- Arthropoda
-
Arthropoda
 ?Arthropoda
?Arthropoda
Rango fósil: Cámbrico - Presente
Clasificación científica Dominio: Eukarya Reino: Animalia Subreino: Eumetazoa Superfilo: Ecdysozoa Filo: Arthropoda
Latreille, 1829Grandes grupos - Trilobitomorpha
- Chelicerata
- Crustacea
- Myriapoda
- Hexapoda
Los artrópodos (Arthropoda, del griego αρθρον, arthron, "articulación" y πούς, pous, "pie") constituyen el filo más numeroso y diverso del reino animal (Animalia). Incluye, entre otros, insectos, arácnidos, crustáceos y miriápodos.
Hay más de un millón de especies descritas, insectos en su mayoría,[1] que representan al menos el 80% de todas las especies animales conocidas. Varios grupos de artrópodos están perfectamente adaptados a la vida en el aire, igual que los vertebrados amniotas, a diferencia de todos los demás filos de animales, que o son acuáticos o requieren ambientes húmedos. Su anatomía, su fisiología y su comportamiento revelan un diseño simple pero admirablemente eficaz.
Contenido
Características
Los artrópodos constituyen una de las grandes divisiones del reino animal, subdividida en diversas clases, algunas de las cuales cuentan con gran número de géneros y especies. Se los denomina de esta manera por estar provistos de patas articuladas. En realidad no son solo las patas, sino todo el cuerpo el que está formado por varios segmentos unidos entre sí por medio de articulaciones.
A pesar de su variedad y su disparidad, los artrópodos poseen en común características morfológicas y fisiológicas fundamentales:
- Presencia de apéndices articulados que muestran una plasticidad evolutiva enorme y que han dado lugar a las estructuras más diversas (patas, antenas), branquias, pulmones, mandíbulas, quelíceros, etc.
- Presencia de un esqueleto externo o exoesqueleto quitinoso que mudan periódicamente. Dado que diversos filos pseudocelomados también mudan la cutícula, algunos autores relacionan los artrópodos con los nematodos y grupos afines, en un clado llamado ecdisozoos.[2]
- Cuerpo constituido por segmentos repetitivos, fenómeno conocido como metamería, con lo que el cuerpo aparece construido por módulos repetidos a lo largo del eje antero-posterior.La segmentación va acompañada de regionalización o tagmatización, con división del cuerpo en dos o tres regiones en la mayoría de los casos. Por este carácter se les ha relacionado tradicionalmente con los anélidos que también son animales metamerizados;[3] [4] [5] pero los defensores del clado ecdisozoos argumentan que es un caso de convergencia evolutiva (véase Articulata y Ecdysozoa, y en este mismo artículo el apartado Filogenia).
Exoesqueleto
Véase también: Cutícula (artrópodos)El exoesqueleto de los artrópodos es una cubierta continua llamada cutícula, que se extiende incluso por los dos extremos del tubo digestivo y por las vías o cavidades respiratorias, y que está situada por encima de la epidermis (llamada en éstos por ese motivo hipodermis), que es quien la secreta.
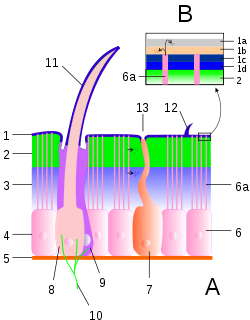 A: Cutícula y epidermis; B: Detalle de la epicutícula. 1: Epicutícula; 1a: Cemento; 1b: Ceras; 1c: Epicutícula externa; 1d: Epicutícula interna. 2: Exocutícula; 3: Endocutícula; 2+3: Procutícula; 4: Epitelio; 5: Lámina basal; 6: Célula epitelial; 6a: Canal poroso; 7: Célula glandular; 8: Célula tricógena; 9: Célula tormógena; 10: Terminación nerviosa; 11: Pelo sensorial; 12: Pelo; 13: Poro glandular.
