- Apomixis
-
Apomixis

En Botánica, se denomina apomixis o apomixia a la reproducción asexual por medio de semillas. Las plantas que presentan este tipo de reproducción (las que se denominan plantas apomícticas) producen sus semillas sin que ocurra meiosis ni fertilización, por lo que sus descendientes son genéticamente idénticos a la planta madre.[1] Aunque desde el punto de vista evolutivo las plantas apomícticas carecen de las ventajas adaptativas que ofrece la reproducción sexual, la apomixis permite la fijación indefinida de genotipos altamente adaptados a su ambiente. Esta ventaja de la apomixis es -desde el punto de vista genético- la misma que presenta la multiplicación vegetativa. No obstante, en el caso de las plantas apomícticas los descendientes no permanecen en las inmediaciones de la planta madre, compitiendo con ella por recursos, sino que gracias a la dispersión de las semillas los nuevos individuos pueden explorar y conquistar nuevos ambientes.[2] [3]
Contenido
Historia
La apomixis fue descrita por primera vez en 1841 en la especie australiana Alchornea ilicifolia cuando un ejemplar femenino de esta especie dioica fue introducido a los Jardines Botánicos de Kew de Londres. La planta aislada floreció y produjo semillas en abundancia si bien no disponía de ninguna fuente de polen para polinizarse, poniendo en evidencia que la misma había producido semilla asexualmente. Paradójicamente, los primeros experimentos con plantas apomícticas fueron realizados en forma involuntaria por Gregor Mendel, quien utilizó cruzas entre especies del género Hieracium para intentar confirmar los resultados obtenidos en sus famosos estudios sobre la herencia de las arvejas de jardín. Mendel atribuyó erróneamente a una supuesta "frecuente autopolinización" la falta de segregación observada. Hoy se sabe que muchas especies de este género de compuestas son apomícticas.[3]
Tipos de apomixis
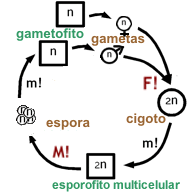 Diagrama esquemático del ciclo de vida haplo-diplonte (con alternancia de generaciones multicelulares).
Diagrama esquemático del ciclo de vida haplo-diplonte (con alternancia de generaciones multicelulares).
Referencias:
n : generación haploide (gametofítica),
2n : generación diploide (esporofítica),
m! : mitosis,
M! : meiosis,
F! : fecundación
círculo: primer estadio del ciclo de vida, unicelular
cuadrado: estadios siguientes del ciclo de vida, multicelularesLa sexualidad de las plantas comprende la alternancia cíclica entre los estados de esporofito (la planta misma, diploide) y de gametofito (el grano de polen y el saco embrionario, ambos haploides). La meiosis que ocurre en las flores posibilita la recombinación y reducción del número de cromosomas y da lugar a la formación de las esporas femeninas (megásporas) en el ovario y masculinas (micrósporas) en las anteras. La megasporogénesis genera cuatro células haploides, tres de las cuales degeneran. La restante constituye la megáspora funcional, que por el proceso de megagametogénesis (tres mitosis sucesivas) desarrolla el megagametófito femenino, también conocido como saco embrionario. El tipo de saco embrionario más común entre las angiospermas (conocido como tipo Polygonum) está formado por 8 núcleos haploides y siete células, a saber: la ovocélula u oósfera, dos sinérgidas, una célula central binucleada, y tres antípodas. Por otra parte, las micrósporas desarrollan los granos de polen mediante un proceso de microgametogénesis. El polen maduro está típicamente integrado por tres células haploides (n), dos de las cuales constituyen los gametos masculinoso núcelos generativos. La otra, denominada célula vegetativa, tiene una función relacionada con el crecimiento del tubo polínico.
La formación de la semilla en las plantas sexuales involucra el proceso de doble fecundación único en las angiospermas: un gameto masculino (n) se fusiona con la ovocélula (n) para originar al cigoto (2n). A partir de este cigoto se desarrolla el embrión. La célula central del saco embrionario con sus dos núcleos polares (n + n) se fusiona con el otro núcleo generativo del grano de polen para originar el endosperma. La fusión de dos gametos haploides únicos derivados de la distribución al azar del material genético durante las meiosis masculina y femenina resulta en la generación de progenies genéticamente diversas. En resumen, en la reproducción sexual la meiosis y la fecundación producen la recombinación génica de los caracteres de ambos progenitores y, por lo tanto, son procesos creadores de nueva variabilidad genética.
