- Scilloideae
-
 Hyacinthaceae
Hyacinthaceae
Hyacinthus orientalisClasificación científica Reino: Plantae División: Angiospermae Clase: Monocotyledoneae Orden: Asparagales Familia: Asparagaceae Subfamilia: Scilloideae Burnett Géneros ver texto
Las escilóideas (nombre científico Scilloideae) forman una subfamilia de plantas herbáceas, bulbosas y perennes que se incluyen dentro de las asparagáceas. Sus casi 10 000 especies se distribuyen predominantemente en climas mediterráneos, especialmente en Sudáfrica y el Mediterráneo hasta Asia Central y Birmania (algunas en Sudamérica). Poseen seis tépalos y seis estambres con ovario súpero, por eso se las solía ubicar dentro de las liliáceas. Se caracterizan por sus hojas bastante carnosas y mucilaginosas que se disponen en una roseta basal y por poseer compuestos venenosos (bufodienolidos y cardenolidos), por lo que sus especies no son comestibles. Algunos géneros son muy populares en jardinería, como Hyacinthus (el conocido jacinto), importante como planta cultivada y como flor cortada, y otros como Muscari, Scilla, Galtonia, Hyacinthoides y Ornithogalum.[1] [2] [3] [4]
Contenido
Descripción
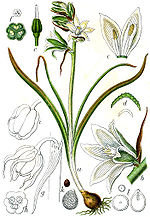 Morfología de Ornithogalum.
Morfología de Ornithogalum.
 Inflorescencia en Hyacinthoides.
Inflorescencia en Hyacinthoides. Inflorescencias de Urginea.
Inflorescencias de Urginea. Hojas y bulbos de Ledebouria.
Hojas y bulbos de Ledebouria.Las hiacintáceas son plantas herbáceas que crecen a partir de bulbos. Las raíces son contráctiles. Poseen saponinas esteroideas, esteroides venenosos (bufodienolidos y cardenolidos) y canales o células mucilaginosas.
Las hojas se disponen en una roseta basal, son bastante carnosas y mucilaginosas, alternas y espiraladas, simples, de margen entero, con venación paralela, envainadoras en la base, sin estípulas y con pelos simples.
Las flores se hallan dispuestas en inflorescencias indeterminadas (usualmente racimos o también espigas) en la extremidad de un escapo áfilo. Los pedicelos no son articulados.
Las flores son hermafroditas, actinomorfas, muchas veces vistosas. El perigonio está compuesto de 6 tépalos distribuidos en dos ciclos, separados o connados. Cuando se hallan connados, el perigonio presenta forma de campana a tubular. Los tépalos son imbricados y petaloideos. Los colores de la corola pueden ser blanco, amarillo, violeta, azul, marrón e, incluso, negro. Una muestra de la diversidad de la familia para la coloración de las flores se aprecia en las figuras que acompañan al texto.
El androceo está compuesto por 6 estambres (excepcionalmente 3, como en Albuca, por ejemplo), con los filamentos separados a connados, a veces adnatos a los tépalos, muchas veces con apéndices. Las anteras son dorsifijas y la dehiscencia del polen ocurre por aberturas longitudinales. El polen es monosulcado.
El gineceo es de ovario súpero, tricarpelar, connado y trilocular. Presenta un solo estigma, capitado a 3-lobado. Pueden contener desde uno a varios óvulos en cada lóculo.
Poseen nectarios en los septos de los ovarios.
El fruto es una cápsula loculicida.
Las semillas presentan una morfología muy diversa, desde globosas hasta aplanadas, ocasionalmente con estructuras de tipo arilo. La cubierta seminal usualmente presenta fitomelaninas y las capas internas comprimidas o colapsadas. El endosperma es oleoso.
El tamaño cromosómico es muy variable, desde 1,2 a 18 µm de longitud, con cariotipos bi- o tri-modales. El número cromosómico básico también es muy variable (X= 2, 6, 7, 10, 15, 17, etc.).[3] [4]
Ecología
Las hiacintáceas se hallan ampliamente distribuidas en Europa, África y Asia, en hábitats templados a tropicales, pero son más diversas en áreas de clima mediterráneo (es decir, con una estación seca pronunciada durante el verano). Las vistosas flores de muchas de las especies de la familia son polinizadas por una amplia gama de insectos (abejas, avispas, moscas, polillas) y de pájaros, y tanto el néctar como el polen son ofrecidos como recompensa de la polinización.
