- Pterosauria
-
 Pterosaurios
PterosauriosRango temporal: Triásico - Cretácico 
Geosternbergia sternbergiClasificación científica Reino: Animalia Filo: Chordata Superclase: Tetrapoda Clase: Sauropsida Subclase: Diapsida (sin clasif.): Archosauria Orden: Pterosauria
Kaup, 1834Subórdenes Los pterosaurios (Pterosauria, gr. "lagartos alados") son un clado de saurópsidos arcosaurios voladores que existieron durante casi toda la Era Mesozoica (228 a 65 millones de años). Fueron los primeros vertebrados en conquistar el aire. Sus alas estaban formadas por una compleja membrana sostenida por el cuarto dedo de la mano, que estaba hipertrofiado. Su cuerpo estaba cubierto de pelo, preservado en fósiles de al menos tres especies distintas. Los fósiles de pterosaurios son muy comunes y se han hallado centenares de especímenes en África, Asia, Australia, Europa, Norteamérica y Sudamérica.
Los pterosaurios no son dinosaurios, aunque sí convivieron con ellos durante decenas de millones de años. El término "dinosaurio" está restringido a ciertos grupos de reptiles con una postura erguida única (el superorden Dinosauria, el cual incluye a las aves), y por lo tanto excluye a los pterosaurios, así como a varios grupos de reptiles marinos extintos como los ictiosaurios, plesiosaurios y mosasaurios.
Los pterosaurios son también frecuentemente denominados como "pterodáctilos", particularmente por periodistas.[1] Este uso es desaconsejado. "Pterodáctilo" se refiere específicamente a los miembros del género Pterodactylus,[2] y más ampliamente a los miembros del suborden Pterodactyloidea.[3] [4]
Contenido
Descubrimiento
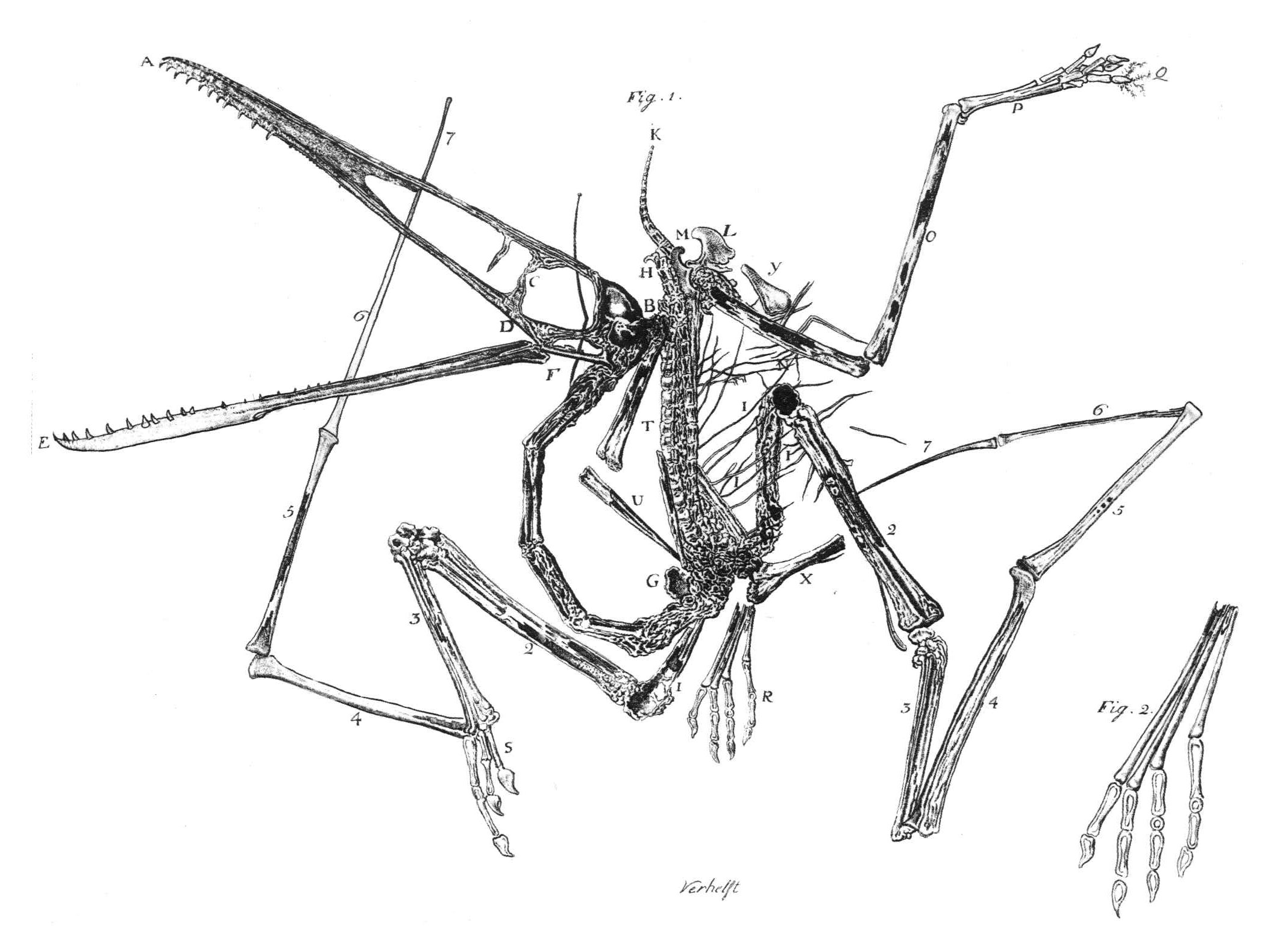
 Grabado del espécimen original de Pterodactylus antiquus por Egid Verhelst II, 1784
Grabado del espécimen original de Pterodactylus antiquus por Egid Verhelst II, 1784
El primer fósil de pterosaurio fue descrito por el naturalista italiano Cosimo Collini en 1784. Collini malinterpretó a su espécimen como una criatura marina que usaba sus largos miembros delanteros como aletas.[5] Unos pocos científicos continuaron apoyando la interpretación acuática incluso hasta 1830, cuando el zoólogo alemán Johann Georg Wagler sugirió que Pterodactylus usaba sus alas como aletas.[6] Georges Cuvier fue el primero en sugerir que los pterosaurios eran criaturas aéreas en 1801,[7] y acuñó el término "Ptero-dactyle" en 1809 para el espécimen recuperado en Alemania.[2] Sin embargo, debido a la estandarización de los nombres científicos, el nombre oficial para este género se volvió Pterodactylus, aunque el nombre "pterodáctilo" se volvió popular y se aplicó incorrectamente a todos los miembros de Pterosauria.[1] Los paleontólogos ahora evitan usar "pterodáctilo" y prefieren el término "pterosaurio". Ellos relegan el término "pterodáctilo" específicamente para los miebros del género Pterodactylus o más ampliamente para los miembros del suborden Pterodactyloidea.[3]
Desde que los primeros fósiles de pterosaurios fueron descubiertos en la Caliza de Solnhofen del Jurásico Superior en 1784, sólo en estos depósitos se han hallado veintinueve clases de pterosaurios. Un famoso hallazgo del Reino Unido fue un ejemplar de Dimorphodon hecho por Mary Anning, en Lyme Regis en 1828. El nombre Pterosauria fue acuñado por Johann Jakob Kaup en 1834, aunque el nombre Ornithosauria (o "aves lagarto", Bonaparte, 1838) fue usado a veces en la literatura científica temprana.[2]
 El cráneo preservado tridimensionalmente de Anhanguera santanae, de la formación Santana, Brasil
El cráneo preservado tridimensionalmente de Anhanguera santanae, de la formación Santana, BrasilMuchos fósiles de pterosaurio están muy mal preservados. Sus huesos eran huecos y cuando los sedimentos se apilaban sobre ellos, los huesos se aplanaban. Los fósiles mejor preservados provienen de la meseta de Araripe, en Brasil. Por alguna razón, cuando los huesos fueron depositados, los sedimentos encapsulaban a los huesos, en vez de aplastarlos. Esto creó fósiles de tres dimensiones que los paleontólogos pueden estudiar. El primer hallazgo en la meseta de Araripe fue realizado en 1974.
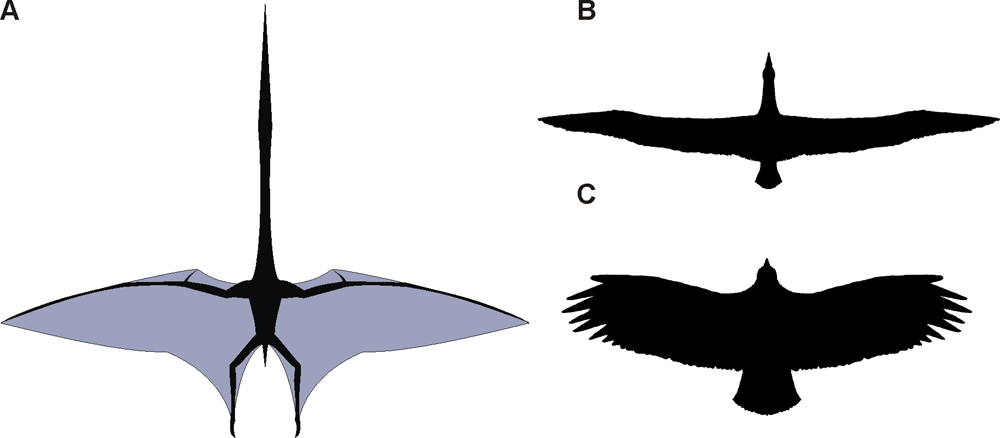
Muchos paleontólogos ahora creen que los pterosaurios estaban adaptados para un vuelo activo, no solo planear como se pensaba al principio. Los fósiles de pterosaurio se han hallado en cada continente. Al menos 60 géneros de pterosaurios se han hallado a la fecha, con tamaños que van desde el de un pájaro pequeño a envergaduras de más de 10 metros.