A: Cutícula y epidermis; B: Detalle de la epicutícula. 1: Epicutícula; 1a: Cemento; 1b: Ceras; 1c: Epicutícula externa; 1d: Epicutícula interna. 2: Exocutícula; 3: Endocutícula; 2+3: Procutícula; 4: Epitelio; 5: Lámina basal; 6: Célula epitelial; 6a: Canal poroso; 7: Célula glandular; 8: Célula tricógena; 9: Célula tormógena; 10: Terminación nerviosa; 11: Pelo sensorial; 12: Pelo; 13: Poro glandular.
La composición del exoesqueleto es glucopeptídica (con una parte glucídica y una parte peptídica). El componente principal y más característico pertenece al primero de estos dos tipos, y es la quitina, un polisacárido derivado del aminoazúcar N-acetil-2-D-glucosamina que se encuentra también, por ejemplo, en la pared celular de los hongos. En muchos casos la consistencia del exoesqueleto gana por el añadido de sustancias minerales, como en el caso de los cangrejos y otros crustáceos decápodos cuya cutícula aparece calcificada, por depósito de carbonato cálcico.
El espesor y dureza de la cutícula no es igual en toda su extensión. Por el contrario, aparece formando zonas endurecidas llamadas escleritos, separadas o unidas entre sí por zonas más delgadas y flexibles. Los escleritos reciben denominaciones complejas que varían en cada grupo, pero se llaman de manera general terguitos los de ubicación dorsal, esternitos los de ubicación ventral y pleuritos los laterales. Pueden existir además crestas del exoesqueleto desarrolladas hacia adentro llamadas apodemas, sobre las que se insertan los músculos. La cutícula suele además estar atravesada por poros.
El exoesqueleto está estructurado en capas:
- Epicutícula. Muy delgada, estratificada a su vez y con propiedades hidrófobas que le confieren una función impermeabilizante. Está compuesta de proteínas y sustancias lipídicas tales como ceras. Donde es más delgada se facilita el intercambio de sustancias, por ejemplo la transpiración.
- Procutícula. Es la parte principal y más gruesa de la cutícula. Está formada a su vez por dos capas:
- Exocutícula. Esta parte es la de espesor más desigual y la más rígida. Su dureza deriva de la presencia de compuestos fenólicos que enlazan a los otros polímeros. Abunda en los escleritos y es más delgada o está ausente en las zonas de articulación.
- Endocutícula. Gruesa pero a la vez flexible y de espesor más uniforme que la exocutícula.
La cutícula aparece muy frecuentemente cubierta de quetas (pelos) de diversa función, incluida la sensorial táctil.
La coloración de los artrópodos suele depender de la cutícula. En la procutícula se depositan pigmentos coloreados o cristales de guanina. La epicutícula puede presentar estriaciones finas que producen colores físicos (no químicos), como la apariencia metálica o irisada de muchos insectos.
Ecdisis
 Ecdisis de una Cigarra
Ecdisis de una CigarraEl esqueleto externo tiene una desventaja y es que, para poder crecer, el animal debe desprenderse de él. Lo hace en un proceso, controlado hormonalmente, de ecdisis o muda. La hipodermis secreta enzimas que ablandan y digieren en parte la capa más inferior de la cutícula (la endocutícula), provocando que el resto se desprenda. Inmediatamente comienza la secreción de una cutícula nueva, primero la exocutícula y luego, debajo de ella, la procutícula. Hasta que no se endurece esta nueva cubierta el animal está relativamente indefenso, con menos posibilidad de escapar o resistirse. Todo el proceso de la muda está controlado hormonalmente; la ecdisona u "hormona de la muda" es la hormona responsable de que estos cambios se produzcan. Se llaman estadios o instares a las sucesivas fases de la existencia del animal entre muda y muda. Este rasgo lo comparten los artrópodos con algunos otros filos, como los nemátodos que también tienen una cutícula y mudan; hay una teoría que los clasifica ahora juntos en un subreino Ecdysozoa.
Apéndices
Para los apéndices el exoesqueleto aporta tubos huecos articulados, en cuyo interior se sitúan los tejidos vivos y específicamente los músculos estriados que, adheridos a ambos lados de las articulaciones, les proporcionan versatilidad y rapidez de movimientos. Se llama artejos (voz que deriva del latín artículo, "articulado") a las piezas articuladas que forman los apéndices.