Se reproducen tanto en forma sexual como asexual. En este último caso, la multiplicación puede ser vegetativa (a través de los bulbillos que crecen al lado del bulbo madre) o a través de semillas (por apomixis).
La dispersión de las semillas puede ocurrir por agua, viento, o por medio de hormigas ya que muchas especies presentan elaiosomas que ofrecen como recompensa.
Filogenia
Existen todavía profundos desacuerdos entre los especialistas acerca de los límites de la familia, el número de géneros que posee y la inclusión de algunos taxones, por lo que en los próximos años seguramente van a existir modificaciones en la taxonomía de Hyacinthaceae, cuando los datos citogenéticos, morfológicos y moleculares arrojen más luz sobre el tema.
 Eucomis autumnalis, una espectacular especie de África. El "penacho" de hojas por sobre la inforescencia se denomina "coma", de donde proviene el nombre del género.
Eucomis autumnalis, una espectacular especie de África. El "penacho" de hojas por sobre la inforescencia se denomina "coma", de donde proviene el nombre del género.La monofilia de Hyacinthaceae está sostenida por datos moleculares, los que también dan sostén a la exclusión de Camassia, Chlorogalum, y afines, es decir la subfamilia Chlorogaloideae (Chase et al. 1995a,[5] Speta 1998[6] ). Los géneros mencionados se consideran actualmente como miembros herbáceos de Agavaceae, y se distinguen de Hyacinthaceae sensu stricto por su cariotipo bimodal, la combinación de semillas más o menos globosas con una testa firmemente adherida, los tépalos usualmente multinervados y los estigmas distintivamente 3-lobados.
La ubicación exacta de Hyacinthaceae dentro de los Asparagales es poco clara, pero puede estar relacionada con Themidaceae, y estas dos familias pueden estar asociadas con las Agavaceae o, menos probablemente, con el grupo formado por (Alliaceae y Amaryllidaceae) (Chase et al. 1995a,[5] 2000,[7] Rudall et al. 1997a,[8] b,[9] Fay et al. 2000,[10] Soltis et al. 2000[11] ). Esta falta de certeza en la ubicación de la familia dentro del orden Asparagales hace que la determinación de las sinapomorfías morfológicas sea problemática, si bien la presencia de bulbos y esteroides venenosos es notable. La inflorescencia indeterminada (usualmente racimosa o una espiga) fácilmente distingue a las Hyacinthaceae de Themidaceae, Alliaceae, y Amaryllidaceae, todos los cuales tienen inflorescencias umbeladas. El hábito herbáceo y la presencia de bulbo, las distingue de Agavaceae. Ver Asparagales para una discusión sobre los clados que incluyen a Hyacinthaceae.
Taxonomía
- Introducción teórica en Taxonomía
Subfamilias
 Inflorescencia de Ornithogalum dubium .
Inflorescencia de Ornithogalum dubium .Hyacinthaceae ha sido subdividida en 4 subfamilias monofiléticas, las más grandes de las cuales son Ornithogaloideae (con brácteas grandes, núcleo con cristales proteínicos) y Hyacinthoideae (con brácteas usualmente pequeñas, sin cristales proteínicos. Los límites genéricos son controversiales (Manning et al. 2004,[2] Pfosser y Speta 1999,[12] Wetsching y Pfosser 2003[13]
Las cuatro subfamilias, sus sinónimos y algunas de sus características más diferenciales relacionadas con el androceo, la semilla y el número cromosómico son:[2] [12] [13]
- Oziroëeae M. W. Chase, Reveal & M. F. Fay, incluye al género Oziroe, distribuido en el oeste de Sudamérica, con el androceo connado y adnato a la corola, semillas redondeadas y el embrión tan largo como la semilla. Los números cromosómicos básicos son n=15 y n=17.
- Ornithogaleae Rouy (sin.: Ornithogalaceae Salisbury), distribuida en Europa, el oeste de Asia y África. El androceo con 3 estambres aplanados y con apéndices, las semillas aplanadas y angulosas, con la testa firmemente adherida. Los números cromosómicos básicos van desde n=2 a n=10. Esta subfamilia incluye a los géneros Ornithogalum, Albuca, Dipcadi, Galtonia, Neopatersonia y Pseudogaltonia.
- Urgineeae Rouy (sin.: Drimyidaceae Baillon), distribuida principalmente en África, el Mediterráneo hasta India y Madagascar. Presentan semillas aplanadas, aladas con la testa escasamente adherida al endosperma. Los números cromosómicos básicos son n=6, 7 y 10. Las urgineóideas incluyen a los géneros Drimia, Bowiea, Urginea, Litanthus, Rhadamanthus, Schizobasis, Tenicroa e Igidia.