Primeros pterosaurios
Orígenes
 Reconstrucción de dos Scleromochlus en un árbol
Reconstrucción de dos Scleromochlus en un árbolDebido a que la anatomía de los pterosaurios estaba muy modificada para el vuelo, y a que no se ha encontrado un predecesor inmediato o "eslabón perdido", los orígenes de los pterosaurios no son muy bien comprendidos. Varias hipótesis se han barajado, incluyendo vínculos con ornitódiros como Scleromochlus, un ancestro entre los arcosauriformes basales como Euparkeria, o entre los prolacertiformes (los cuales incluyen formas planeadoras como Sharovipteryx).[8]
Dos investigadores, Chris Bennett (1996) y David Peters (2000), han encontrado que los pterosaurios eran prolacertiformes o estaban cercanamente relacionados a estos. Bennett sólo llegó a ese resultado después de remover características de los miembros posteriores en su análisis, en un intento de examinar la idea de que dichas características son el resultado de evolución convergente entre los pterosaurios y los dinosaurios. Sin embargo, análisis posteriores hechos por Dave Hone y Michael Benton (2007) no pudieron reproducir ese resultado. Hone y Benton encontraron que los pterosaurios eran parientes cercanos de los dinosaurios incluso sin los rasgos de los miembros posteriores. Ellos también criticaron los estudios previos de David Peters, planteando "serias dudas" sobre los métodos que él usó para situar a los pterosaurios entre los prolacertiformes. Hone y Benton concluyeron que aunque se necesitan más pterosauromorfos primitivos para clarificar sus relaciones, es mejor considerar a los pterosaurios arcosaurios, y específicamente ornitódiros, dada la evidencia actual. En el análisis de Hone y Benton, los pterosaurio son o considerados como el grupo hermano de Scleromochlus o caen entre éste y Lagosuchus en el árbol familiar de los ornitódiros.[9] Sterling Nesbitt (2011) halló un apoyo sólido para un clado compuesto de Scleromochlus y los pterosaurios.[10]
Pterosaurios primitivos
Los primeros pterosaurios pertenecen al suborden de los ranforrincoideos o pterosaurios de cola larga. Los fósiles más antiguos de pterosaurios pertenecen al menos al Triásico superior. El más antiguo se llama Eudimorphodon. Es de pequeño tamaño, cola con extremo en forma de rombo, y dientes puntiagudos en ambas mandíbulas.
Los pterosaurios, desarrollados entre el Triásico tardío y principios del Jurásico, eran similares al Eudimorphodon; de pequeño tamaño, cola larga, y dientes (como el Rhamphorynchus, que tenía dientes que cerraban en forma de trampa, y el Scaphognathus, que era uno de los pterosaurios más hábiles en el vuelo). Scaphongnathus significa «mandíbula de barca», y se cree que pudieron devorar a otros pterosaurios. Los ranforrincoideos llegaron a ser muy abundantes en el Jurásico inferior y medio de Inglaterra, Alemania, India y Arizona.
La mayor abundancia y mejor preservación se da en los depósitos del Jurásico superior de Solnhofen (Baviera), yacimiento que constituye uno de los tesoros del conocimiento paleontológico más preciosos del mundo, con sus más de 600 especies inmaculadamente preservadas, en una caliza tan fina que antaño era utilizada en la impresión de litografías.
Ciertas impresiones indican que Rhamphorhynchus poseía una bolsa en la garganta en la que almacenaba los peces, elementos básicos de la dieta de los pterosaurios de Solnhofen. Su sistema de pesca consistía seguramente en rápidos descensos en picado, gracias a los cuales capturaba a los peces de las aguas marinas superficiales con sus dientes salientes. Su cabeza era más larga que la de pterosaurios más evolucionados. Se cree que, en lugar de caminar, este género reptaba de manera parecida a los lagartos. Las amplias mandíbulas de otro género, Anurognathus sugieren que capturaba insectos con alas. Estos pterosaurios tenían alas muy poderosas, dientes en forma de gancho y un cuerpo menudo.
Pterodactylus, que ha sido hallado en África oriental, Inglaterra y Francia, es el primero de un nuevo grupo de pterosaurios, los pterodactiloideos, caracterizados por una cola muy corta, su mayor cabeza y un cuello más largo. Su nombre proviene del griego (pterodaktylos = dedo volador). Se cree que los pterodáctilos recorrían las costas en bandadas, al igual que hoy hacen las aves marinas. Poseían un hocico muy largo, probablemente útil para atrapar insectos en sus madrigueras, y quizás también comieran peces.
Otros géneros destacados fueron Batrachognathus, Germanodactylus, Dimorphodon, Sordes.
Pterosaurios del Cretácico
Los pterosaurios de cola larga (Rhamphorhynchoidea) no sobrevivieron en el Cretácico; sin embargo, los pterodactiloideos (pterosaurios de cola corta) se diversificaron y se extendieron por todos los continentes, originado formas sin dientes (como el Pteranodon), otros del tamaño de un avión de la Primera Guerra Mundial (como Quetzalcoatlus, la criatura voladora más grande conocida), numerosos con crestas (como Dsungaripterus y Pteranodon), y otros con otras características (como el Ctenochasma, que filtraba plancton por medio de sus finos dientes). Dsungaripterus tenía una envergadura alar de 3 metros, una inusual cresta ósea recorriendo su hocico y mandíbulas curvas, estrechas y largas que terminaban en punta. Vivió durante el Cretácico inferior.
Así pues, la tendencia era el aumento de tamaño, la pérdida progresiva de los dientes, y el ahuecamiento de los huesos de las partes no sometidas a los principales esfuerzos. Esto condujo a que, al final del Cretácico, hubiera dos familias:
- Pteranodóntidos, Vivieron en Norteamérica. Destaca Pteranodon, descubierto en la Formación Niobrara. Su envergadura alar alcanzaba de 5 a 8 metros. Su maciza cabeza acababa en un afilado pico desprovisto de dientes en la parte delantera, equilibrado por una estructura trasera que en ocasiones presentaba una forma similar a la del pico, y otras veces formaba una cresta ascendente, aerodinámica. Se cree que esta última pudo funcionar también de timón. Su cuerpo era minúsculo: las alas y la cabeza constituían una gran proporción de su total. No poseía cola y sus patas eran muy débiles, por lo que es improbable que pudiera andar.
- Azhdárquidos. Se extendieron por todo el mundo. Esta familia toma su nombre de Azhdarcho, un gran pterosaurio hallado en Uzbekistán en 1984 y bautizado como un dragón de la mitología uzbeka.
Aparentemente, los pterosaurios perdieron mucha de su diversidad antes de concluir la Era Mesozoica. Para esta epóca sólo se conocen especies grandes de pterosaurios. Las menores parecen haberse extinto, siendo llenados sus nichos por las aves.[11] Sólo se conoce una especie que existiese en el momento de la gran extinción masiva del Cretácico: Quetzalcoatlus northropi. Este animal fue descubierto en Texas, en 1975, y arrebató a Pteranodon su condición de «el mayor animal volador de todos los tiempos». Su nombre proviene de un dios azteca. Las estimaciones sobre la envergadura alar de aquel monstruo varían entre 11 y 15 metros, lo que cuadruplica la envergadura del mayor pájaro moderno, el cóndor. En lo concerniente a su tamaño y características de vuelo, tenía más en común con un ligero aeroplano que con cualquier criatura voladora actual. Sin embargo, el declive de lo spterosaurios (si realmente ocurrió) no parece directamente relacionado a la expansión de las aves.[12] Muchos pterosaurios parecen haberse especializado en un estilo de vida en hábitas marinos. Consecuentemente, los cambios ambientales a finales del Cretácico empezaron a afectar severamente la vida marina de la que dependían, muchos habrían desaparecido. Por otra parte, pterosaurios como los azdárquidos y los istiodactílidos eran animales de ambientes terrestres. Por supuesto, la extinción final de este grupo ocurrió en la extinción masiva del Cretácico Terciario, en la que también se produjo la extinción de los dinosaurios no avianos y muchos tipos de aves primitivas.
 Comparación de tamaño del Quetzalcoatlus
Comparación de tamaño del QuetzalcoatlusOtros géneros destacados fueron: Cearadactylus, Gnathosaurus, Ornithocheirus, Pterodaustro, Tropeognathus.