La apomixis elude la ruta sexual evitando la reducción meiótica y la fecundación. El óvulo desarrolla una semilla cuyo embrión contiene exactamente el mismo genotipo que la planta que lo origina. Los embriones apomícticos pueden formarse a través de una ruta esporofítica o gametofítica. En la primera, también llamada embrionía adventicia, los embriones surgen directamente de una célula somática de la nucela o de los tegumentos del óvulo. Esta forma de apomixis aparece comúnmente en los cítricos, los cuales se convirtieron en un sistema modelo para estudiar el proceso. En la apomixis gametofítica se forman siempre sacos embrionarios que difieren en algunos aspectos del gametófito femenino haploide (n) generado a partir de la megáspora funcional. Su principal diferencia es precisamente el hecho de ser diploides (2n) ya que los núcleos que los conforman no han pasado por el proceso meiótico y por lo tanto no han reducido su número cromosómico. Por eso se dice que estos sacos embrionarios surgen por un proceso de apomeiosis ("sin meiosis"). De acuerdo con el origen de la célula que genera al saco embrionario y al embrión, la apomixis gametofítica puede ser clasificada como: diplosporía, cuando el saco embrionario se origina a partir de la célula madre de la megáspora misma ya sea por mitosis o luego de una falla en la meiosis o aposporía, cuando el saco embrionario se origina directamente por mitosis a partir de una célula somática, usualmente una célula de la nucela.[1]
Los sacos embrionarios, sean éstos apospóricos o diplospóricos, contienen un gameto femenino diploide (2n), la ovocélula, a partir de la cual se desarrolla directamente el embrión por partenogénesis sin que exista fecundación. Así, mientras en el proceso sexual la reducción meiótica se complementa con la fecundación que restaura el nivel de ploidía 2n, en la apomixis gametofítica la ausencia de reducción se complementa con la partenogénesis.[4]
En la apomixis gametofítica la partenogénesis excluye uno de los procesos de la doble fecundación: la unión de los gametos masculinos con los femeninos. Sin embargo no necesariamente se anula la fecundación de los núcleos polares. Aunque en algunos casos el endosperma puede desarrollarse en forma autónoma (sin la unión de un gameto masculino con los núcleos polares del saco embrionario apospórico o diplospórico) en muchas especies apomícticas, como ocurre en la mayoría de las gramíneas tropicales, es necesario que un gameto masculino se fusione con el o los núcleos polares de la célula central del saco embrionario para formar el endospermo. A este proceso se lo llama pseudogamia.[4]
Por lo expuesto, existen tres tipos diferentes de mecanismos de apomixis:[1] [4]
- Diplosporia: la célula madre del saco embrionario o gametófito femenino se desarrolla directamente en un embrión, proceso conocido con el nombre de partenogénesis diploide. El embrión es diploide.
- Aposporia: el saco embrionario tiene su origen en una célula somática de las múltiples que rodean la célula madre del saco embrionario (nucela). El embrión es diploide.
En ambos casos se desarrolla un gametofito pero la meiosis no existe o, en el caso de que se produzca, no tiene consecuencias observables. Por esta razón se llama también a este fenómeno apomixis gametofítica.
- Embrionía adventicia: no se desarrolla saco embrionario. El embrión se desarrolla a partir de células del esporofito diploide (ejemplo, integumento).
Un ejemplo único de la apomixis masculina se ha descubierto en el ciprés de Sáhara, Cupressus dupreziana, donde el núcleo de los embriones derivan enteramente del progenitor masculino (del polen) sin que exista contribución genética del progenitor femenino.[5] [6]
Distribución
La mayoría de las especies apomícticas son facultativas, de modo que pueden producir semilla sexualmente en un sistema balanceado. Los apomícticos obligados se dan particularmente en aquellas especies que son sexualmente estériles, como por ejemplo en las que solo un sexo está presente (Elodea canadensis, Petasites hybridus), donde solo hay un genotipo para los alelos de autoincompatibilidad (Menyanthes trifoliata, Hottonia palustris), o donde la sexualidad no puede funcionar normalmente debido a los disturbios en la meiosis ocasionados por la hibridación interespecífica (Mentha, Mimulus) o por imparipoliploidía (Potentilla anserina, Holcus mollis).[7] [8]
La apomixis se halla ampliamente distribuida en ciertas familias de plantas, tales como las poáceas, las rosáceas y las compuestas. En estas últimas la apomixis es un tipo de reproducción obligada en muchas especies, mientras que en las otras dos familias la apomixis y la reproducción sexual pueden tener lugar, lado a lado, en el mismo individuo (apomixis facultativa).[9] La reproducción apomíctica se ha detectado en varios géneros de compuestas, tales como Achillea, Crepis, Hieracium, Taraxacum, Conyza,[10] Eupatorium, [11] [12] Erigeron,[13] Brachycome,[14] Parthenium,[15] y Árnica.[16] En las gramíneas, la apomixis fue descrita por primera vez en 1933 en una especie de Poa. Desde aquel momento se ha identificado este mecanismo en cientos de especies de poáceas, particularmente en las paníceas y en las andropogóneas. Algunos de los géneros que presentan especies apomícticas, además del ya mencionado Poa, son Capillipedium, Heteropogon, Themeda, Sorghum, Bothriochloa, Dichanthium, Cenchrus, Setaria y Paspalum.[17] [18] [19]
