- Polinización en orquídeas
-
Polinización en orquídeas

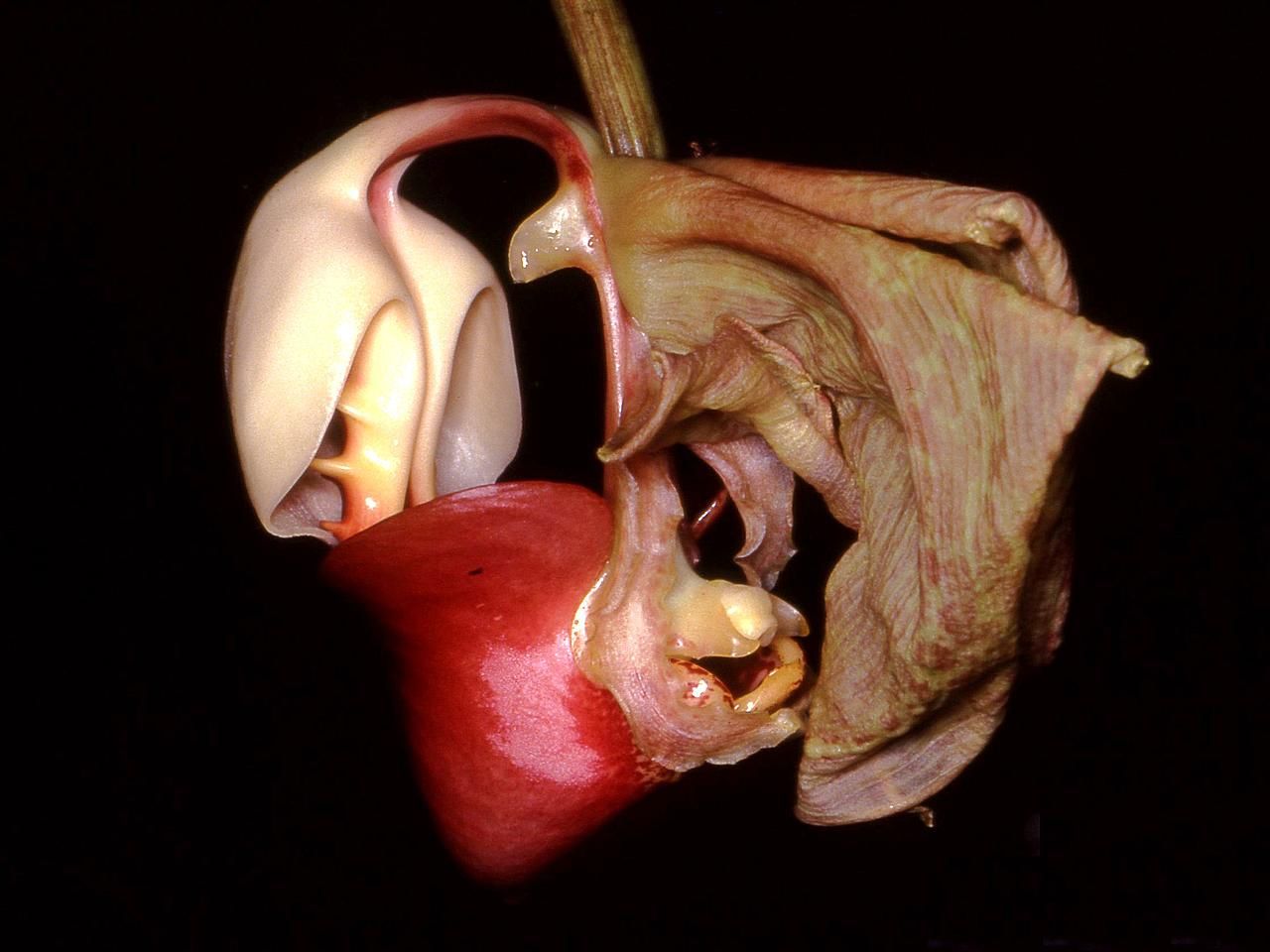
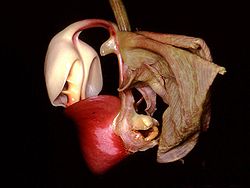 Las formas, colores y fragancias de las orquídeas son el resultado de su coevolución con los animales polinizadores. En la imagen se observa una flor Coryanthes leucocorys, especie que ha desarrollado uno de los mecanismos más fascinantes de atracción de polinizadores por medio de una bolsa llena de líquido.
Las formas, colores y fragancias de las orquídeas son el resultado de su coevolución con los animales polinizadores. En la imagen se observa una flor Coryanthes leucocorys, especie que ha desarrollado uno de los mecanismos más fascinantes de atracción de polinizadores por medio de una bolsa llena de líquido.