- Véase también:
Anatomía
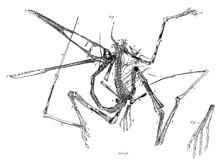
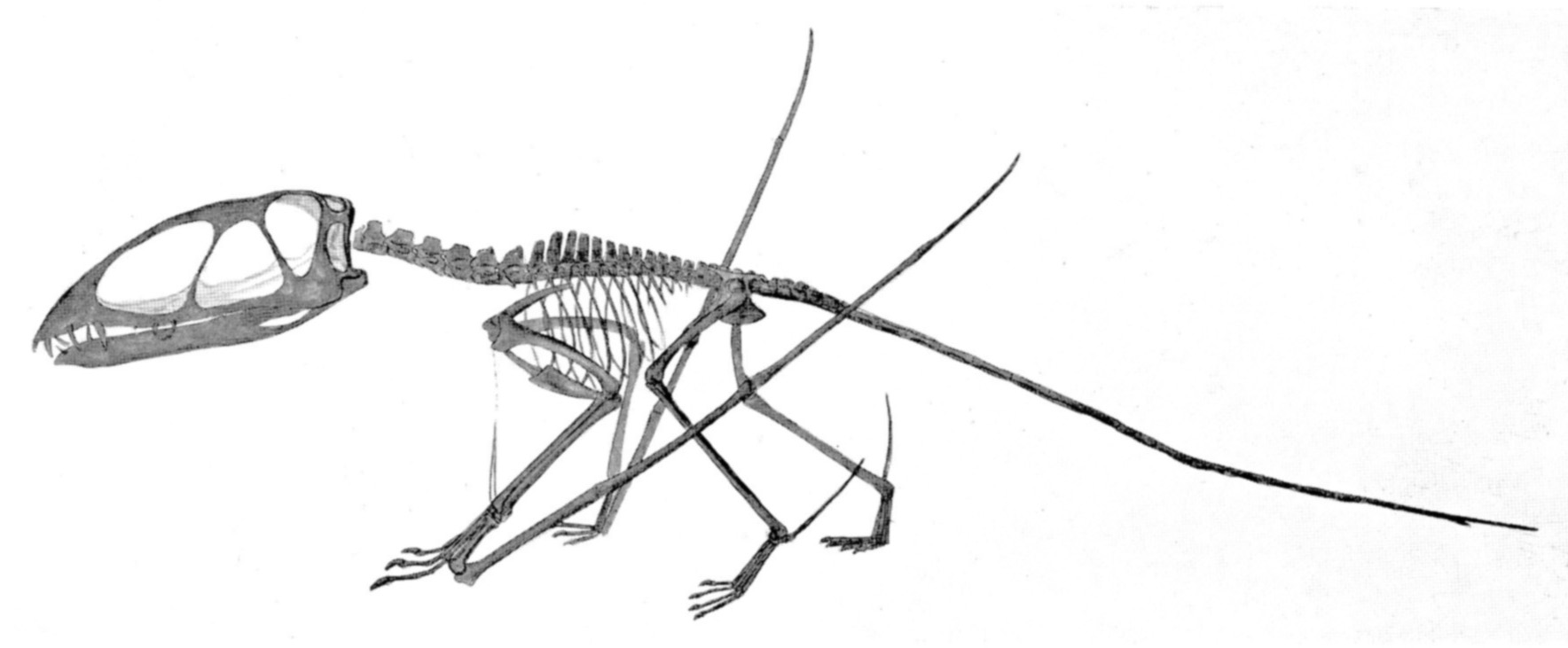
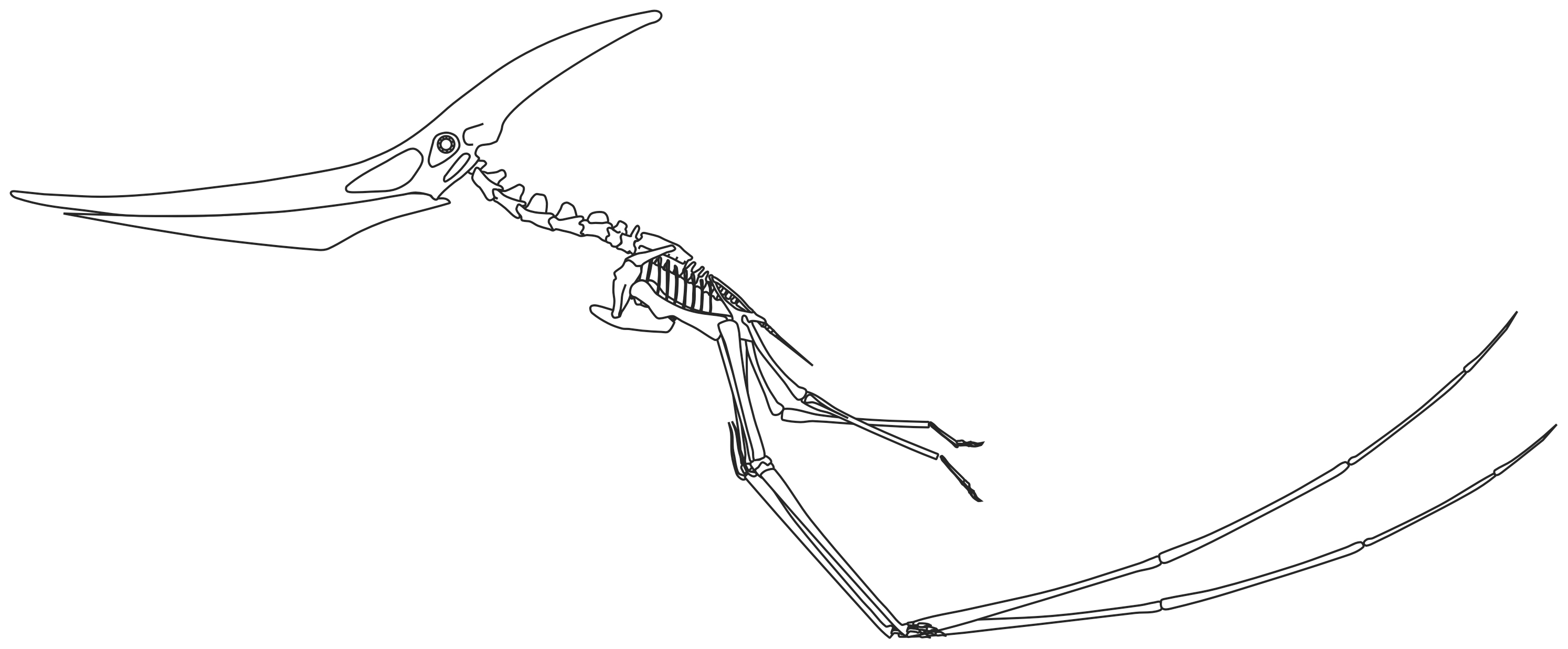
 Esqueleto de Pterodactylus.
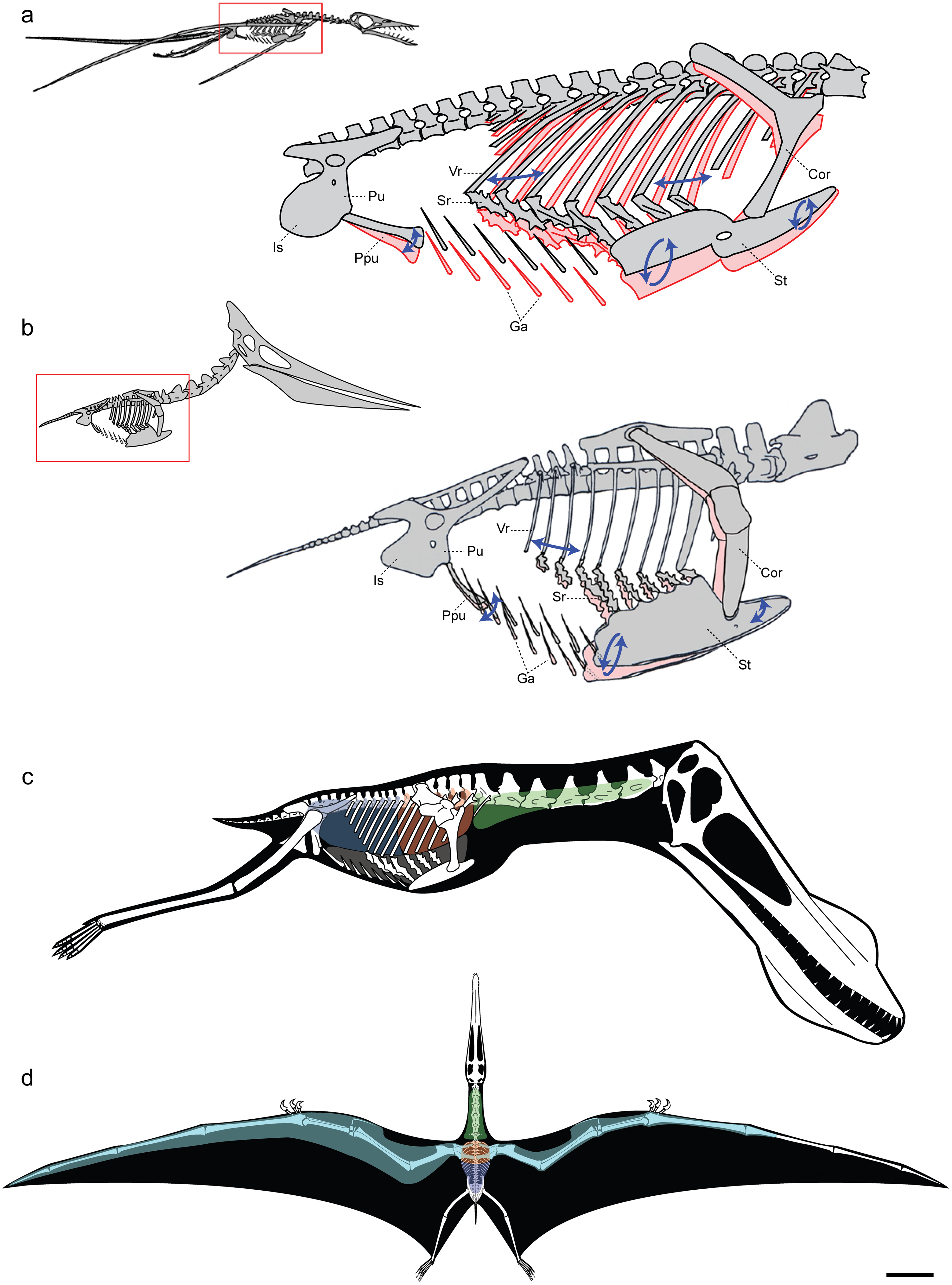
Esqueleto de Pterodactylus.El tamaño de los pterosaurios fue muy variable, y existieron representantes de tamaño enorme así como otros tan pequeños como un pájaro. Poseían un esternón que estaba desarrollado para el anclaje de los músculos implicados en el vuelo, y un gran cerebro que muestra características de especialización al vuelo.[13] En algunos pterosaurios, tardíos la columna vertebral sobre los hombros se fusionó en una estructura conocida como el notarium, que servía para darle rigidez al torso durante el vuelo, y daba un soporte estable para la escápula (omóplato).
Como en las aves actuales, los huesos eran huecos y poseían paredes muy delgadas, con el fin de ahorrar peso, lo que también se considera especialización al vuelo; en el espacio interior había un entramado de varillas óseas que aumentaba mucho la resistencia del hueso.