Existen dos tipos básicos de apéndices, los unirrámeos, formados por un solo eje, propios de los artrópodos terrestres (arácnidos, miriápodos e insectos), y los birrámeos formados por dos ejes y propios de los artrópodos acuáticos (trilobites y crustáceos). No hay acuerdo sobre cuál fue el apéndice ancestral.
En el curso de la evolución ha existido la tendencia a restringir los apéndices a determinadas regiones del cuerpo y a especializarlos para funciones distintas. Los apéndices de la cabeza están adaptados para la percepción sensorial, la defensa y para manipular los alimentos; los del tórax sirven para andar y nadar; los abdominales cumplen funciones respiratorias y reproductoras, como retener los huevos o aferrarse a la pareja durante la cópula. Otros se han modificado de tal modo que cuesta reconocerlos como tales (hileras de las arañas, peines de los escorpiones).
Aparato digestivo
Véase también: Aparato digestivo (insectos)El aparato digestivo de los artrópodos se divide en tres regiones bien diferenciadas, el estomodeo, el mesodeo y el proctodeo. Estomodeo y proctodeo son las regiones situadas en el extremo anterior y del posterior, respectivamente; son de origen ectodérmico por lo que están recubiertas de cutícula que se renueva cada vez que el animal muda. La parte media del tubo digestivo, el mesodeo, deriva del endodermo (segunda hoja blastodérmica) y es la que produce las secreciones digestivas y donde se realiza la mayor parte de la absorción de nutrientes; frecuentemente presenta derivaciones o ciegos laterales que amplían su superficie.
Respiración
Muchos artrópodos son demasiado pequeños como para tener o necesitar órganos respiratorios, dada su gran relación superficie/volumen.
Los artrópodos acuáticos, lo que incluye a los primeros miembros del grupo, suelen presentar branquias, apéndices internamente más vascularizados que los otros órganos. Se encuentran en los crustáceos, como especializaciones de la rama dorsal de los apéndices torácicos, y de la misma manera en los xifosuros o en los euriptéridos o los primeros escorpiones fósiles. También se encuentran branquias secundarias (derivadas de las tráqueas) en las larvas acuáticas de algunos insectos, como las efímeras.
Como es general en los animales, los miembros del grupo de vida aérea respiran por órganos internalizados, que en los artrópodos pueden ser de dos tipos:
- Tráqueas. Los insectos, algunos órdenes de arácnidos, los miriápodos o las cochinillas de la humedad (crustáceos del orden isópodos adaptados a la vida terrestre) presentan una red de conductos (sistema traqueal) que comunican con el exterior por orificios llamados espiráculos, frecuentemente dotados de aberturas provistas de válvulas que regulan su diámetro. La cutícula se extiende por ellos en una versión muy delgada y permeable, que en todo caso se desprende cuando llega la muda. En algunos casos se observa una ventilación activa, con movimientos cíclicos de inspiración y espiración.
- Pulmones en libro. Presentan una estructura interna muy plegada (pulmones en libro), lo que multiplica la superficie por la que se realiza el intercambio de gases, y se abren al exterior por aberturas propias e independientes. Se encuentran pulmones en libro en varios órdenes de arácnidos, entre los que destacan las arañas y los escorpiones.
Circulación
El aparato circulatorio de los artrópodos es abierto. Es decir, no existe un circuito cerrado de vasos por el que circule un líquido diferenciado, lo que propiamente se podría llamar sangre. Lo que existe es un motor de bombeo que es un vaso especializado de posición dorsal, al que se denomina corazón dorsal, que mueve el líquido corporal interno, la hemolinfa, que recibe de vasos posteriores abiertos e impulsa hacia adelante por vasos igualmente abiertos. La red de vasos está siempre escasamente desarrollada, salvo en las branquias de los artrópodos acuáticos. No hay células sanguíneas especializadas en el transporte de oxígeno, aunque, como en todos los animales existen amebocitos (células ameboideas) con funciones de inmunidad celular y hemostasis (coagulación y cicatrización). Sí puede haber pigmentos respiratorios, pero disueltos en la hemolinfa.