La polinización en las orquídeas es un capítulo extremadamente complejo en la biología de esta familia de plantas que se distinguen, por sobre las demás angiospermas, por la complejidad de sus flores y por las intrincadas interacciones ecológicas con sus agentes polinizadores. Es un tema que ha captado la atención de numerosos científicos a través del tiempo, incluyendo a Charles Darwin, padre de la teoría de la evolución por selección natural, quien realizó las primeras observaciones acerca del papel fundamental de los insectos en la polinización de las orquídeas, las que fueron publicadas en 1862 en su libro «La fecundación de las orquídeas».[a][1] De hecho, Darwin no exageró cuando afirmó que las variadas estratagemas que usan las orquídeas para atraer a sus polinizadores trasciende la imaginación de cualquier ser humano.[2]
Contenido
Adaptaciones de las orquídeas a la polinización por animales
El 97% de las especies de orquídeas necesitan de un polinizador para que se lleve a cabo la transferencia de los granos de polen de una planta a los pistilos de otro individuo y, por ende, para que se produzca la fecundación y la formación de las semillas.[3] Se debe tener en cuenta que el polen de las orquídeas se halla agrupado en masas compactas llamadas polinias (singular: "polinario"), de tal modo que por sí solo, o por acción del viento, el polen no se puede dispersar de una flor a otra por lo que los polinizadores son imprescindibles para asegurar su reproducción sexual.[3] Estos polinizadores son muy variados y, según cuál sea la especie en cuestión, pueden ser moscas, mosquitos, abejas, avispas, mariposas, coleópteros, y aves (especialmente colibríes).[4] [5] [3]
La zoofilia que caracteriza a las orquídeas presupone que los animales polinizadores visiten las flores de manera regular y se detengan en ellas el tiempo suficiente; que las anteras y el estigma sean rozados o tocados con cierta frecuencia y que el primero quede adherido a los visitantes de modo tan perfecto que pueda llegar con la debida seguridad a los estigmas de otras flores. El resultado de la zoofilia depende esencialmente de que los animales puedan reconocer las flores desde una cierta distancia y de que se vean compelidos a visitar durante un cierto tiempo las flores de la misma especie. Las flores zoófilas, entonces, deben poseer productos atractivos (cebos, como el polen y el néctar), medios de reclamo (tales como olores y colores) y, además, polen viscoso o adherente.[4]
En el curso de la evolución de las angiospermas se ha producido una diferenciación muy intensa de los medios de atracción y reclamo, así como de la forma de la flor; gracias a ello un número cada vez más grande de animales han podido colaborar en la polinización. De la visita casual de las flores por animales variados se ha pasado de modo progresivo, en el transcurso de la evolución, al establecimiento de estrechas relaciones entre los «animales antófilos » y las «flores zoófilas», con evidentes ventajas para ambos grupos. Para las plantas implicó una precisión creciente en la atracción de sólo determinados visitantes y una transferencia del polen a los estigmas de otras plantas cada vez más segura, lo que resultó en un ahorro progresivo en la producción de polen. De hecho, la relación entre el número de granos de polen y el número de óvulos producidos por una flor es del orden de un millón para las plantas anemófilas, mientras que en las orquídeas es de uno. Para los animales polinizadores especializados, la competencia con otros animales antófilos resultó disminuida y la polinización orientada o especializada en una sola especie pasó a ser, en última instancia, ventajosa para ellos.[4] [3]
El desarrollo evolutivo de las angiospermas zoófilas y de los grupos de animales que se han ido adaptando a ellas sólo puede comprenderse como una coevolución condicionada por relaciones recíprocas. La adaptación entre sí de las orquídeas y sus polinizadores ha llegado a veces tan lejos que no pueden existir el uno sin el otro.[4]
Los mecanismos de polinización son el fruto de la coevolución de las orquídeas y de sus polinizadores. Tales mecanismos son beneficiosos para ambas partes: el agente polinizador obtiene néctar de la flor de las orquídeas y ésta a su vez se beneficia con la transferencia del polen de una flor a otra. No obstante, en muchos casos la atracción de los polinizadores por las orquídeas no involucran recompensas sino engaños.[3]
Las flores de las orquídeas

 Ilustración de la polinización de Angraecum sesquipedale por una polilla hipotética de larga proboscis. El dibujo fue realizado en 1867 por Alfred Russell Wallace, siguiendo la predicción realizada por Charles Darwin de 1862. En 1903 fue descubierta la polilla en Madagascar y bautizada como Xanthopan morganii praedicta.
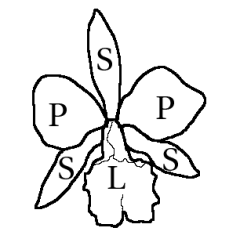
Ilustración de la polinización de Angraecum sesquipedale por una polilla hipotética de larga proboscis. El dibujo fue realizado en 1867 por Alfred Russell Wallace, siguiendo la predicción realizada por Charles Darwin de 1862. En 1903 fue descubierta la polilla en Madagascar y bautizada como Xanthopan morganii praedicta.Las flores de las orquídeas son en general hermafroditas (raramente unisexuales) y usualmente cigomorfas (es decir, de simetría bilateral). En la gran mayoría de los géneros, las flores están formadas por tres elementos externos llamados sépalos, dos laterales y uno dorsal, y tres elementos internos llamados pétalos, el inferior modificado en un labio o labelo, de mayor tamaño y de color más intenso que los restantes. El labelo a menudo es trilobulado, o de formas inusuales, con chichoncitos o crestas carnosas o con un espolón basal, y muchas veces con un patrón de colores totalmente diferente al de los otros dos pétalos.[6]