Alas
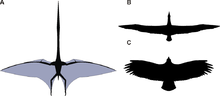 Superficie alar reconstruida de Quetzalcoatlus northropi comparada a la del albatros viajero Diomedea exulans y el cóndor andino Vultur gryphus (no a escala)
Superficie alar reconstruida de Quetzalcoatlus northropi comparada a la del albatros viajero Diomedea exulans y el cóndor andino Vultur gryphus (no a escala)Las alas de los pterosaurios estaban formadas por membranas de piel y de otros tejidos. La membrana principal se unía al extremadamente largo cuarto dedo de cada brazo y se extendía por los lados del cuerpo y las patas.
Aunque históricamente se ha pensado que eran una sencilla estructura de piel correosa, la investigación reciente ha mostrado que las membranas alares de los pterosaurios eran realmente estructuras muy complejas y dinámicas adaptadas a un estilo de vuelo activo. Primero, las alas exteriores (desde la punta hasta el codo) estaban fortalecidas por fibras poco espaciadas llamadas actinofibrillas.[14] Las actinofibrillas mismas consistían de tres capas distintas en el ala, formando un patrón entrecruzado cuando se superponía la una a la otra. La función real de las actinofibrillas es desconocida, así como el material preciso del que estaban hechas. Dependiendo de su composición exacta (queratina, músculo, estructuras elásticas, etc.), estas pudieron haber sido agentes de endurecimiento o de fortalecimiento en la parte externa del ala.[15] Las membranas del ala también contenían una delgada capa de músculo, tejido fibroso, y un complejo sistema circulatorio único de vasos sanguíneos en bucle.[16]
Como se evidencia de las cavidades vacías en los huesos del ala de las mayores especies y el tejido blando preservado en al menos un espécimen, algunos pterosaurios extendieron su sistema de sacos aéreos respiratorios dentro de la propia membrana alar.[17]
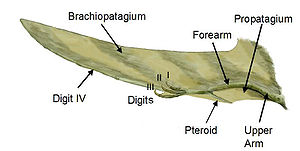
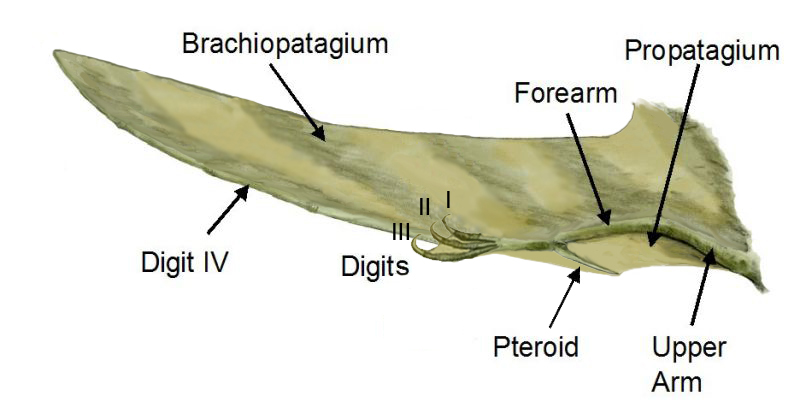
Partes del ala de un pterosaurio
 Sordes, como se representa aquí, evidencia la posibilidad de que los pterosaurios tuvieran un cruropatagio (una membrana conectando las patas, pero dejando la cola libre a diferencia del uropatagio de los quirópteros)
Sordes, como se representa aquí, evidencia la posibilidad de que los pterosaurios tuvieran un cruropatagio (una membrana conectando las patas, pero dejando la cola libre a diferencia del uropatagio de los quirópteros)La membrana del ala de los pterosaurios se divide en tres unidades básicas. La primera, llamada el propatagio ("primera membrana"), estaba en la parte delantera del ala y se sujetaba entre la muñeca y el hombro, creando el "borde de ataque" durante el vuelo. Esta membrana pudo haber incorporado a los primeros tres dedos de la mano, como se evidencia en algunos especímenes.[16] El braquiopatagio ("membrana del brazo") era el componente primario del ala, extendiéndose desde el alargado cuarto dedo de la mano a los miembros posteriores (aunque el punto exacto de sujeción es controversial y pudo haber variado entre especies). Finalmente, al menos algunos pterosaurios tenían una membrana que se extendía entre las piernas, posiblemente conectándose o incorporando la cola, llamada uropatagio; la extensión de esta membrana es incierta, y algunos estudios sobre Sordes parece sugerir que ésta simplemente se conectaba a las patas dejando de lado a la cola (representando así un cruropatagio). Es de consenso general que los pterosaurios no pterodactiloides tenían un amplio uro/cruropatagio, mientras los pterodactiloides sólo tenían membranas a lo largo de las piernas; Pteranodon en particular pudo haber desarrollado o re-evolucionado un uropatagio, dada la estructura de su cola.
 Anatomía del ala
Anatomía del alaUn hueso único de los pterosaurios, conocido como el pteroide, se conectaba a la muñeca y ayudaba a sostener la membrana delantera (el propatagio) entre la muñeca y el hombro. Evidencia de membranas entre los tres dedos del brazo sugiere que esta membrana delantera puede haberse extendido más allá de la conexión del pteroide al hombro que tradicionalmente se usa para las reconstrucciones en vida.[16] La posición del hueso pteroide en sí misma ha sido controversial. Algunos científicos, como David Unwin, han argumentado que el pteroide apuntaba hacia adelante, extendiendo la membrana delantera.[18] Sin embargo, esta idea fue refutada en un artículo de 2007 por Chris Bennett, quien mostró que el pteroide no podía articularse de esa manera, sino que más bien apuntaba hacia el cuerpo como tradicionalmente se ha pensado.[19]
Tres líneas de evidencia, morfológica, de desarrollo e histológica, indican que el pteroide es un hueso verdadero, en vez de ser cartílago osificado. El origen del pteroide no es claro: pudo haber sido un hueso carpo modificado, el primer metacarpo, o un neomorfo (hueso nuevo).[20]
La muñeca consiste en dos huesos carpos internos (proximales) y cuatro carpos externos (distales), excluyendo al hueso pteroide, el cual pudo haber sido un carpo distal modificado. Los carpos distales se fusionaron juntos en un "sincarpo" en especímenes maduros, mientras que tres de los carpos distales se fusionan para formar un sincarpo distal. El carpo distal restante, conocido como el carpo medio, pero también conocido como el distal lateral, o carpo preaxial, se articula en una faceta biconvexa verticalmente alargada sobre la superficie anterior del sincarpo distal. El carpo medio tenía una profunda fóvea cóncava que se abría anteriormente, ventralmente y un poco medialmente, dentro del cual se articulaba el pteroide.[21]