Excreción
Véase también: Tubos de MalpighiLos crustáceos presentan para la excreción glándulas antenales y maxilares, en la base de esos apéndices. Los arácnidos suelen disponer de glándulas coxales, que desembocan en la base de las patas locomotoras. En insectos y en miriápodos aparecen órganos tubulares característicos, llamados tubos o conductos de Malpighi, que desembocan entre el intestino medio y el intestino posterior (proctodeo); sus productos se suman a la composición de las heces.
Los artrópodos terrestres suelen ser uricotélicos, es decir, que para la excreción nitrogenada no producen amoníaco o urea, sino ácido úrico o, a veces, guanina.
En los artrópodos es frecuente la excreción por acumulación, como alternativa o complemento de la excreción por secreción. En este caso se acumulan los productos de excreción en nefrocitos, células pericárdicas o directamente en la cutícula. La acumulación suele ser de uratos o guanina, bases nitrogenadas muy poco solubles que forman depósitos sólidos. En este último caso las mudas sirven para la función añadida de librarse de esas excretas.
Sistema nervioso
Véase también: Sistema nervioso (insectos)Como corresponde a los protóstomos, el sistema nervioso se desarrolla en el lado ventral del cuerpo y, como corresponde a animales metaméricos, su organización es segmentaria. En cada segmento aparece un par de ganglios, de posición más o menos ventrolateral, con los dos ganglios de un par soldados o unidos por una comisura transversal y los de pares consecutivos unidos por nervios conectivos.
Sistema nervioso central
En los artrópodos es un órgano de tipo anelidiano, por tanto, tiene una estructura primariamente en forma de escalera de cuerda, o sea, dos cordones nerviosos longitudinales que recorren la parte ventral del cuerpo, con un par de ganglios por metámero unidos tranversalmente por comisuras; no obstante, se producen procesos de concentración de ganglios debidos a la formación de tagmas.
Cerebro o sincerebro
Normalmente está formado por tres pares de ganglios que se asocian, correspondientes al procéfalon. Se pueden diferenciar tres regiones:
- Protocerebro. Es el resutado de la fusión entre el ganglio impar del arquicerebro, dependiente del acron, y del par de ganglios del prosocerebro; es preoral. El protocerebro posee las estructuras relacionadas con los ojo compuestos, ocelos y el sistema endocrino:
- Lóbulos prefrontales. Es una amplia región de la zona media del protocerebro donde se diferencian grupos de neuronas que constituyen la pars intercerrebralis; están relacionados con los ocelos y con el complejo endocrino. También se diferencia el cuerpo central y los cuerpos pedunculados o fungiformes. Estos dos centros son de asociación, están muy desarrollados en los insectos sociales. Van a regir en ellos la conducta de la colonia y el gregarismo de la misma.
- Lóbulos ópticos. Inervan los ojos compuestos, y en ellos radica la visión. Están muy desarrollados en animales con ojos complejos como hexápodos o crustáceos. Se diferencian tres centros:
- Lámina externa
- Médula externa
- Médula interna
- Éstos están relacionados entre sí por quiasmas.
- Deutocerebro. Resultado de la fusión de un par de ganglios; preoral. Del deutocerebro parten nervios que inervan el primer par de antenas (anténulas) de crustáceos y la antenas de hexápodos y miriápodos. En esos nervios hay que diferenciar dos ramas, la motora y la sensitiva. Además existen grupos de neuronas en los que residen centros de asociación con función olfativa y gustativa. Esos centros también se presentan en el tritocerebro. Los quelicerados carecen de deutocerebro; unos autores opinan que está atrofiado, mientras que otros creen que nunca lo han tenido.
- Tritocerebro. Resultado de la fusión de un par de ganglios; en origen es postoral. El tritocerebro inerva el segundo par de antenas de crustáceos, y en hexápodos y miriápodos, el segmento intercalar o premandibular, carente de apéndices. En los quelicerados inerva los quelíceros. De él parten nervios que lo relacionan con el sistema nervioso simpático o vegetativo (en el caso de los hexápodos, con el denominado ganglio frontal). Además del tritocerbro parte un conectivo periesofágico que se une al primer par de ganglios de la cadena nerviosa ganglionar ventral, y una comisura subesofágica que une los dos ganglios tritocerebrales entre sí.