- Hyacintheae Dumortier, con las hojas con pústulas o manchas, semillas redondeadas, con elaiosomas. La gran variabilidad de esta subfamilia permite a su vez subdividirla en tres tribus:
 Lachenalia orchioides en su hábitat natural, adviértanse las manchas oscuras en las hojas, típicas de la subfamilia de las hyacintóideas. Fotografía tomada en Ciudad del Cabo, Sudáfrica, en enero de 2007.
Lachenalia orchioides en su hábitat natural, adviértanse las manchas oscuras en las hojas, típicas de la subfamilia de las hyacintóideas. Fotografía tomada en Ciudad del Cabo, Sudáfrica, en enero de 2007.- Pseudoprospero Manning & Goldblatt, tribu monotípica del sur de África. La única especie de la tribu, Pseudoprospera firmifolium, presenta 2 óvulos por carpelo, 1 semilla por lóculo y número básico n=9.
- Massoniinae Bentham & Hooker f., distribuida en África del sur hasta el Sahara e India. Con 2 o más óvulos por carpelo, elaiosomas en las semillas y números básicos 5 a 10. Los géneros que se incluyen en esta tribu son Daubenya, Drimiopsis, Eucomis, Lachenalia (con 110 especies), Ledebouria (con 80 especies), Massonia, Merwilla, Schizocarphus, Veltheimia y Whiteheadia.[14]
- Hyacinthinae Parlatore (sin.: Scillaceae Vest), se distribuye en Europa, el Mediterráneo y el norte de África. Caracterizada por 2 a 8 óvulos por carpelo, elaiosomas presentes en las semillas y números básicos 4 a 8. En esta tribu se incluyen los géneros Amphisiphon, Androsiphon, Bellevalia (con 50 especies), Chionodoxa, Hyacinthoides (sin.: Endymion), Hyacinthus, Muscari (50 especies), Neobakeria, Polyxena, Próspero (25 especies), Puschkinia, Scilla (con 30 especies) y Thuranthos.
Un cladograma que representa las relaciones entre las 4 subfamilias es el siguiente:
Oziroëoideae
Géneros
Algunos géneros tradicionalmente reconocidos como independientes, han sido reducidos a sinonimia, Botryanthus ha sido incluido en Muscari, Brachyscypha en Lachenalia, Neobakeria Schltr. en Massonia Thunb. ex L.f., Resnova Van der Merwe en Drimiopsis Lindl. & Paxton, Schizocarphus Van der Merwe en Scilla L., Elsiea F.M.Leight. en Ornithogalum L., Endymion Dumort. en Hyacinthoides Heist. ex Fabr. y Urgineopsis Compton en Urginea Steinh.
 Glory of the Snow ("Gloria de la nieve") es el nombre vulgar de Chionodoxa luciliae en los países de habla inglesa.
Glory of the Snow ("Gloria de la nieve") es el nombre vulgar de Chionodoxa luciliae en los países de habla inglesa.Los géneros Camassia y Chlorogalum, hasta hace poco tiempo reconocidos dentro de Hyacinthaceae, actualmente han sido dispuestos en la familia Agavaceae. Asimismo, se ha propuesto que los géneros Dipcadi, Albuca, Neopatersonia, Galtonia y Pseudogaltonia sean incluidos dentro de Ornithogalum.
Los géneros más representados son Ornithogalum (200 especies), Drimia (100 especies), Albuca (50 especies), Muscari (50 especies), y Scilla (30 especies).
Los siguientes géneros son aceptados dentro de la familia
- Albuca
- Alrawia
- Androsiphon
- Bowiea
- Brimeura
- Chionodoxa
- × Chionoscilla
- Daubenya (sin.: Amphisiphon). A.M.van der Merwe & J.C.Manning in Strelitzia, 9: 713 (2000)
- Dipcadi
- Drimia
- Drimiopsis
- Eucomis
- Galtonia
- Hastingsia
- Hyacinthella
- Hyacinthoides
- Hyacinthus
- Lachenalia
- Ledebouria
- Leopoldia
- Massonia
- Muscari
- Neopatersonia
- Ornithogalum
- Pseudogaltonia
- Pseudomuscari
- Puschkinia
- Schizobasis
- Schoenolirion
- Scilla
- Veltheimia
- Whiteheadia
Importancia económica
Charybdis marítima Speta (sin. Urginea marítima), ha sido utilizada como planta medicinal desde tiempos inmemoriales. De hecho, su empleo para el tratamiento de edemas está mencionada en un papiro de 1554 a. C. del Imperio Medio de Egipto. Los compuestos bufadienolidos aislados de C. maritima y Urginea indica se utilizan para la producción de sustancias para el tratamiento de afecciones cardíacas.