Ha existido una considerable discusión entre los paleontólogos acerca de sí la membrana alar principal (braquiopatagio) se unía a los miembros posteriores, y si era así, en dónde. Fósiles del ranforrincoide Sordes,[22] el anurognátido Jeholopterus,[23] y de pterodactiloides de la formación Santana parecen demostrar que la membrana alar se unía a los miembros posteriores, al menos en algunas especies.[24] Sin embargo, los modernos murciélagos y ardillas voladoras muestran una considerable variación en la extensión de sus membranas alares y es posible que, como en estos grupos, diferentes especies de pterosaurios tuvieran diferentes diseños de alas. Incluso, análisis de las proporciones de las extremidades de los pterosaurios muestran que tenían mucha variación, posiblemente reflejando una variedad de configuraciones alares.[25]
Muchos, si no todos los pterosaurios también tenían pies con membranas.[26]
Cráneo, dientes y crestas

Muchos cráneos de pterosaurios tenían mandíbulas alargadas, similares a picos. Algunas formas avanzadas eran desdentadas (como los pteranodóntidos y los azdárquidos), aunque muchos lucían una dotación completa de dientes similares a agujas.[8] En algunos casos, se ha preservado tejido del pico córneo, aunque en las formas dentadas, el pico era pequeño y se restringía a la punta de las mandíbulas, sin envolver a los dientes.[27]
A diferencia de muchos arcosaurios, los cuales tenían varias aberturas en el cráneo en el frente de sus ojos, en los pterosaurios pterodactiloides la abertura anteorbital y la abertura nasal se combinaron en una única gran abertura, llamada la fenestra nasoanteorbital. Esta probablemente evolucionó como una característica para ahorrar peso y así aligerar el cráneo para volar.[8]
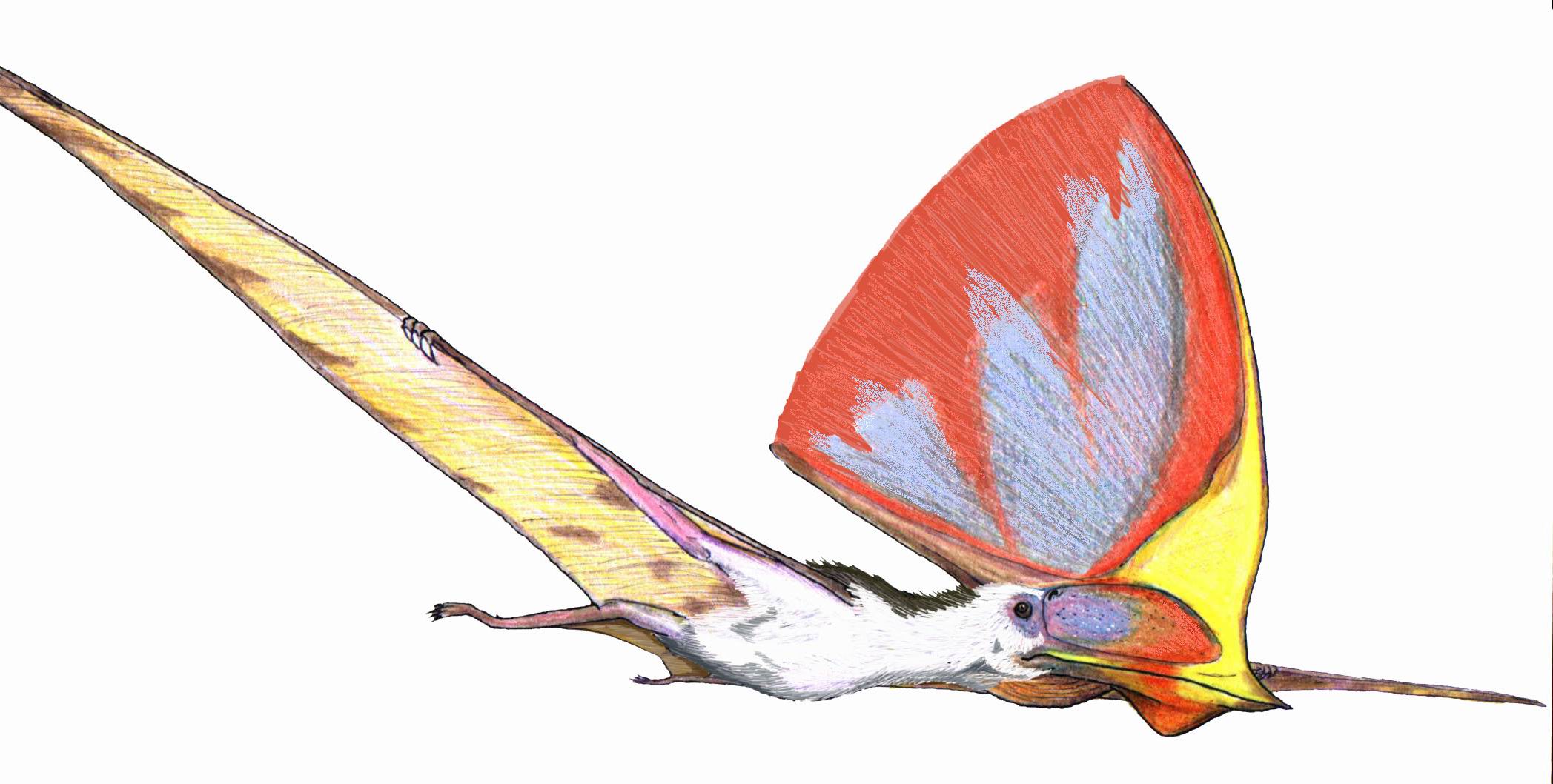
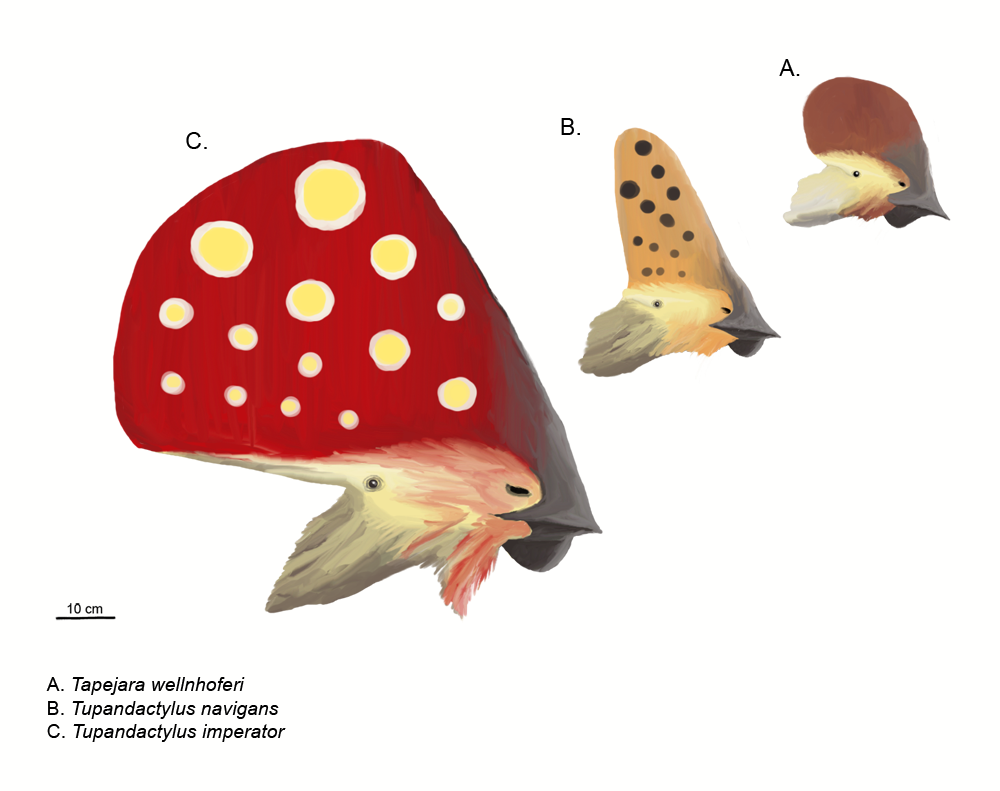
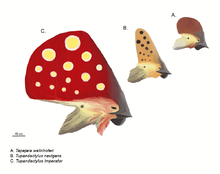 Reconstrucción de crestas: tres tapejáridos crestados. De arriba a abajo: Tapejara wellnhoferi, "Tapejara" navigans y Tupandactylus imperator (dibujados a escala).
Reconstrucción de crestas: tres tapejáridos crestados. De arriba a abajo: Tapejara wellnhoferi, "Tapejara" navigans y Tupandactylus imperator (dibujados a escala).Los pterosaurios son bien conocidos por sus frecuentemente elaboradas crestas. La primera y probablemente mejor conocida de éstas es la distintiva cresta que apuntaba hacia atrás de algunas especies de Pteranodon, aunque algunos pterosaurios, como los tapejáridos y Nyctosaurus lucían crestas increíblemente grandes que frecuentemente adicionaban extensiones de tejido queratinoso u otros tejidos blandos a la base ósea de la cresta.
Desde la década de 1990, nuevos descubrimientos y estudios más exhaustivos de los viejos especímenes han mostrado que las crestas estaban más extendidas entre los pterosaurios de los que inicialmente se había pensado, debido principalmente al hecho de que estaban frecuentemente hechas de extensiones de queratina, la cual no se fosiliza tan comúnmente como el hueso.[16] En los casos de pterosaurios como Pterorhynchus y Pterodactylus, la verdadera extensión de sus crestas se ha descubierto usando fotografías ultravioleta.[27] [28] El descubrimeinto de Pterorynchus y Austriadactylus, ambos "ranforrincoides" crestados, muestra que incluso los pterosaurios primitivos tenían estos adornos (previamente, se pensaba que estaban restringidos a los más avanzados pterodactiloides).[16]
Picnofibras
Al menos algunos pterosaurios estaban recubiertos con filamentos similares a pelo conocidos como picnofibras, similares pero no homólogos con el pelo de los mamíferos (es decir, no comparten una estructura común). Las picnofibras no eran verdadero pelo como el de los mamíferos, sino una estructura única que desarrolló una apariencia similar a través de la evolución convergente. Al menos en algunos casos las actinofibrillas (fibras estructurales internas) en la membrana del ala han sido confundidas con picnofibras o pelo verdadero, pero fósiles como los de Sordes pilosus (cuyo nombre se traduce como "demonio peludo") y Jeholopterus ninchengensis muestran inconfundibles impresiones de las picnofibras en la cabeza y cuerpo, no muy distinto a los murciélagos modernos, otro ejemplo de evolución convergente.[22] La presencia de las picnofibras (y las necesidades energéticas para el vuelo) implican que los pterosaurios eran endotérmicos (de sangre caliente).
El término "picnofibra", que significa "filamento denso", fue acuñado originalmente en un artículo sobre las impresiones de tejido blando de Jeholopterus por el paleontólogo Alexander W.A. Kellner y colegas en 2009.[15]
Sistema nervioso
Un estudio de la cavidad del cerebro en los pterosaurios usando rayos X reveló que los animales (específicamente Rhamphorhynchus muensteri y Anhanguera santanae) tenían un enorme flóculo. El flóculo es una región del cerebro que integra señales de las articulaciones, músculos, piel y órganos de balanceo.[13]
El flóculo de los pterosaurios ocupaba el 7.5% de la masa total del cerebro del animal, más que en cualquier otro vertebrado. Las aves tienen un flóculo inusualmente grande comparado con otros animales, pero este sólo ocupa entre el 1 al 2% de la masa cerebral total.[13]
El flóculo envía señales neurales que producen pequeños movimientos automáticos en los músculos de los ojos. Estos mantienen estable la imagen en la retina de un animal. Los pterosaurios pueden haber tenido un flóculo mayor debido a su gran tamaño alar, el cual debió implicar un mayor volumen de información sensorial para procesar.[13]
Fisiología
Los pterosaurios poseían un fino recubrimiento piloso, como puede apreciarse en algunos especímenes de Solnhofen y también, de forma aún más clara, en nuevos materiales procedentes de Kazajistán. El pelaje constituye un indicador muy fiable: la mayoría de los paleontólogos acepta que los pterosaurios eran animales de sangre caliente. Este rasgo posibilitaría la elevada tasa metabólica necesaria para desarrollar una activa existencia voladora. No hay duda de que los pterosaurios batían las alas al volar, pese a que algunas de las formas del Cretácico tardío eran tan grandes que debe suponerse que, en vez de batir las alas, planeaban.
