de Camerún y Senegal. Hay menores frecuencias fuera de África: en Qatar y Omán.
- E2* en bajas frecuencias en Namibia, Senegal, Zimbabue y en los mandinga (África Occidental).
- E2a (M41) con alta frecuencia en el pueblo alur de R. D. del Congo (67%).[14] Desde el África Centro-Oriental hasta Sudán, especialmente en la región de los Grandes Lagos de África.
- E2b (M54, M90, M98) Destacan los xhosa de Sudáfrica con 28%,[14] pero está extendido en toda el África Subsahariana, incluyendo Madagascar.
- E2b*
- E2b1 (M85)
- E2b1a (M200)
Adán cromosómico-Y A BT B CT DE CF D E C F G H IJK IJ K I J LT MNOPS L T M NO P S N O Q R R1 R2 R1a R1b Personaje famoso
Se ha encontrado el subclado E1b1b1b en Adolf Hitler.[25]
Enlaces externos
- Learn about Y-DNA Haplogroup E. Por Mark Lechner
- Dispersión del haplogrupo E, de la National Geographic Society
- Dispersión del haplogrupo E3a (ahora E1b1a)
- Dispersión del haplogrupo E3b (ahora E1b1b)
- Mapa de E3a (o E1b1a)
- E1b1b1g (E-M293), mapa.
- Genética poblacional del África subsahariana en haplogrupos del cromosoma Y
Referencias
- ↑ a b c d e f g Semino Ornella et al 2004, Origin, Diffusion, and Differentiation of Y-Chromosome Haplogroups E and J: Inferences on the Neolithization of Europe and Later Migratory Events in the Mediterranean Area», American Journal of Human Genetics 74 (5): 1023–1034, doi:10.1086/386295, PMID 15069642
- ↑ Chiaroni J. et al 2009, Y chromosome diversity, human expansion, drift, and cultural evolution
- ↑ a b Karafet et al. (2008), Abstract New Binary Polymorphisms Reshape and Increase Resolution of the Human Y-Chromosomal Haplogroup Tree, Genome Research, DOI: 10.1101/gr.7172008
- ↑ Trombetta, Beniamino 2011, A New Topology of the Human Y Chromosome Haplogroup E1b1 (E-P2) Revealed through the Use of Newly Characterized Binary Polymorphisms
- ↑ a b c Cruciani et al. (2007), "Tracing Past Human Male Movements in Northern/Eastern Africa and Western Eurasia: New Clues from Y-Chromosomal Haplogroups E-M78 and J-M12", Molecular Biology and Evolution 24: 1300-1311, doi:10.1093/molbev/msm049
- ↑ a b Sims et al. (2007), Sub-Populations Within the Major European and African Derived Haplogroups R1b3 and E3a Are Differentiated by Previously Phylogenetically Undefined Y-SNPs, HUMAN MUTATION Mutation in Brief #940 (2007) Online
- ↑ Gemma Berniell-Lee et al 2009, Genetic and Demographic Implications of the Bantu Expansion: Insights from Human Paternal Lineages
- ↑ Peter A. Underhill, Peidong Shen, Alice A. Lin et al., "Y chromosome sequence variation and the history of human populations," Nature Genetics, Volume 26, November 2000
- ↑ Cruciani, Fulvio et al 2002, A Back Migration from Asia to Sub-Saharan Africa Is Supported by High-Resolution Analysis of Human Y-Chromosome Haplotypes American Journal of Human Genetics 70:1197–1214, 2002.
- ↑ Hammer, Michael F. et al 2005, Population structure of Y chromosome SNP haplogroups in the United States and forensic implications for constructing Y chromosome STR databases
- ↑ Luis, J.R.; D. J. Rowold (2004 March). "The Levant versus the Horn of Africa: Evidence for Bidirectional Corridors of Human Migrations". The American Society of Human Genetics 74 (3): 532–44. doi:10.1086/382286. PMID 14973781. PMID 14973781. PMC 1182266.
- ↑ a b Rosa et al. 2007 in a study of Guinea Bissau, showed that the Fulani there are about 10% E-M78. Note that this study did not test specifically for V12 or V22, so the E-M78 may have a different exact breakdown of diversity as well as a lower frequency.
- ↑ Semino et al. (2002) "Ethiopians and Khoisan share the deepest clades of the human Y-chromosome phylogeny". Am J Hum Genet 70:265–268
- ↑ a b c Elizabeth T Wood, Daryn A Stover, Christopher Ehret et al., "Contrasting patterns of Y chromosome and mtDNA variation in Africa: evidence for sex-biased demographic processes," European Journal of Human Genetics (2005) 13, 867–876. (cf. Appendix A: Y Chromosome Haplotype Frequencies)
- ↑ Matthew E. Hurles et al 2005, The Dual Origin of the Malagasy in Island Southeast Asia and East Africa: Evidence from Maternal and Paternal Lineages
- ↑ J. R. Luis et al 2004, The Levant versus the Horn of Africa: Evidence for Bidirectional Corridors of Human Migrations
- ↑ Ehret et al. (2004), «The Origins of Afroasiatic», Science 306 (5702): 1680, doi:, http://wysinger.homestead.com/afroasiatic_-_keita.pdf
- ↑ Sanchez et al. 2005, High frequencies of Y chromosome lineages characterized by E3b1, DYS19-11, DYS392-12 in Somali malesEuropean Journal of Human Genetics 13 856–866
- ↑ a b Cruciani et al. 2004, Phylogeographic Analysis of Haplogroup E3b (E-M215) Y Chromosomes Reveals Multiple Migratory Events Within and Out Of Africa.American Journal of Human Genetics 74 1014–1022
- ↑ Peričic et al. (2005), «High-resolution phylogenetic analysis of southeastern Europe traces major episodes of paternal gene flow among Slavic populations», Mol. Biol. Evol. 22 (10): 1964–75, doi:, PMID 15944443, http://mbe.oxfordjournals.org/cgi/content/full/22/10/1964.
- ↑ Battaglia et al. 2008 "Y-chromosomal evidence of the cultural diffusion of agriculture in southeast Europe" European Journal of Human Genetics.
- ↑ Bosch et al. 2001 High-resolution analysis of human Y-chromosome variation shows a sharp discontinuity and limited gene flow between north-western Africa and the Iberian Peninsula.Am J Hum Genet 68 1019–1029
- ↑ Flores et al. 2005 "Isolates in a corridor of migrations: a high-resolution analysis of Y-chromosome variation in Jordan" J Hum Genet 50 435–441
- ↑ Henn et al. 2008 "Y-chromosomal evidence of a pastoralist migration through Tanzania to southern Africa"
- ↑ DNA genealogists surprised that Hitler's Y chromsome DNA is African
- E (L339, M40/SRY4064/SRY8299, M96, P29, P150, P152, P154, P155, P156, P162, P168, P169, P170, P171, P172, P173, P174, P175, P176): Con unos 50.000 - 55.000 años de antigüedad.[3] y el origen más probable en África Oriental[1]
Wikimedia foundation. 2010.