Varias especies son tóxicas. En Sudáfrica, por ejemplo, Eliokarmos thyrsoides, Ledebouria cooperi, L. inguinata, L. ovatifolia, L. revoluta, Zahariadia saundersiae y varios miembros de la subfamilia Urgineoideae son ponzoñosos para el ganado. El compuesto tóxico scillirosido (un bufadienolido) se utiliza para envenenar ratas.
Las hiacintáceas se utilizan sólo ocasionalmente como plantas alimenticias para el humano. En Grecia, los bulbos de Muscari comosum se consumían como encurtidos. En Francia, la inflorescencia de Loncomelos pyrenaicus se consume como una hortaliza. En África, algunas tribus consumen los bulbos de Ledebouria apertiflora y' 'Ledebouria revoluta.
No obstante todo lo anterior, la mayor importancia de esta familia radica en su uso como plantas ornamentales y en floricultura. Varias especies de los géneros Chouardia, Hyacinthoides, Hyacinthus, Muscari, Othocallis, Puschkinia y Scilla, se cultivan como ornamentales en parques y jardines, floreciendo en la primavera. En Sudáfrica las especies de Eucomis, Galtonia, Veltheimia, entre otras, se cultivan como ornamentales. Eliokarmos thyrsoides (sin.: Ornithogalum thyrsoides) y los distintos cultivares de jacinto, son importantes en el mercado de flor cortada.[15]
Véase también
Referencias citadas
- ↑ Judd, W. S.; C. S. Campbell, E. A. Kellogg, P. F. Stevens, M. J. Donoghue (2007). «Hyacinthaceae». Plant Systematics: A Phylogenetic Approach, Third edition.. Sunderland, Massachusetts: Sinauer Associates. pp. 269-270. ISBN 978-0-87893-407-2.
- ↑ a b c Manning, J. C.; Goldblatt, P., y M. F. Fay (2004). «A revised generic synopsis of Hyacintheaceae in sub-Saharan Africa, based on molecular evidence, including new combinations and the new tribe Pseudoprospereae.». Edinburgh J. Bot. (60): pp. 533-568. http://journals.cambridge.org/action/displayAbstract;jsessionid=695E522B7131A3E5953D4584531C2836.tomcat1?fromPage=online&aid=214959. Consultado el 25 de febrero de 2008.
- ↑ Error en la cita: El elemento
<ref>no es válido; pues no hay una referencia con texto llamadaAPW - ↑ a b Watson, L.; Dallwitz, M. J.. «Hyacinthaceae» (en inglés). The families of flowering plants: descriptions, illustrations, identification, and information retrieval. Version: 1st June 2007.. Consultado el 4 de noviembre de 2007.
- ↑ a b Chase, M. W.; Duvall, M. R., Hills, H. G., Conran, J. G., Cox, A. V., Eguiarte, L. E., Hartwell, J., Fay, M. F., Caddick, L. R., Cameron, K. M., y Hoot, S. (1995). «Molecular systematics of Lilianae.». En Rudall, P. J., Cribb, P. J., Cutler, D. F.. Monocotyledons: Systematics and evolution. (Royal Botanic Gardens edición). Kew. pp. 109-137.
- ↑ Speta, F. (1998). «Hyacinthaceae.». En Kubitzki, K.. The families and genera of vascular plants, vol 3, Monocotyledons: Lilianae (except Orchidaceae). (Royal Botanic Gardens edición). Berlin: Springer-Verlag. pp. 261-285.
- ↑ Chase, M. W.; Soltis, D. E., Soltis, P. S., Rudall, P. J., Fay, M. F., Hahn, W. H., Sullivan, S., Joseph, J., Molvray, M., Kores, P. J., Givnish, T. J., Sytsma, K. J., y Pires, J. C. (2000). «Higher-level systematics of the monocotyledons: An assessment of current knowledge and a new classification.». En Wilson, K. L. y Morrison, D. A.. Monocots: Systematics and evolution. (CSIRO Publ. edición). Collingwood, Australia. pp. 3-16.