En el protocerebro y deutocerebro, no se diferencian comisuras ni conectivos. El tritocerebro está formado por un par de ganglios que se unen a los anteriores en las cabezas denominadas tritocefálicas, perdiéndose los conectivos, mientras que en las cabezas deutocefálicas, se mantiene independiente, conservando los conectivos con el deutocerebro. Esto ocurre en algunos crustáceos como branquiópodos o cefalocáridos. En todos los casos, se diferencia la comisura, que es subesofágica.
Dentro de la cápsula cefálica, el cerebro tiene posición vertical; el protocerebro y el deutocerebro se sitúan hacia arriba, y el tritocerebro es inferior y se dirige hacia atrás.
Cadena nerviosa ganglionar ventral.
Está formada por un par de ganglios por metámero que en principio presentan conectivos y comisuras. En grupos primitivos, los ganglios de cada par de segmentos se presentan disociados, y la estructura recuerda a una escalera de cuerda. Los grados de concentración y de acortamiento se deben a la supresión de las comisuras y los conectivos respectivamente.
Destaca el ganglio subesofágico; en hexápodos es resultado de la fusión de tres pares de ganglios ventrales correspondientes a los metámeros IV, V y VI e inerva las piezas bucales (las mandíbulas y los dos pares de maxilas) y por ello se llama gnatocerebro; en los decápodos, son seis los ganglios que se asocian (pues se incluyen los tres ganglios de los maxilípedos.
Sistema nervioso simpático o vegetativo
Neuronas sensitivas y motoras que forman ganglios y que se sitúan sobre las paredes del estomodeo. Este sistema está relacionado con el sistema nervioso central y con el sistema endocrino. En el sistema nervioso simpático se diferencian dos partes.
- Sistema simpático estomatogástrico. Siempre existe. Es de forma diversa, está formado por ganglios impares, unidos entre sí por nervios recurrentes. Tiene como función la regulación de los procesos de deglución y los movimientos peristálticos del tubo digestivo. Regula también los latidos cardíacos.
- Sistema simpático terminal o caudal. Puede o no existir. Es también impar, y está ligado a los últimos ganglios de la cadena nerviosa ganglionar ventral. Tiene como función la de inervar el proctodeo, actuar en procesos reproductores, de puesta de huevos y transferencia de esperma, y también regula los latidos de los estigmas de los últimos segmentos del abdomen.
Sentidos
Véase también: Sensilia Ojos compuestos de una libélula. Ampliando la imagen se distinguen los omatidios.
Ojos compuestos de una libélula. Ampliando la imagen se distinguen los omatidios.La mayoría de los artrópodos están dotados de ojos, de los que existen varios modelos distintos.
- Los ojos simples son cavidades esferoidales con una sencilla retina y cubiertos frontalmente por una córnea transparente, Su rendimiento óptico es muy limitado, con la excepción de los grandes ojos de algunas familias de arañas, como los saltícidos.
- Los ojos compuestos están constituidos por múltiples elementos equivalentes llamados omatidios que se disponen radialmente, de manera que cada uno apunta en una dirección diferente y entre todos cubren un ángulo de visión más o menos amplio. Cada omatidio contiene varias células sensibles, retinianas, detrás de elementos ópticos transparentes, cumpliendo la función que la córnea y el cristalino desempeñan en los ojos de los vertebrados. También hay células que envuelven el omatidio sellándolo frente a la luz. No todos los grupos presentan ojos compuestos, que están ausentes, por ejemplo, en los arácnidos.
La visión de muchos artrópodos presenta ventajas que suelen faltar en vertebrados, como la habilidad para ver en un espectro extendido que incluye el ultravioleta próximo, o para distinguir la dirección de polarización de la luz. La visión del color está casi siempre presente y puede ser muy rica; el crustáceo Squilla mantis, la galera, presenta trece pigmentos distintos con diferente sensibilidad al color, lo que contrasta con el pobre sistema tricromático (de tres pigmentos) de la mayoría de los primates, incluida nuestra especie.