Vuelo
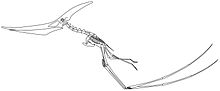 Reconstrucción esquelética de un Pteranodon longiceps impulsándose de forma cuadrúpeda
Reconstrucción esquelética de un Pteranodon longiceps impulsándose de forma cuadrúpedaLa mecánica del vuelo de los pterosaurios no es completamente entendida o modelada en la actualidad.[29] [30]
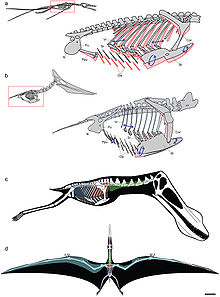 Diagramas mostrando los movimientos respiratorios (las dos imágenes superiores) y el sistema interno de sacos aéreos (las dos inferiores)
Diagramas mostrando los movimientos respiratorios (las dos imágenes superiores) y el sistema interno de sacos aéreos (las dos inferiores)Katsufumi Sato, un científico japonés, hizo cálculos usando a las aves modernas y concluyó que era imposible para un pterosaurio permanecer en el aire.[31] En el libro Posture, Locomotion, and Paleoecology of Pterosaurs (postura, locomoción y paleoecología de los pterosaurios) se teoriza que éstos eran capaces de volar debido a la densa atmósfera rica en oxígeno del período Cretácico Superior.[32] Sin embargo, se debe notar que tanto Katsufumi como los autores de Posture, Locomotion, and Paleoecology of Pterosaurs basaron su investigación en las ahora anticuadas ideas de que los pterosaurios eran como aves marinas, y el límite de tamaño no se aplica a los pterosaurios terrestres como los azdárquidos y los tapejáridos [3] Más aún, Darren Naish concluyó que las diferencias atmosféricas entre el presente y el Mesozoico no eran necesarias para que los pterosaurios alcanzaran grandes tamaños.[33]
Por su parte, Mark Witton y Mike Habib, de la Universidades de Portsmouth y de Johns Hopkins, respectivamente, afirman que los pterosaurios usaban un mecanismo de salto usando sus propias alas para alcanzar el vuelo, en vez de esperar corrientes de aire cálidas o usar riscos para impulsarse.[34] Una vez en el aire, los pterosaurios podían alcanzar velocidades de más de 120 km por hora y viajar por centenares de kilómetros.[34]
Sacos aéreos y respiración
Un estudio de 2009 mostró que los pterosaurios tenían un sistema de sacos aéreos pulmonares y una bomba respiratoria controlada de manera precisa, que apoya un modelo de ventilación pulmonar de flujo continuo en los pterosaurios, análogo al de las aves. La presencia de un sistema de sacos aéreos subcutáneos en la menos algunos pterodactiloides (particularmente los linajes de mayor tamaño, Azdarchoidea y Ornithocheiroidea) pudo haber adicionalmente reducido la densidad del animal vivo.[17]
Depredadores naturales
Se sabe que los pterosaurios eran devorados por terópodos. En la edición de julio 1 de 2004 de la revista Nature, el paleontólogo Eric Buffetaut comentó un fósil de tres vértebras de un pterosaurio del Cretácico Inferior con un diente roto de un espinosáurido incrustado en éstas. Se sabe que las vértebras no fueron comidas y expuestas a la digestión, ya que aún se encontraban articuladas.[35]
Locomoción terrestre
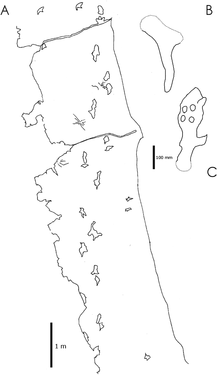 El fósil de las pisadas del probable azdárquido Haenamichnus uhangriensis.
El fósil de las pisadas del probable azdárquido Haenamichnus uhangriensis.Los acetábulos de las caderas de los pterosaurio están levemente orientados hacia arriba, y la cabeza del fémur (el hueso del muslo) está sólo moderadamente vuelta hacia adentro, sugiriendo que los pterosaurios tenían una postura semierecta. Es posible que hayan podido alzar el muslo de modo que adoptara una postura horizontal durante el vuelo como hacen los lagartos planeadores.
Ha existido un considerable debate sobre si los pterosaurios caminaban de manera cuadrúpeda o bípeda. En los años 1980, el paleontólogo Kevin Padian sugirió que los pterosaurios más pequeños con largos miembros posteriores como Dimorphodon pueden haber caminado o incluso podían correr bípedamente, además de volar, como los correcaminos.[36] Sin embargo, un gran número de icnitas de pterosaurios fueron más tarde halladas con distintivas marcas de pies con cuatro dedos y manos con tres dedos; estas son las inconfundibles huellas de pterosaurios caminando sobre cuatro patas.[37] [38]
A diferencia de muchos vertebrados, los cuales caminan sobre sus patas con los tobillos fuera del contacto con con el suelo (digitígrados), las huellas fósiles muestran que los pterosaurios ponían el pie entero en contacto con el suelo (eran plantígrados), de manera similar a la de los humanos y osos. Pisadas de azdárquidos muestran que al menos algunos pterosaurios caminaban con una postura erguida, en vez de extendida.[26]
 Pisadas fósiles muestran que los pterosaurios como Quetzalcoatlus northropi eran cuadrúpedos.
Pisadas fósiles muestran que los pterosaurios como Quetzalcoatlus northropi eran cuadrúpedos.Aunque tradicionalmente representados como desgarbados y torpes cuando estaban en el suelo, la anatomía de algunos pterosaurios (particularmente pterodactiloides) sugiere que eran caminantes y corredores competentes.[39] Los huesos de las extremidades anteriores de los azdárquidos y los ornithoqueíridos eran inusualmente largos comparados con los de otros pterosaurios, y en los azdárquidos, los huesos del brazo y la mano (metacarpales) estaban particularmente alargados. Más aún, los miembros delanteros de los azdárquidos en su totalidad estaban proporcionados de manera similar a los de los mamíferos ungulados corredores. Sus miembros posteriores, por otro lado, no estaban hechos para la velocidad, pero era muy largos comparados con muchos pterosaurios, y permitían una larga zancada. Aunque los pterosaurios azdárquidos probablemente no corrían, ellos pudieron haber sido relativamente rápidos y energéticamente eficientes.[26]
El tamaño relativo de las manos y pies de los pterosaurios (por comparación a los de animales modernos como las aves) pueden indicar qué tipo de vida llevaban en el suelo. Los azdárquidos tenían pies relativamente pequeños comparados a su tanmaño corporal y a la longitud de su pierna, de modo que la longitud del pie abarcaba sólo el 25%-30% de la longitud de la parte inferior de la pierna. Esto sugiere que los azdárquidos estaban bien adaptados a caminar en suelos secos y relativamente sólidos. Pteranodon tenía pies ligeramente más grandes (47% de la longitud de la tibia), mientras que los pterosaurios filtradores como los ctenocasmatoides tenían pies muy grandes (69% de la longitud de la tibia en Pterodactylus, 84% en Pterodaustro), adaptados a caminar en suelos suaves fangosos, similares a las modernas aves zancudas.[26]
Reproducción y crecimiento
 Fósil de una cría de pterodactiloide de la Caliza de Solnhofen
Fósil de una cría de pterodactiloide de la Caliza de SolnhofenMuy poco se sabe a cerca de la reproducción de este grupo, y los huevos de pterosaurios son muy raros. Con respecto a su reproducción y crianza, el primer huevo conocido fue hallado en las canteras de Liaoning, el mismo sitio en donde se han descubierto los famosos "dinosaurios emplumados". El huevo era aplanado y aplastado sin signos de rompimiento, por lo que poseían una cáscara blanda, como en los lagartos modernos.[40] Un más reciente descubrimiento de un fósil de un Darwinopterus, fosilizado en exacto instante en el cual estaba desovando un huevo, en el fondo de un lago en la provincia nororiental china de Liaoning en 2009 y denominado "Mrs. T", ha permitido despejar algunos aspectos y detalles de los mismos. Según los hallazgos del equipo de investigadores dirigidos por Junchang Lü, de la Academia China de Ciencias Geológicas en Pekín, los pterosaurios macho tenían una pelvis relativamente más pequeña y un hueso de cresta más largo en su cabeza, mientras que la pelvis de las hembras era más ancha y además éstas carecían de la mencionada cresta. El huevo, con alrededor de seis gramos de peso, era relativamente pequeño en comparación con el cuerpo de la hembra, de manera similar a lo que ocurre en la actualidad con los reptiles.[41] Un ave actual de un tamaño similar al pterosaurio hallado en China pone huevos que tienen entre dos y tres veces el tamaño del encontrado, porque los huevos deben contener todas las sustancias para el desarrollo del pichón. La envoltura blanda, similar a un pergamino, de los huevos de los pterosaurios hace suponer que las hembras enterraban a sus huevos, al igual que lo hacen muchos reptiles actuales y, luego, prácticamente no brindaban más atención, porque la envoltura permitía la entrada de agua del entorno. Es posible que poco antes de que naciera la cría, el huevo de "Mrs T" duplicara su peso. Los científicos suponen que "Mrs. T" estaba a punto de poner el huevo, cuando un acontecimiento desconocido causó la ruptura de su extremidad delantera izquierda.[42] El enterramiento de los huevos puede haber sido beneficioso en la evolución temprana de los pterosaurios, al permitirle más adaptaciones de aligeramiento, pero este método de reproducción también pudo haber limitado la variedad de ambientes en que podían vivir, y pudo haberlos puesto en desventaja cuando se enfrentaron a la competencia ecológica de las aves.[43]
Las membranas del ala preservadas en embriones de pterosaurio están bien desarrollados, lo que sugiere que los pterosaurios estaban listo para volar poco después de nacer.[44] Fósiles de pterosaurios de sólo unos pocos días o semanas de edad (llamados flaplings -"aleteantes"- en la literatura científica en inglés) se han descubierto en varias familias de pterosaurios, incluyendo pterodactílidos, ranforrínquidos, ctenocasmátidos y azdárquidos.[8] Todos los huesos preservados muestran un relativamente alto grado de endurecimiento (osificación) para su edad, y las proporciones de sus alas son similares a las de los adultos. De hecho, muchas crías de pterosaurios se han considerado adultos y colocado en especies separadas en el pasado. Adicionalmente, las crías son usualmente halladas en los mismos sedimentos que los adultos y juveniles de la misma especie, como ocurre con Pterodactylus y Rhamphorhynchus de la Caliza de Solnhofen de Alemania, y Pterodaustro de Brasil. Todos fueron hallados en un ambiente de agua profunda, lejos de la costa.[45]
No se sabe si los pterosaurios practicaban alguna forma de cuidado paternal, pero su habilidad para volar tan pronto salían del cascarón y las numerosas crías halladas en ambientes lejos de sus nidos y junto a los adultos ha llevado a muchos investigadores, incluyendo a Christopher Bennett y David Unwin, a concluir que los jóvenes sólo dependían de sus padres por un breve período de tiempo, mientras que las alas crecían lo suficiente para volar, y dejaban el nido para defenderse por sí mismos pocos días después de la eclosión.[8] Alternativamente, ellos pudieron haber usado la yema almacenada para alimentarse durante sus primeros días de vida, como en muchos reptiles modernos, más que depender de los padres para su comida.[45]
Las tasas de crecimiento de los pterosaurios una vez eclosionaban variaban según los distintos grupos. En los más primitivos, las formas de cola larga ("ranforrincoides") como Rhamphorhynchus, la tasa promedio de crecimiento en su primer año de vida estaba entre 130% a 173%, levemente más rápida que la de un aligator. El crecimeinto en estas especies era más lento después de la madurez sexual, y pudo haberle tomado más de tres años a un Rhamphorhynchus alcanzar su tamaño máximo. En contraste, los pterodactiloides más avanzados y grandes como Pteranodon crecía a su tamaño adulto dentro de su primer año de vida. Adicionalmente, los pterodactiloides tenían un crecimiento determinado, lo que significa que los animales alcanzaban un tamaño máximo predeterminado en su adultez y entonces paraban de crecer.[45]
Patrones de actividad diaria
Las compariciones entre los anillos escleróticos de los pterosaurio con aves y reptiles modernos han sido usadas para inferir los patrones de actividad diaria de los pterosaurios. Los géneros Pterodactylus, Scaphognathus y Tupuxuara se han considerado como diurnos, Ctenochasma, Pterodaustro y Rhamphorhynchus han sido inferidos como nocturnos, y Tapejara ha sido inferido como un catemeral, es decir que estaba activo todo el día por cortos intervalos. Como resultado, los posibles piscívoros Ctenochasma y Rhamphorhynchus pueden haber tenido patrones de actividad similares a muchas aves marinas nocturnas, y el filtrador Pterodaustro puede haber sido como las modernas aves anseriformes que se alimentan de noche. Las diferencias entre los patrones de actividad de los pterosaurios de la Caliza de Solnhofen como Ctenochasma, Rhamphorhynchus, Scaphognathus y Pterodactylus puede indicar una división de nichos ecológicos entre estos géneros.[46]
Clasificación y filogenia
 Diagrama que compara los huesos de pterosaurios, murciélagos y aves.