El androceo presenta usualmente uno o dos estambres (a veces 3) y está unido al estilo y al estigma, formando una "columna" (también llamada "ginostemo" o "ginostegio"). El polen comúnmente se halla agrupado en 2 o 4 (o pueden ser 1-12) masas suaves a duras llamadas polinias. La polinia junto con un tallito pegajoso (derivado de la antera o del estigma) forman en conjunto el "polinario", la unidad de transporte durante la polinización. El gineceo está formado por 3 carpelos unidos, es ínfero. El estilo y estigma se hallan altamente modificados, el estilo es solitario y terminal y es el principal componente de la "columna", con una porción del estigma formando un lóbulo alargado usualmente no receptivo, llamada "rostelo", posicionada sobre la región estigmática, y una porción del rostelo que puede formar una plataforma pegajosa, llamada "viscidio". El "viscidio" está adjuntado al tallito del "polinario".[6]
En general son productoras de néctar, como recompensa a los polinizadores. Los nectarios son variables en posición y tipo. Por ejemplo, se encuentran en un espolón del labio, o en los ápices de los sépalos, o en los septos del gineceo. Algunas especies de orquídeas son autógamas o apomícticas, es decir, no necesitan de polinizadores para producir semillas.[7]
Atracción de los polinizadores mediante recompensas
Muchas especies de orquídeas recompensan a los polinizadores con alimento (como por ejemplo, néctar, pelos alimenticios o aceites) y otros compuestos, tales como ceras, resinas y fragancias. Estas recompensas, a su vez, refuerzan la conducta de los polinizadores. No obstante, la especialización en un solo tipo de polinizador para asegurar una transferencia más eficiente de polen, determinó una creciente especialización morfológica y estructural en las flores de las orquídeas para garantizar la atracción de una sola especie de insecto.[8] [3] [9] [10]
Este punto central en la evolución de las relaciones interespecíficas entre las orquídeas y sus polinizadores fue captado por Darwin al estudiar las especies británicas y algunas especies exóticas de esta familia. Por ejemplo, la especie de Madagascar Angraecum sesquipedale posee en su flor un espolón de más de 30 cm cuyo fondo se llena de néctar. Darwin trató infructuosamente de remover las polinias de la flor usando agujas. Sólo pudo hacerlo después de introducir hasta el fondo del espolón un cilindro con un diámetro de 2,5 mm y traccionar nuevamente hacia afuera, ya que en ese momento el viscidio quedaba adherido al cilindro. Darwin razonó que cuando una mariposa intentara llegar con su trompa hasta el fondo del espolón para libar el néctar, una vez que retirase su probóscide, las polinias quedarían adheridas a su cabeza. Al visitar la siguiente flor, la misma sería polinizada ya que la mariposa estaría depositando las polinias en su estigma.[11] Este razonamiento implicaba que el insecto polinizador de Angraecum sesquipedale debería ser una mariposa con una espiritrompa de más de 30 cm de longitud, una idea que sonaba ridícula para los biólogos de la época. No obstante, la predicción de Darwin pudo ser comprobada en 1903, cuando una mariposa nocturna con una espiritrompa de ese tamaño fue descubierta en Madagascar y denominada Xanthopan morganii praedicta. El epíteto subespecífico "predicta", por supuesto, hace referencia a que su existencia fue predicha por Darwin.[3]
La mariposa es atraída hacia la flor por la fragancia que ésta despide, especialmente de noche. Cuando se acerca a la flor desenrolla su probóscide y la inserta en una hendidura del rostelo que conduce al espolón. Una vez que termina de libar el néctar de la base del espolón, levanta la cabeza mientras remueve la probóscide del espolón. Cuando realiza ese movimiento hace que el viscidio se adhiera a su cabeza o a otra parte de su cuerpo. El viscidio penetra un pequeño pedicelo (la caudícula) que lleva en su extremo las polinias. Cuando la mariposa termina de enrollar su trompa y vuela hacia otra flor, la caudícula se deshidrata, se seca, lo que determina que se modifique su ángulo con respecto al cuerpo del insecto, de modo tal que cuando éste visite la próxima flor e inserte su trompa en ella, las polinias quedarán enfrentadas, y posteriormente adheridas, al estigma. Una vez que las polinias de una planta quedan adheridas al estigma de la flor de otra planta, es decir que ha ocurrido la polinización, las flores dejan de producir fragancia y sus tépalos se marchitan poco después. No obstante, el proceso de transferencia de polen, fecundación y formación de miles de nuevos individuos ya ha quedado asegurado.[3]
Muchas orquídeas, como el caso de Angraecum sesquipedale, son polinizadas por mariposas nocturnas y, por esa razón, presentan flores de colores claros, casi blancas y producen fragancia durante la tarde o la noche. Ejemplos de este tipo de orquídeas son las especies Bonatea speciosa, Habenaria epipactidea, el género Satyrium, Disa cooperi y D. ophrydea.[12] La polinización por mariposas, por otra parte, ha evolucionado en varios géneros de orquídeas. Las flores de estas especies presentan brillantes colores y recompensan a sus polinizadores con néctar.[12]
Las especies que son polinizadas por abejas (denominadas melitófilas) frecuentemente emiten una fuerte fragancia durante el día y son de colores fuertes y luminosos. Ejemplos de estas orquídeas son Satyrium erectum y Disa versicolor. Las diferentes especies de abejas que polinizan orquídeas no sólo pueden ser recompensadas con néctar sino que, en muchos casos, también son retribuidas con aceites. Esta recompensa -un fenómeno bastante raro en plantas- se conoce para varias decenas de especies de orquídeas, entre ellas unas 55 especies sudafricanas, y las abejas la utilizan para alimentar a sus larvas. Los géneros Disperis, Pterygodium, Corycium, Ceratandra, Evotella, Satyrium y Pachites, entre otros, presentan especies que recompensan a sus polinizadores con aceites.[12]