Distribuidos por todo el cuerpo, pueden encontrarse sensilias, que son receptores sensibles a los estímulos químicos, como los del gusto o el olfato, y receptores táctiles, asociado a antenas y palpos y también a setas táctiles, pelos que está asociados a una célula sensible. Algunos insectos disponen de un sentido del oído, lo que es revelado por la existencia de señales auditivas de comunicación intraespecífica, como por ejemplo en los grillos. Muchos son sensibles a las vibraciones del suelo, por las que detectan la presencia de presas o depredadores; otros, como las moscas, poseen tricobotrios capaces de percibir mínimos cambios de presión ambiental, por ello, los matamoscas están perforados.
Los artrópodos suelen estar dotados de sensores de posición, sencillos pero eficaces, que les ayudan a mantener la posición y el equilibrio, como los órganos cordotonales que un díptero tiene en los halterios.
Reproducción
Siempre se reproducen sexualmente. Las hembras, tras ser fecundadas por los machos, ponen huevos. El desarrollo, a partir del huevo, puede ser directo o indirecto.
- En el desarrollo directo nace un individuo similar al adulto, aunque, como es lógico, de menor tamaño.
- En el desarrollo indirecto nace una larva que implica una serie de cambios profundos denominados metamorfosis.
Se dan frecuentes casos de partenogénesis, sobre todo en crustáceos e insectos.
Filogenia
Durante muchas décadas, las relaciones filogenéticas de los Celomados se basaron en la concepción de los Articulados de Cuvier,[6] un clado formado por Anélidos y Artrópodos. Numerosos análisis morfológicos modernos basados en principios cladistas han corroborado la existencia del clado Articulados, por ejemplo, Brusca & Brusca,[3] Nielsen[4] o Nielsen et al.,[5] entre otros.
No obstante diversos análisis cladísticos, como el de datos combinados de Zrzavý et al (1998)[2] están llegando a la conclusión de que anélidos y artrópodos no están directamente ralcionados. La presencia de metamerización en anélidos y artrópodos debería considerarse, pues, como una convergencia evolutiva. Por el contrario, estos estudios proponen el clado Ecdysozoa en el que los artrópodos muestran estrechas relaciones filogenéticas con grupos pseudocelomados, como nematodos, nematomorfos, priapúlidos y quinorrincos, por la presencia compartida de una cutícula quitinosa y un proceso de muda (ecdisis) de la misma.
La filogenia de los Artrópodos ha sido muy controvertida, con una enfrentada polémica entre los partidarios del monofiletismo y los del polifiletismo. Snodgrass[7] y Cisne[8] han defendido el monofiletismo, aunque el primero contempla los Artrópodos divididos en Aracnados + Mandibulados, y el segundo los interpreta divididos en Esquizorrámeos y Atelocerados. Tiegs & Manton[9] defendieron el difiletismo, con los Artrópodos divididos en Esquizorrámeos + Unirrámeos y los Onicóforos como grupo hermano de Miriápodos + Hexápodos. Posteriormente, Manton[10] y Anderson[11] sostuvieron el polifiletismo del grupo (ver Uniramia).
Con la aparición de los primeros estudios basados en datos moleculares y análisis combinados de datos morfológicos y moleculares, parece que la antigua polémica sobre monofilia y polifilia ha quedado superada, ya que todos ellos corroboran que los Artrópodos son un grupo monofilético en el que incluyen también los Tardígrados (el clado se ha dado en llamar Panartrópodos); la mayoría también proponen la existencia del clado Mandibulados. No obstante, han surgido nuevas controversias, sobre todo alrededor de dos hipótesis alternativas mutuamente excluyentes que están siendo debatidas en numerosos artículos sobre filogenia y evolución de Artrópodos: Atelocerados (Miriápodos + Hexápodos) (Wheeler)[12] (cladograma A) versus Pancrustáceos (Crustáceos + Hexápodos) (Giribert & Ribera)[13] (cladograma B):
A
Panarthropoda Mandibulata Atelocerata ___ B
Panarthropoda Mandibulata Pancrustacea Clasificación
La clasificación de los artrópodos varía según se tenga en cuenta la hipótesis de los pancrustáceos o la de los atelocerados:
Hipótesis Pancrustacea
- Subfilo Trilobitomorpha†
- Subfilo Chelicerata
- Arachnida - arañas, escorpiones, ácaros, etc.