Diagrama que compara los huesos de pterosaurios, murciélagos y aves.Puesto que la anatomía de los pterosaurios estaba enormemente modificada para el vuelo, y los predecesores inmediatos no se han descrito convenientemente, el linaje de los pterosaurios resulta confuso. La clasificación de los pterosaurios siempre ha sido difícil, porque existen muchos huecos en el registro fósil. Así pues, la clasificación del orden Pterosauria, con sus grupos más importantes, es como sigue:
- Orden Pterosauria†
- Suborden Rhamphorhynchoidea*
- Familia Dimorphodontidae
- Familia Anurognathidae
- Familia Campylognathoididae
- Familia Rhamphorhynchidae
- Suborden Pterodactyloidea
- Superfamilia Ornithocheiroidea
- Familia Istiodactylidae
- Familia Ornithocheiridae
- Familia Pteranodontidae
- Familia Nyctosauridae
- Superfamilia Ctenochasmatoidea
- Familia Gallodactylidae
- Familia Pterodactylidae
- Familia Ctenochasmatidae
- Superfamilia Dsungaripteroidea
- Familia Germanodactylidae
- Familia Dsungaripteridae
- Superfamilia Azhdarchoidea
- Familia Lonchodectidae
- Familia Tapejaridae
- Familia Azhdarchidae
- Superfamilia Ornithocheiroidea
- Suborden Rhamphorhynchoidea*
El cladograma completo es el siguiente:
Pterosauria Macronychoptera Dimorphodontidae
Caelidracones Anurognathidae
Lonchognatha Campylognathoididae
Breviquartossa Pterodactyloidea Ornithocheiroidea Euornithocheira Pteranodontidae
Lophocrania Ctenochasmatoidea Euctenochasmia Ctenochasmatidae
Dsungaripteroidea Dsungaripteridae
Azhdarchoidea En la cultura popular
 Modelos de Quetzalcoatlus en South Bank, creados por Mark Witton para el Aniversario no. 350 de la Real Sociedad
Modelos de Quetzalcoatlus en South Bank, creados por Mark Witton para el Aniversario no. 350 de la Real SociedadLos pterosaurios han sido matera prima de la cultura popular tanto como sus primos los dinosaurios, aunque ellos no aparecen usualmente de manera prominente en películas, literatura u otras artes. Adicionalmente, mientras que la representación de los dinosaurios en los medios de comunicación ha cambiado radicalmente en respuesta a los avances en la paleontología, una imagen mayormente anticuada de los pterosaurios ha persistido desde mediados del siglo XX.[47]
El número y la diversidad de los pterosaurios en la conciencia popular tampoco es tan alta como lo ha sido históricamente para los dinosaurios. Mientras que el término genérico "pterodáctilo" es frecuentemente usado para describir a estas criaturas, los animales frecuentemente representados representan a Pteranodon o bien a Rhamphorhynchus, o a un híbrido ficticio de los dos.[47] Muchos juguetes para niños y caricaturas presentan "pterodáctilos" con crestas como la de Pteranodon y dientes y largas colas como en Rhamphorhynchus, una combinación que nunca existió en la naturaleza. Sin embargo, al menos un tipo de pterosaurio tuvo una cresta como la de Pteranodon y dientes, como es el caso de Ludodactylus, un nombre que significa "dedo de juguete" por su parecido a los viejos e inexactos juguetes infantiles.[48] Asimismo, algunas representaciones de pterosaurios incorrectamente los identifican como "aves", cuando en la vida real eran un grupo de reptiles voladores, y las aves de hecho son descendientes de los dinosaurios terópodos.
Los pterosaurios aparecieron en la ficción por primera vez en la novela de Arthur Conan Doyle de 1912 El mundo perdido, y en su correspondiente adaptación filmíca de 1925. Ellos también han aparecido en un cierto número de películas y programas de televisión desde entonces, incluyendo la película de 1933 King Kong, y Un millón de años a.C. en 1966. En este última, el animador Ray Harryhausen tuvo que añadir alas erróneas similares a las de los murciélagos para su modelo de stop motion con el fin de evitar que las membranas del ala se desmoronaran, aunque este error particular fue común en el arte incluso antes de que el filme fuera realizado. Los pterosaurios estuvieron mayormente ausentes de cualquier aparición en películas notables hasta 2001, en Jurassic Park III. Sin embargo, el paleontólogo Dave Hone ha notado que incluso después de 40 años de investigación, los pterosaurios en dicha película no han sido significativamente actualizados para reflejar el conocimiento actual. Entre los errores que él notó como persistentes desde 1960 a la década de 2000 estaban los dientes incluso en las especies desdentadas (los pterosaurios de Jurassic Park III se supone que eran Pteranodon, cuyo nombre significa "alado y desdentado"), comportamiento de anidación que se sabe ya era erróneo en 2001, y alas como cuero, en vez de las membranas tensadas de fibra muscular que realmente estaban presentes y que eran necesarias para que los pterosaurios volaran.[47]
Una mutación ficcionalizada de un pterosaurio fue presentada en la película japonesa de 1956 Rodan. La película fue estrenada por Toho, el mismo estudio responsable de las cintas de Godzilla. El personaje más tarde apareció en algunas de las películas de Godzilla entre 1964 a 2004.
Referencias
- ↑ a b Darren Naish. «Pterosaurs: Myths and Misconceptions». Pterosaur.net. Consultado el 18 de junio de 2011.
- ↑ a b Caroline Arnold & Laurie A. Caple (2004). «Pterodactyl». Pterosaurs: rulers of the skies in the dinosaur age. Houghton Mifflin Harcourt. p. 23. ISBN 9780618313549. http://books.google.com/books?id=ltaev1auSTAC&lpg=PP1&pg=PA23#v=onepage&q&f=false.
- ↑ a b David E. Alexander & Steven Vogel (2004). Nature's Flyers: Birds, Insects, and the Biomechanics of Flight. JHU Press. p. 191. ISBN 9780801880599. http://books.google.com/books?id=zj395mz_GYkC&lpg=PP1&pg=PA191#v=onepage&q&f=false.
- ↑ Ron Redfern (2001). Origins: the evolution of continents, oceans, and life. University of Oklahoma Press. p. 335. ISBN 9780806133591. http://books.google.com/books?id=PqyMMs--IM4C&lpg=PP1&pg=PA335#v=onepage&q&f=false.
- ↑ Collini, C A. (1784). "Sur quelques Zoolithes du Cabinet d’Histoire naturelle de S. A. S. E. Palatine & de Bavière, à Mannheim." Acta Theodoro-Palatinae Mannheim 5 Pars Physica, pp. 58–103 (1 plate).
- ↑ Wagler, J. (1830). Natürliches System der Amphibien Munich, 1830: 1-354.
- ↑ Cuvier, G. (1801). [Reptile volant]. In: Extrait d’un ouvrage sur les espèces de quadrupèdes dont on a trouvé les ossemens dans l’intérieur de la terre. Journal de Physique, de Chimie et d’Histoire Naturelle, 52: 253–267.
- ↑ a b c d e Unwin, David M. (2006). The Pterosaurs: From Deep Time. New York: Pi Press. p. 246. ISBN 0-13-146308-X.
- ↑ Hone D.W.E., Benton M.J. (2007). «An evaluation of the phylogenetic relationships of the pterosaurs to the archosauromorph reptiles». Journal of Systematic Palaeontology 5 (4): pp. 465–469. doi:.
- ↑ Nesbitt, S.J. (2011). «The early evolution of archosaurs: relationships and the origin of major clades». Bulletin of the American Museum of Natural History 352: pp. 1–292. http://digitallibrary.amnh.org/dspace/bitstream/2246/6112/1/B352.pdf.
- ↑ Slack KE, Jones CM, Ando T, et al (June 2006). «Early penguin fossils, plus mitochondrial genomes, calibrate avian evolution». Mol. Biol. Evol. 23 (6): pp. 1144–55. doi:. PMID 16533822. http://mbe.oxfordjournals.org/cgi/reprint/23/6/1144.