La polinización por moscas (miofilia) es el segundo síndrome más común entre las orquídeas, con especies de 20 familias de dípteros registradas como polinizadoras.[13] Las flores emiten aromas que semejan el alimento natural o los substratos que visitan las moscas, los cuales comúnmente son materiales orgánicos en descomposición, excrementos o carroña, en búsqueda de alimento o para depositar sus huevecillos.[8] Diferentes partes florales producen olores putrescentes o dulces, comúnmente forman trampas que retienen al polinizador, además de poseer varios apéndices que los atraen y colores ya sea brillantes o que semejan carne o algún otro tinte de materia putrefacta.[14] [15] [16] [17] Las flores de Stelis hymenantha, por ejemplo, despiden un aroma intenso a mentol dulce. En la base del labelo se produce una substancia que se adhiere fuertemente a su superficie, a modo de película, semejante al néctar. Las flores de S. immersa, otra orquídea polinizada por dípteros, son también fragantes, pero con un aroma diferente, parecido al melón; y a diferencia de la primera, la sustancia pegajosa se localiza tanto en los pétalos como en la base del labelo. Los visitantes de estas dos especies son principalmente dípteros de varias familias. La mayoría de las moscas usualmente permanecen fuera de la flor ‘‘examinando’’ el líquido viscoso de la superficie externa de los pétalos. Los polinizadores en Stelis immersa son hembras de una pequeña mosca del género Megaselia (Phoridae), la cual es la única con el tamaño preciso para efectuar la transferencia de polen. Después de examinar la substancia similar al néctar, el insecto se introduce lateralmente en la flor y se posa en el labelo que se encuentra en la posición dirigida hacia abajo. Cuando esto sucede, el labelo se levanta presionando al insecto contra el viscidio del polinario y atrapándolo. Para salir, la mosca retrocede y el viscidio se adhiere a su tórax. El labelo vuelve a su posición original liberando al animal.[18]
Atracción y recompensa por medio de perfumes
Las flores de las especies de las subtribus Stanhopeinae y Catasetinae pertenecen a las más fascinantes y extravagantes de todas las orquídeas. No son de las más bellas, pero debido a su polinización tan particular a veces han creado flores bizarras con mecanismos de polinización muy extraños. Las especies de esta tribu son polinizadas exclusivamente por machos euglosinos que buscan y recolectan perfumes en estas flores. Hasta ahora no se sabe para qué los machos colectan y guardan estos perfumes; puesto que no sirven para nutrición ni protección. Se piensa que deben estar relacionados con los rituales de apareamiento de estos insectos. La manera de colectar estos perfumes es siempre muy parecida. El macho se acerca a la fuente del aroma, el osmóforo; normalmente se posa sobre el labelo y empieza a colectar con sus patas delanteras las sustancias responsables del aroma. En la mayoría de casos son líquidos; en algunas especies en forma cristalina. Si son sólidos, el macho los disuelve primero con secreciones de sus glándulas salivales. Con los pelos largos y densos de sus patas delanteras, el macho recoge el líquido presente en el labelo. Si este órgano está saturado de los componentes aromáticos, la abeja sale de la flor para transferir los aromas con la ayuda de las patas medias a cavidades que se encuentran en las patas traseras. En el interior de estas cavidades las sustancias pueden ser guardadas por mucho tiempo. Distintos géneros de orquídeas pegan los polinarios (o polinia) a diferentes partes del cuerpo de estos polinizadores que son atraídos químicamente a sus flores.[19] Las orquídeas, al igual que otras plantas, atraen selectivamente a un grupo específico de machos euglossinos por medio de la producción de mezclas de aromas especie-específicos que aparentemente actúan como mecanismos de aislamiento reproductivo. Así pues, diferentes especies de plantas atraen diferentes grupos de polinizadores. Inclusive, algunas orquídeas presentan modificaciones morfológicas en sus flores de tal manera que solo liberan el polinario cuando son visitadas por ciertas especies de abejas (según su tamaño y comportamiento). Por esta razón no todos los euglosinos que visitan una especie de orquídea son polinizadores efectivos de ésta.[20]
Atracción de los polinizadores mediante engaños
 Ophrys bombyliflora es una de las especies de orquídea que atrae a los insectos mediante la estrategia del engaño sexual: su labelo mimetiza a las hembras de su polinizador.
Ophrys bombyliflora es una de las especies de orquídea que atrae a los insectos mediante la estrategia del engaño sexual: su labelo mimetiza a las hembras de su polinizador.Muchas orquídeas han recurrido a la táctica de seducir a los polinizadores ofreciendo aromas, formas, colores o movimientos que imitan algo que a éstos les interesa sin ofrecer nada a cambio. Los mecanismos de engaño son tan variados como sorprendentes y se listan a continuación:[21] [22]
- engaño alimenticio generalizado: las flores imitan la forma y el colorido de las especies que usualmente recompensan a los polinizadores.
- engaño alimenticio mediado a través de mimetismo floral:[23] en este caso, las flores imitan exactamente a una especie particular que recompensa a los polinizadores y con la que cohabitan.[24]
- imitación de sitios de anidamiento: las flores imitan los sitios de postura de huevos de los polinizadores.
- imitación de sitios de resguardo: las flores proveen a los polinizadores de sitios para resguardarse. Esta estrategia puede no ser engañosa, sino favorable para ambos, el insecto y la orquídea.