- Merostomata (Eurypterida y Xiphosura)
- Pycnogonida
- Subfilo Myriapoda
- Subfilo Pancrustacea
Hipótesis Atelocerata
- Subfilo Trilobitomorpha†
- Subfilo Chelicerata
- Arachnida - arañas, escorpiones, ácaros, etc.
- Merostomata (Eurypterida y Xiphosura)
- Pycnogonida
- Subfilo Crustacea[14]
- Subfilo Atelocerata
Referencias
- ↑ Chapman, A. D., 2005. Numbers of Living Species in Australia and the World
- ↑ a b Zrzavý, J., Mihulka, S., Kepka, P., Bezdék, A. & Tietz, D., 1998. Phylogeny of Metazoa based on morphological and 18S ribosomal DNA evidence. Cladistics, 14(3): 249-285.
- ↑ a b Brusca, R. C. & Brusca, G. J., 1990. Invertebrates. Sinauer, Sunderland.
- ↑ a b Nielsen, C., 1985. Animal phylogeny in the light of the trochaea theory. Biol. J. Linnean Soc., 25: 243-299.
- ↑ a b Nielsen, C., Scharff, N. & Eibye-Jacobsen, D., 1996. Cladistic analyses of the animal kingdom. Biol. J. Linnean Soc., 57: 385-410.
- ↑ Cuvier, G., 1812. Sur un nouveau repprochement à establir entre les classes qui composant le Règne Animal. Ann. Mus. Hist., 19: 73-84.
- ↑ Snodgrass, R. E., 1938. The evolution of Annelida, Onychophora, and Arthropoda. Smithson. Misc. Coll., 97: 1-159.
- ↑ Cisne, J. L., 1974. Trilobites and the origin of Arthropods. Science, 186: 13-18.
- ↑ Tiegs, O. W. & Manton, S. M., 1958. The evolution of the Arthropoda. Biol. Rev., 33: 255-337.
- ↑ Manton, S., 1964. Mandibular mechanisms and the evolution of Arthropods. Philos. Trans. R. Soc. Lond. (Ser. B, Biol. Sci.), 247: 1-183.
- ↑ Anderson, D. T., 1973. Embryology and Phylogeny in Annelids and Arthropods. Pergamon Press, Oxford.
- ↑ Wheeler, W. C., 1998. Sampling, grounplans, total evidence and the systematics of arthropods. En: R. A. Fortey & R. H. Thomas (eds.): Arthropod Relationships: 87-96. Chapman & Hall, London
- ↑ Giribert, G. & Ribera, C., 1998. The position of arthropods in the animal kingdom: A search for a reliable outgroup for internal arthropod phylogeny. Mol. Phylog. Evol., 9: 481-488.
- ↑ a b J. W. Martin & G. E. Davis (2001). An Updated Classification of the Recent Crustacea. Natural History Museum of Los Angeles County, pp. 132 pp.
Enlaces externos
 Wikiespecies tiene un artículo sobre Arthropoda.
Wikiespecies tiene un artículo sobre Arthropoda.
 Wikimedia Commons alberga contenido multimedia sobre Arthropoda.
Wikimedia Commons alberga contenido multimedia sobre Arthropoda. Wikiversidad alberga proyectos de aprendizaje sobre Arthropoda.Wikiversidad
Wikiversidad alberga proyectos de aprendizaje sobre Arthropoda.Wikiversidad- Biología artrópodos
- Web de la Sociedad Entomológica Aragonesa
- Entomoblog; magnífico blog sobre artrópodos
Categorías: Arthropoda | Filos del reino animal | Entomología



Wikimedia foundation. 2010.