- ↑ Copyrighted excerpt from Richard Butler, Paul Barrett, Steven Nowbath & Paul Upchurch [1]; might require new link
- ↑ a b c d Witmer LM, Chatterjee S, Franzosa J, Rowe T (October 2003). «Neuroanatomy of flying reptiles and implications for flight, posture and behaviour». Nature 425 (6961): pp. 950–3. doi:. PMID 14586467.
- ↑ Bennett SC (2000). «Pterosaur flight: the role of actinofibrils in wing function». Historical Biology 14 (4): pp. 255–84. doi:. http://www.informaworld.com/smpp/content~content=a907600799~db=all.
- ↑ a b Kellner, A.W.A., Wang, X., Tischlinger, H., Campos, D., Hone, D.W.E. and Meng, X. (2009). "The soft tissue of Jeholopterus (Pterosauria, Anurognathidae, Batrachognathinae) and the structure of the pterosaur wing membrane." Proceedings of the Royal Society B, published online before print August 5, 2009, doi:10.1098/rspb.2009.0846
- ↑ a b c d e Naish D, Martill DM (2003). «Pterosaurs — a successful invasion of prehistoric skies». Biologist 50 (5): pp. 213–6.
- ↑ a b Claessens LP, O'Connor PM, Unwin DM (2009). «Respiratory evolution facilitated the origin of pterosaur flight and aerial gigantism». PLoS ONE 4 (2): pp. e4497. doi:. PMID 19223979. PMC 2637988. http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0004497.
- ↑ Wilkinson MT, Unwin DM, Ellington CP (January 2006). «High lift function of the pteroid bone and forewing of pterosaurs». Proc. Biol. Sci. 273 (1582): pp. 119–26. doi:. PMID 16519243.
- ↑ Bennett SC (2007). «Articulation and Function of the Pteroid Bone of Pterosaurs». Journal of Vertebrate Paleontology 27 (4): pp. 881–91. doi:. ISSN 0272-4634. http://www.bioone.org/perlserv/?request=get-abstract&doi=10.1671%2F0272-4634(2007)27%5B881%3AAAFOTP%5D2.0.CO%3B2.
- ↑ Unwin, D. M.; Frey, E., Martill, D. M., Clarke, J. B., and Riess, J. (1996). «On the nature of the pteroid in pterosaurs». Proceedings of the Royal Society: Biological Sciences 263 (1366): pp. 45–52. doi:.
- ↑ Wilkinson M.T., Unwin D.M., Ellington C.P. (2006). «High lift function of the pteroid bone and forewing of pterosaurs». Proceedings of the Royal Society B 273 (1582): pp. 119–126. doi:. PMID 16519243.
- ↑ a b Unwin DM, Bakhurina NN (1994). «Sordes pilosus and the nature of the pterosaur flight apparatus». Nature 371 (6492): pp. 62–4. doi:.
- ↑ Wang X, Zhou Z, Zhang F, Xu X (2002). «A nearly completely articulated rhamphorhynchoid pterosaur with exceptionally well-preserved wing membranes and "hairs" from Inner Mongolia, northeast China». Chinese Science Bulletin 47 (3): p. 3. doi:. ISSN 1001-6538.
- ↑ Frey et al., (2003) New specimens of Pterosauria (Reptilia) with soft parts with implications for pterosaurian anatomy and locomotion Geological Society London Special Publications
- ↑ Dyke GJ, Nudds RL, Rayner JM (July 2006). «Limb disparity and wing shape in pterosaurs». J. Evol. Biol. 19 (4): pp. 1339–42. doi:. PMID 16780534.
- ↑ a b c d Witton MP, Naish D (2008). «A reappraisal of azhdarchid pterosaur functional morphology and paleoecology». PLoS ONE 3 (5): pp. e2271. doi:. PMID 18509539. PMC 2386974. http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0002271.
- ↑ a b Frey E, Martill DM (1998). «Soft tissue preservation in a specimen of Pterodactylus kochi (Wagner) from the Upper Jurassic of Germany». Neues Jahrbuch fu ̈r Geologie und Pala ̈ontologie, Abhandlungen 210: pp. 421–41.
- ↑ Czerkas, S.A., and Ji, Q. (2002). A new rhamphorhynchoid with a headcrest and complex integumentary structures. In: Czerkas, S.J. (Ed.). Feathered Dinosaurs and the Origin of Flight. The Dinosaur Museum:Blanding, Utah, 15-41. ISBN 1-932075-01-1.
- ↑ Alleyne, R., Pterodactyls Were Too Heavy To Fly, Scientist Claims, Telegraph, Oct 2008
- ↑ Powell, D., Were Pterosaurs Too Big To Fly?, Oct 2008
- ↑ Alleyne, Richard. «Pterodactyls were too heavy to fly, scientist claims», The Daily Telegraph, 01-10-2008. Consultado el 22-05-2010.
- ↑ Templin, R. J.; Chatterjee, Sankar (2004). Posture, locomotion, and paleoecology of pterosaurs. Boulder, Colo: Geological Society of America. p. 60. ISBN 0-8137-2376-0. http://books.google.com/?id=idta6AVV-tIC&pg=PA60&dq=Pterosaur+oxygen.
- ↑ Naish, Darren (18 de febrero de 2009). «Pterosaurs breathed in bird-like fashion and had inflatable air sacs in their wings». ScienceBlogs. Consultado el 20 de octubre de 2010.
- ↑ a b «Did giant pterosaurs vault aloft like vampire bats?», New Scientist.
- ↑ Buffetaut E, Martill D, Escuillié F (July 2004). «Pterosaurs as part of a spinosaur diet». Nature 430 (6995): p. 33. doi:. PMID 15229562.
- ↑ Padian K (1983). «A Functional Analysis of Flying and Walking in Pterosaurs». Paleobiology 9 (3): pp. 218–39.
- ↑ Padian K (2003). «Pterosaur Stance and Gait and the Interpretation of Trackways». Ichnos 10 (2–4): pp. 115–126. doi:.
- ↑ Hwang K, Huh M, Lockley MG, Unwin DM, Wright JL (2002). «New pterosaur tracks (Pteraichnidae) from the Late Cretaceous Uhangri Formation, southwestern Korea». Geological Magazine 139 (4): pp. 421–35. doi:.
- ↑ Unwin DM (1997). «Pterosaur tracks and the terrestrial ability of pterosaurs». Lethaia 29 (4): pp. 373–86. doi:.
- ↑ Ji Q, Ji SA, Cheng YN, et al (December 2004). «Palaeontology: pterosaur egg with a leathery shell». Nature 432 (7017): p. 572. doi:. PMID 15577900.
- ↑ Lü J., Unwin D.M., Deeming D.C., Jin X., Liu Y., Ji Q. (2011). «An egg-adult association, gender, and reproduction in pterosaurs». Science 331 (6015): pp. 321–324. doi:. PMID 21252343.
- ↑ http://www.lagaceta.com.ar/nota/418551/Informaci%C3%B3n_General/reptiles-voladores-enterraban-sus-huevos-como-lo-hacen-cocodrilos.html
- ↑ Grellet-Tinner G, Wroe S, Thompson MB, Ji Q (2007). «A note on pterosaur nesting behavior». Historical Biology 19 (4): pp. 273–7. doi:.
- ↑ Wang X, Zhou Z (June 2004). «Palaeontology: pterosaur embryo from the Early Cretaceous». Nature 429 (6992): p. 621. doi:. PMID 15190343.
- ↑ a b c Bennett S. C. (1995). «A statistical study of Rhamphorhynchus from the Solnhofen Limestone of Germany: Year-classes of a single large species». Journal of Paleontology 69: pp. 569–580.
- ↑ Schmitz, L.; Motani, R. (2011). «Nocturnality in Dinosaurs Inferred from Scleral Ring and Orbit Morphology». Science in press. doi:. PMID 21493820.
- ↑ a b c Hone, D. (2010). "Pterosaurs In Popular Culture." Pterosaur.net, http://www.pterosaur.net/popular_culture.php Accessed online 27-August-2010.
- ↑ Frey, E., Martill, D., and Buchy, M. (2003). A new crested ornithocheirid from the Lower Cretaceous of northeastern Brazil and the unusual death of an unusual pterosaur. In: Buffetaut, E., and Mazin, J.-M. (eds.). Evolution and Palaeobiology of Pterosaurs. Geological Society Special Publication 217:56-63. ISBN 1-86239-143-2.
- Bennett, S.C. (2000): Pterosaur flight: the role of actinofibrils in wing function. Historical Biology, 14: 255-284
- Unwin, D. M. (2003): On the phylogeny and evolutionary history of pterosaurs. En: Buffetaut, E. y Mazin, J.-M. (Eds.): Evolution and Palaeobiology of Pterosaurs. Londres, Geological Society, London, Special Publications, 217: 139-190
- Wellnhofer, P. (1991): The Illustrated Encyclopedia of Pterosaurs. Nueva York, Crescent Books. 192 págs. ISBN 0-517-03701-7 [Enciclopedia Ilustrada de los Pterosaurios. Madrid, Susaeta Ediciones, S.A. 192 págs. (1994) ISBN 84-305-7602-9]
- Witmer, W.M.; Chatterjee, S.; Franzosa, J. y Rowe, T. (2003): Neuroanatomy of flying reptiles and implications for flight, posture and behaviour. Nature, 425: 950-953
Enlaces externos
 Wikiespecies tiene un artículo sobre Pterosauria. Wikispecies
Wikiespecies tiene un artículo sobre Pterosauria. Wikispecies Wikimedia Commons alberga contenido multimedia sobre Pterosauria. Commons
Wikimedia Commons alberga contenido multimedia sobre Pterosauria. Commons- Base de datos de pterosaurios (inglés)
- Ilustraciones de pterosaurios (alemán)
Notas
Categorías:- Fósiles
- Pterosauria









Wikimedia foundation. 2010.