- pseudoantagonismo: la planta atrae a los polinizadores a través de la invocación de mecanismos de defensa innatos. Así, imita la forma de otra especie de insecto, por ejemplo, que el polinizador desea alejar o matar. El polinizador, al ver a su supuesto enemigo, lo ataca, una y otra vez. En esta pelea inútil contra una flor, el insecto se cubre de polen que distribuirá a otras flores cuando sea engañado nuevamente.[25]
- atracción "rendezvous": las flores imitan a otras flores que son atractivas para las hembras de los polinizadores
- engaño sexual: en este caso, las flores mimetizan las señales de apareamiento (tanto visuales como olfativas) de las hembras de los polinizadores.
De los mecanismos descritos, el común entre las orquídeas es el de engaño alimenticio generalizado (informado en 38 géneros) seguido del engaño sexual (18 géneros).[26] [22]
Atracción de los polinizadores mediante engaño por alimento
La capacidad de atraer a los polinizadores sin ofrecerles ninguna recompensa a cambio ha evolucionado independientemente en varios linajes de angiospermas, pero usualmente en solo unas pocas especies por familia.[27] En contraste, se ha estimado que cerca de una tercera parte de las especies de orquídeas utilizan el mecanismo de engaño alimenticio.[28] [29] Este mecanismo consiste en la atracción de polinizadores mediante la señalización de la presencia de alimento, tales como néctar o polen, pero sin proveerles ningún tipo de recompensa. Para ello, las orquídeas utilizan la estrategia de parecerse a especies que si recompensan a sus polinizadores y con las cuales habitan conjuntamente.[30] Más frecuentemente, el engaño alimenticio consiste en una semejanza general con las especies que recompensan a los polinizadores, es decir, este tipo de orquídeas presenta flores grandes y de colores brillantes, estrategia que explota las preferencias innatas de los polinizadores por esa clase de flores.[29] [31] [22]
Imitación de otras plantas
En este caso la orquídea "engaña" a los polinizadores típicos de otras especies de plantas para que realicen la polinización. El método para engañarlos es imitar las flores de otras especies. Un caso muy descriptivo de esto se observa en la orquídea Epidendrum ibaguense. Esta orquídea terrestre o litófila, abunda desde México hasta Bolivia y Brasil. Sus flores de color naranja con labelo amarillo intenso imitan a las flores de una asclepiadácea, Asclepias curassavica. Existe una mariposa, Agraulis vanillae, que visita usualmente a esta especie para procurarse néctar a cambio del transporte de polen. En muchas oportunidades, no obstante, la mariposa atraída por el color y la forma de las flores de Epidendrum se dirige hacia ellas e introduce su aparato bucal (su espiritrompa) en un conducto estrecho (ginostemo) que, por su muy escaso diámetro, determina que la espiritrompa quede atrapada unos instantes. El forcejeo del insecto por liberarse hace que las polinias de la orquídea se le adhieran a su cabeza. Luego de soltarse, y al ser engañado nuevamente por otra planta de Epidendrum, transportará el polen y permitirá así la polinización en esta especie pero sin haber recibido néctar por sus servicios.[32]
Atracción de polinizadores por engaño sexual
Comparación de las concentraciones de varios compuestos en las feromonas del insecto Andrena y en la fragancia emitida por la orquídea Ophrys.[33] Compuesto Andrena Ophrys Heneicosano 1,6±0,5 1,8±0,3 Docosano 0,6±0,1 0,5±0,1 Tricosano 28,7±2,4 30,6±1,8 Tetracosano 2,0±0,2 3,1±0,2 (Z)-9-pentacoseno 3,4±1,8 0,6±0,1 Pentacosano 34,9±2,2 20,2±1,3 Hexacosano 1,6±0,1 2,1±0,2 (Z)-12 &(Z)-11-heptacoseno 0,7±0,3 6,0±0,8 (Z)-9-heptacoseno 5,1±1,6 7,6±1,0 Heptacosano 11,2±1,1 11,5±1,5 (Z)-12 &(Z)-11-nonacoseno 3,7±1,4 6,7±1,0 (Z)-9-nonacoseno 6,6±0,4 9,4±1,2 Hay flores engañosas que imitan la forma, pilosidad y olor de las hembras de ciertas avispas o abejas. El caso mejor conocido es el de Ophrys insectifera orquídea del sur de Europa; es visitada únicamente por dos especies de avispas del género Argogorytes. Los machos nacen en primavera varias semanas antes que las hembras, y en sus primeros vuelos son atraídos por la fragancia de las flores de Ophrys, similar a las feromonas secretadas por las hembras. Además, el labelo presenta forma, color y textura similar a las hembras. El proceso se designa como pseudocopulación porque las avispas macho intentan aparearse con la flor, y al hacerlo, entran en contacto con la antera, trasladando los polinios de una flor a otra en intentos sucesivos.[34]
La polinización por pseudocopulación fue descripta por primera vez en 1916 y 1917 por A. Pouyanne y H. Correvon cuando estudiaron las interrelaciones entre la orquídea Ophrys speculum y la avispa escólida Campsoscolia ciliata en Argelia.[35] [36] Las publicaciones de estos biólogos franceses pasaron inadvertidas hasta que Robert Godfrey confirmó sus observaciones en 1925, lo que determinó un incremento en el interés en el tema.[b] El trabajo de 1925 fue seguido de una gran cantidad de publicaciones de la bióloga australiana Edith Coleman acerca de la polinización de las orquídeas del género Cryptostylis por los machos de la avispa icneumonoidea Lissopimpla excelsa.[37] [38] [39]
Varios géneros de orquídeas terrestres se reproducen por este mecanismo. Los géneros más conocidos y mejor documentados son, además de los ya mencionados Ophrys y Cryptostylis, Drakaea, Caladenia,[40] Chiloglottis,[41] Geoblasta,[42] Arthrochilus, Calochilus,[43] Leporella[44] y Spiculaea. La gran mayoría de los géneros de orquídeas terrestres que utilizan la pseudocopulación se encuentran en Australia y el género más grande de orquídeas con especies pseudocopuladas es Ophrys de Europa. El mecanismo no está en absoluto circunscripto a un continente en particular, ya que también se ha informado para una especie sudamericana (Geoblasta penicillata)[45] y dos orquídeas sudafricanas del género Disa.[46]
Idéntico mecanismo utiliza Tolumnia henekeni, cuya flor semeja la hembra de una especie de abeja, Centris insularis, y lo hace tan bien que el macho de esta especie es engañado y trata de copular la imitación de hembra que le ofrece la flor. Haciendo esto poliniza la flor.