- ↑ Rudall, P.; Furness, C. A., Chase, M. W., y Fay, M. F. (1997a). «Microsporogenesis and pollen sulcus type in Asparagales (Lilianae).». Canad. J. Bot. (75): pp. 408-430. http://cat.inist.fr/?aModele=afficheN&cpsidt=10769694. Consultado el 25 de febrero de 2008.
- ↑ Rudall, P.; Chase, M. W., y Conran, J. G. (1997b). «New circunscriptions and a new family of asparagoid lilies: genera formerly included in Anthericaceae.». Kew Bull. (51): pp. 667-680.
- ↑ Fay, M. F. (2000). «Phylogenetic studies of Asparagales based on four plastid DNA regions.». En K. L. Wilson y D. A. Morrison. Monocots: Systematics and evolution. (Royal Botanic Gardens edición). Kollingwood, Australia: CSIRO. pp. 360-371.
- ↑ Soltis DE; PS Soltis, MW Chase, ME Mort, DC Albach, M Zanis, V Savolainen, WH Hahn, SB Hoot, MF Fay, M Axtell, SM Swensen, LM Prince, WJ Kress, KC Nixon, y JS Farris. (2000). «Angiosperm phylogeny inferred from 18S rDNA, rbcL, and atpB sequences.». Bot. J. Linn. Soc. (133): pp. 381-461. http://www.ingentaconnect.com/content/ap/bt/2000/00000133/00000004/art00380. Consultado el 25 de febrero de 2008.
- ↑ a b Pfosser, M.; Speta, F. (1999). «Phylogenetics of Hyacinthaceae based on plastid DNA sequences.». Ann. Missouri Bot. Gard. (86): pp. 852-875. http://links.jstor.org/sici?sici=0026-6493(199923)86:4%3C852:POHBOP%3E2.0.CO;2-L. Consultado el 25 de febrero de 2008.
- ↑ a b Wetsching,W.; Pfosser, M. (2003). «The Scilla plumbea puzzle - present status of the genus Scilla sensu lato in southern Africa and description of Spetaea lachenaliiflora, a new genus and species of Massonieae (Hyacinthaceae).». Taxon (52): pp. 75-91. http://links.jstor.org/sici?sici=0040-0262(200302)52%3A1%3C75%3ATSPPSO%3E2.0.CO%3B2-3. Consultado el 25 de febrero de 2008.
- ↑ M. Pfosser; , W. Wetschnig, S. Ungar & G. Prenner (2004). «Phylogenetic relationships among genera of Massonieae (Hyacinthaceae) inferred from plastid DNA and seed morphology». Journal of Plant Research 116 (2): pp. 115-132. http://www.springerlink.com/content/xmn09lv45ka601ej/. Consultado el 25 de abril de 2008.
- ↑ Martin Pfosser & Franz Speta (2001 en adelante). «Hyacinthaceae» (en inglés). Consultado el 28 de abril de 2008.
Bibliografía general
- Judd, W. S.; C. S. Campbell, E. A. Kellogg, P. F. Stevens, M. J. Donoghue (2007). «Hyacinthaceae». Plant Systematics: A Phylogenetic Approach, Third edition.. Sunderland, Massachusetts: Sinauer Associates. pp. 269-270. ISBN 978-0-87893-407-2.
- APG. 2003. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG II. Botanical Journal of the Linnean Society 141:399-436.
- J. Manning, P. Goldblatt & M.F. Fay. 2004. A revised generic synopsis of Hyacinthaceae in sub-Saharan Africa, including new combinations and the new tribe Pseudoprospereae. Edinburgh Journal of Botany 60(3): 533-568.
- Mabberley, D. 1997. The Plant-Book. Cambridge University Press, Cambridge, United Kingdom. 858p.
- Stevens, P. F. (Versión 8, Junio 2007, y actualizado desde entonces.). «Hyacinthaceae». Angiosperm Phylogeny Website. Consultado el 28 de abril de 2008.
Enlaces externos
 Wikimedia Commons alberga contenido multimedia sobre Scilloideae. Commons
Wikimedia Commons alberga contenido multimedia sobre Scilloideae. Commons Wikiespecies tiene un artículo sobre Scilloideae. Wikispecies
Wikiespecies tiene un artículo sobre Scilloideae. Wikispecies
- Watson, L.; Dallwitz, M. J.. «Hyacinthaceae» (en inglés). The families of flowering plants: descriptions, illustrations, identification, and information retrieval. Version: 1st June 2007.. Consultado el 4 de noviembre de 2007.









Wikimedia foundation. 2010.