En Caleana, denominada en los países de habla inglesa "duck orchid" ("orquídea pato") por la semejanza de su labelo con la cabeza de un pato y el resto de la flor con el cuerpo de un pato en pleno vuelo, se ha descripto un mecanismo de pseudocopulación algo diferente. Esta especie usa un mecanismo de resorte para atrapa a los insectos en una bolsa, siendo su única posibilidad de escapar a través del polinio y del estigma. Los machos de los insectos polinizadores aterrizan en el labelo, lo que dispara un mecanismo de relajación de dos bisagras (una es la del labelo-lámina y la otra es la de la lámina-perianto) que da vuelta al insecto con la parte de atrás de su tórax en la bolsa que contiene el estigma y la polinia.[47] [48]
La pseudocopulación no solo está restringida a polinizadores pertenecientes al orden de los himenópteros (abejas y avispas). De hecho, ha sido descripta también para dípteros (un mosquito del género Bradysia) que pseudocopulan especies de Lepanthes, uno de los géneros más amplios de angiospermas que habitan las selvas neotropicales.[49]
Las flores de las orquídeas del género Ophrys no solo imitan la forma, el tamaño y el color de las hembras de sus polinizadores, sino que también emiten una fragancia que incluye varios compuestos que se encuentran en las feromonas sexuales de las hembras, con lo cual logran reforzar la conducta sexual de los machos. Se han comparado los compuestos volátiles emitidos por Ophrys iricolor y las feromonas femeninas de su especie polinizadora, Andrena morio, tanto desde el punto de vista químico como electrofisiológico. Se han descubierto más de 40 compuestos, entre ellos alcanos y alquenos de 20 a 29 átomos de carbono, aldehídos con 9 a 24 carbonos y dos ésteres. Casi todos esos compuestos se hallaron en similares proporciones tanto en los extractos florales de O. iricolor como en los extractos de la superficie cuticular de las hembras de A. morio, como se puede observar en la tabla de arriba.[33] Los compuestos volátiles biológicamente activos de este patrón son muy similares a los utilizados por otras especies de Ophrys que son pseudocopuladas por machos de los géneros Andrena y Colletes.[50] [51] [52] [53] [54]
Notas
a. ↑ El libro fue editado con el título "Sobre las variadas estrategias por las cuales las orquídeas británicas y foráneas son fertilizadas por insectos, y sobre los buenos efectos de la polinización cruzada" (en la edición original en inglés: On the various contrivances by which British and foreign orchids are fertilised by insects, and on the good effects of intercrossing).[1] Este libro fue la primera demostración detallada del poder de la selección natural, que podía explicar de qué modo habían surgido las interrelaciones tan complejas entre las orquídeas y sus polinizadores. En este libro, en el cual se describen las detalladas observaciones de Darwin acerca del mecanismo de polinización de varias orquídeas, sus experimentos y disecciones de flores, los cuales explicaron una gran cantidad de aspectos de la biología de la polinización de esta familia previamente ignorados, tales como el "rompecabezas" de Catasetum, que se pensaba que estaba constituida por flores de tres especies diferentes en la misma planta, y también realizó predicciones que más tarde fueron confirmadas en la naturaleza.[55]
b. ↑ Tema que pasó a denominarse "mimetismo Pouyaniano" en honor a su descubridor.[56]
Referencias citadas
- ↑ a b «Darwin Online: Fertilisation of Orchids». Consultado el 20 de marzo de 2009.
- ↑ Darwin, 1862, pp. 366
- ↑ a b c d e f g h van der Cingel, N. 2001. An atlas of orchid pollination: America, Africa, Asia and Australia. CRC Press. ISBN 90-5410-486-4, 9789054104865
- ↑ a b c d Strassburger, E. 1994. Tratado de Botánica. 8va. edición. Omega, Barcelona, 1088 p.
- ↑ Gola, G., Negri, G. y Cappeletti, C. 1965. Tratado de Botánica. 2da. edición. Editorial Labor S.A., Barcelona, 1110 p.
- ↑ a b Phillip Cribb (2001) Morphology of Orchidaceae. Em A. M. Pridgeon, P. J. Cribb, M. W. Chase, and F. N. Rasmussen eds., Genera Orchidacearum, vol. 1. Oxford University Press, Oxford, UK ISBN 0-19-850513-2.
- ↑ MRM Neiland, CC Wilcock (1998), "Fruit set, nectar reward, and rarity in the Orchidaceae", American Journal of Botany 85 (12): 1657–1657, doi:, http://www.amjbot.org/cgi/content/full/85/12/1657
- ↑ a b Pijl, L. van der y C. H. Dodson. 1969. Orchid flowers. Their pollination and evolution. University of Miami Press. Coral Gables. pp. 101-122.
- ↑ Dressler, Robert L. 1993, Phylogeny and classification of the orchid family. Cambridge University Press ISBN 0-521-45058-6, 9780521450584
- ↑ Proctor, Michael; Yeo, Peter, Lack, Andrew. 1996. The natural history of pollination. Portland, Oregon. Timber Prees. 479 p.
- ↑ Darwin, 1862, pp. 197–203
- ↑ a b c Kurzweil, H. 2000. South african orchids: reproduction and pollination. National Biodiversity Institute, South Africa. Consultado el 30 de marzo de 2009.
- ↑ Christensen, D. E. 1994. Fly pollination in the Orchidaceae. In: Arditti, J. (ed.). Orchid biology: reviews and perspectives. VI. John Wiley & Sons. Ithaca. New York. pp. 415-454.
- ↑ Chase, M. W. 1985. Pollination of Pleurothallis endosthachys. Am. Orch. Soc. Bull. 54:431-434
- ↑ Johnson, S. D. y T. J. Edwards. 2000. The structure and function of orchid pollinaria. Pl. Syst. Evol. 222: 243-269.
- ↑ Borba, E. L., J. M. Felix, V. N. Solferini y J. Semir. 2001b. Fly-pollinated Pleurothallis (Orchidaceae) species have high phenetic variability: evidence from isozyme markers. Am. J. Bot. 88: 419-428.
- ↑ Borba, E. L. y J. Semir. 2001. Pollinator specificity and convergence in fly pollinated Pleurothallis (Orchidaceae) species: a multiple population approach. Ann. Bot. 88: 75-88.
- ↑ Octavio Albores-Ortiz y Victoria Sosa. Polinización de dos especies simpátricas de Stelis (PLEUROTHALLIDINAE, ORCHIDACEAE). Acta Botanica Mexicana 74: 155-168 (2006)
- ↑ GÜNTER GERLACH. 2003.LA SUBTRIBU STANHOPEINAE: SUS NOTABLES MECANISMOS DE POLINIZACIÓN, LA QUÍMICA DE SUS AROMAS FLORALES E IMPLICACIONES EN SISTEMÁTICA Y TAXONOMÍA LANKESTERIANA 7: 104-106.
- ↑ Santiago Ramírez, Robert L. Dressler y Mónica Ospina. 2002.Abejas euglosinas (Hymenoptera: Apidae) de la Región Neotropical: Listado de especies con notas sobre su biología Biota Colombiana 3:7-118.
- ↑ Jana Jersáková, Steven D. Johnson & Pavel Kindlmann. 2006. Mechanisms and evolution of deceptive pollination in orchids. Biological Reviews (2006), 81:2:219-235
- ↑ a b c Richard J. Waterman & Martin I. Bidartondo. 2008. Deception above, deception below: linking pollination and mycorrhizal biology of orchids J. Exp. Bot. 59: 1085-1096.
- ↑ Existen dos tipos generales de mimetismo relevantes para la ecología de la polinización: el mimetismo Mulleriano y el mimetismo Batesiano. En el primer caso (que muchos autores toman como convergencia más que como mimetismo) varias especies con una abundancia similar en la naturaleza, desarrollan para su mutua ventaja un mismo "estilo de propaganda" como advertencia a depredadores, por ejemplo. En contraste, un organismo escasamente representado en la naturaleza, que imita a otro más abundante para engañar a un tercero, desarrolla una ventaja unidireccional que se denomina mimetismo Batesiano. (Proctor & Yeo, pag.: 375)
- ↑ A Dafni. 1984. Mimicry and Deception in Pollination. Annual Review of Ecology and Systematics. Vol. 15: 259-278
- ↑ Orlean, S. A Plant With Smarts. NOVA, Consultado el 28 de marzo de 2009.by
- ↑ Jana Jersáková, Steven D. Johnson & Pavel Kindlmann. 2006. Mechanisms and evolution of deceptive pollination in orchids. Biological Reviews (2006), 81:2:219-235
- ↑ Renner SS. Rewardless flowers in the angiosperms and the role of insect cognition in their evolution. In: Plant–pollinator interactions: from specialization to generalization—Waser NM, Ollerton J, eds. (2006) Chicago, IL, USA: University of Chicago Press. 123–144.
- ↑ Ackerman JD. Mechanisms and evolution of food-deceptive pollination systems in orchids. Lindleyana (1986) 1:108–113
- ↑ a b Nilsson LA. (1992) Orchid pollination biology. Trends in Ecology and Evolution 7:255–259.
- ↑ Johnson SD. Batesian mimicry in the non-rewarding orchid Disa pulchra, and its consequences for pollinator behaviour. Biological Journal of the Linnean Society (2000) 71:119–132.
- ↑ Schiestl FP. On the success of a swindle: pollination by deception in orchids. Naturwissenschaften (2005) 92:255–264.
- ↑ Alejandro Eduardo Taborda. «Polinización natural en Orquídeas» (en español). Club Peruano de Orquídeas. Consultado el 20 de marzo del 2009.
- ↑ a b Johannes Stökl, Robert Twele, Dirk H. Erdmann, Wittko Francke, Manfred Ayasse. 2007. Comparison of the flower scent of the sexually deceptive orchid Ophrys iricolor and the female sex pheromone of its pollinator Andrena morio. Chemoecology Volume 17 4):231- 233
- ↑ Wolff, T. 1950. Pollination and fertilization of the fly ophrys, Ophrys insectifera L. in Allindelille Fredskov, Denmark. Oikos 2:20–59.
- ↑ Correvon, H. & Pouyanne, A. 1916. Un curieux cas de mimetisme chez les Ophrydées. J. Soc. Nat. Horticult. France 17, 29, 41, 84.
- ↑ Pouyanne, A. 1917. La fécondation des Ophrys par les insectes. Bull Soc Hist Nat Afr Nord 1917;8:6–7
- ↑ Coleman, E. (1927) Pollination of the orchid Cryptostylis leptochila. Victorian Naturalist 44: 20–22.
- ↑ Coleman, E. (1929) Pollination of Cryptostylis subulata (Labill.) Reichb. Victorian Naturalist 46: 62–66.
- ↑ Coleman, E. (1930) Pollination of Cryptostylis erecta R.Br. Victorian Naturalist 46: 236–238
- ↑ Peakall, R., and Beattie, A. J. (1996). Ecological and genetic consequences of pollination by sexual deception in the orchid Caladenia tentaculata. Evolution 50, 2207-2220.
- ↑ Mant, J., Peakall, R., and Weston, P.H. (2005) Specific pollinator attraction and the diversification of sexually deceptive Chiloglottis (Orchidaceae). Plant Systematics and Evolution , 253, 185-200.
- ↑ Correa, M.N., 1968. Rehabilitacion del genero Geoblasta Barb Rodr. Rev. Museo La Plata n.s. 11, 69–74.
- ↑ Fordham, F., 1946. Pollination of Calochilus campestris. Victorian Nat. 62, 199–201.
- ↑ Peakall, R. (1989a) The unique pollination of Leporella fimbriata (Orchidaceae) by pseudocopulating winged male ants Myrmecia urens (Formicidae). Plant Systematics and Evolution 167, 137-148.
- ↑ Ciotek, L., P. Giorgis, S. Benitez-Vieyra, & A. A. Cocucci. 2006. First confirmed case of pseudocopulation in terrestrial orchids of South America: pollination of Geoblasta pennicillata (Orchidaceae) by Campsomeris bistrimacula (Hymenoptera, Scoliidae). Flora 201:365–369.
- ↑ Whitehead, V. B. & Steiner, K. E. 1991. Males of Podalonia canescens (Sphecidae) pollinating the orchid Disa atricapilla. Proceedings 8th Congress, Entomological Society of southern Africa, Bloemfontein, 1-4 July.
- ↑ Cady, L. 1965. Notes on the pollination of Caleana major. Orchadian 2:34.
- ↑ Stephen D. Hopper, Andrew P. Brown. (2006) Australia's wasp-pollinated flying duck orchids revised (Paracaleana: Orchidaceae). Australian Systematic Botany 19:3, 211
- ↑ MARIO A. BLANCO & GABRIEL BARBOZA. 2005. Pseudocopulatory Pollination in Lepanthes (Orchidaceae: Pleurothallidinae) by Fungus Gnats. Ann Bot 95: 763-772.
- ↑ Mant JG, Brändli C, Vereecken NJ, Schulz C, Francke W & Schiestl FP. 2005. Cuticular hydrocarbons as source of the sex pheromone in Colletes cunicularius (Hymenoptera: Colletidae) and the key to its mimicry by the sexually deceptive orchid Ophrys exaltata. Journal of Chemical Ecology 31 (8): 1765-1787.
- ↑ F. P. Schiestl; M. Ayasse, H. F. Paulus, C. Löfstedt, B. S. Hansson, F. Ibarra, W. Francke. 2000. Sex pheromone mimicry in the early spider orchid (Ophrys sphegodes): patterns of hydrocarbons as the key mechanism for pollination by sexual deception. Journal of Comparative Physiology A: Neuroethology, Sensory, Neural, and Behavioral Physiology 186:567- 574.
- ↑ Ayasse, M., F. P. Schiestl, H. F. Paulus, D. Erdmann, and W. Francke. 1997. Chemical communication in the reproductive biology of Ophrys sphegodes. Mitteilungen der Deutschen Gesellschaft fu¨r allgemeine und angewandte Entomologie 11:473–476.1
- ↑ Ayasse, M., F. P. Schiestl, H. F. Paulus, F. Ibarra, and W. Francke. 2003. Pollinator attraction in a sexually deceptive orchid by means of unconventional chemicals. Proceedings of the Royal Society B:Biological Sciences 270:517–522.
- ↑ Schiestl, F.P., Peakall, R. and Mant, J. (2004) Chemical communication in the sexually deceptive orchid genus Cryptostylis. Botanical Journal of the Linnean Society 144, 199-205.
- ↑ Darwin, Charles (2006), «Journal», en van Wyhe, John, [Darwin's personal 'Journal' (1809-1881)], Darwin Online, CUL-DAR158.1-76, http://darwin-online.org.uk/content/frameset?viewtype=side&itemID=CUL-DAR158.1-76&pageseq=1, consultado el 7 de febrero de 2009
- ↑ G Pasteur. 1982. A Classificatory Review of Mimicry Systems. Annual Review of Ecology and Systematics. Vol. 13: 169-199. (doi:10.1146/annurev.es.13.110182.001125)
Categorías: Orchidaceae | Polinización | Reproducción vegetal | Relaciones biológicas interespecíficas




Wikimedia foundation. 2010.