En algunos biomas, la mayor parte de las especies se reproducen por apomixis facultativa. Por ejemplo, el 60% de la flora británica puede reproducirse asexualmente y el 25% de ese grupo son apomícticos facultativos, es decir que combinan la reproducción asexual con una dispersión especializada, de modo tal que escapan de la competencia con sus progenitores.[8]
Control genético
La apomixis es un carácter heredable, pero su control genético no fue aún completamente esclarecido. Tradicionalmente se consideró que sus determinantes básicos podrían haberse originado por mutación y que la mayoría de los otros genes involucrados son similares a los de la sexualidad. A pesar de su amplia distribución en las angiospermas, el carácter no es muy común en los cultivos más comunes o en las especies modelo (tales como arroz y Arabidopsis). Esta condición determinó que los estudios en este campo deban ser realizados en especies silvestres que son poliploides, altamente heterocigotas y con una escasa caracterización genética. La disección de las bases genéticas del carácter es, por lo tanto, dificultosa y compleja. Los estudios de herencia sólo son posibles si pueden cruzarse progenitores completamente sexuales con plantas apomícticas y la progenie F1 segregante puede ser examinada desde el punto de vista citológico y embriológico. Asimismo, todas las progenies pueden ser analizadas por medio de marcadores moleculares para determinar si sus genotipos son exactamente iguales al de la planta madre o bien, son el producto de cruzamientos. Los diferentes estudios desarrollados en las gramíneas coinciden en señalar que tanto la aposporía como la diplosporía parecen estar controladas por un único locus en donde un gen mayor dominante o un grupo de genes ligados y coadaptados, que funcionan como una sola unidad debido a una fuerte supresión de la recombinación genética, son los responsables de la expresión del carácter.[20] [21] En las compuestas, en cambio, la genética de la apomixis parece tener un control más complejo. Así, se demostró que es de herencia monogénica en dos especies de Hieracium,[22] pero se halla determinada por dos genes con segregación independiente en la especie triploide Erigeron annuus.[13]
El modelo genético más simple propuesto para especies tetraploides de Panicum y Brachiaria, supone que la constitución genética de las plantas sexuales sería del tipo nuliplexo (genotipo «aaaa») mientras que los individuos apomícticos serían uniplexos (genotipo «Aaaa»), siendo «A» el alelo dominante responsable de la apomixis. Los cruzamientos de individuos de este tipo generan progenies F1 que segregan en una relación 1:1 de individuos sexuales a individuos. apomícticos. Sin embargo, en este modelo existen ciertas cuestiones que no han sido resueltas. Por ejemplo en el caso de Panicum maximum (que es una especie apomíctica facultativa) si bien se postula que las plantas apomícticas son uniplexas («Aaaa») hasta ahora no se han encontrado en la naturaleza plantas completamente sexuales («aaaa»). Esta situación se repite en otras especies apomícticas. Tampoco se puede explicar el hecho de que siempre que se utilizó como polinizador a una planta apomíctica ésta resultó ser un genotipo «Aaaa».[23] [24] Este modelo genético para la apomixis indica que las plantas apomícticas llevan el potencial para la reproducción sexual, pero en un estado más o menos reprimido, debido a la presencia del alelo dominante para la apomixis. La penetrancia limitada del alelo para apomixis explica la existencia de la apomixis facultativa. Asimismo, la presencia de los alelos recesivos para sexualidad explica porqué el cruzamiento entre dos plantas apomícticas facultativas puede generar una gran cantidad de progenie enteramente sexual. A pesar que la presencia de apomixis puede ajustarse a este modelo, el grado de apomixis depende de las condiciones ambientales o de genes modificadores.[4] [25] [26] En los últimos años la tecnología de marcadores moleculares y los procedimientos de biología molecular han generado una cantidad considerable de conocimientos nuevos sobre las bases moleculares de la apomixis que pueden ser útiles para el futuro aislamiento del/los genes que controlan el carácter. Han sido identificados varios marcadores que cosegregan con la apomixis en distintos géneros de gramíneas como Tripsacum, Pennisetum, Brachiaria y Paspalum. Estos marcadores ligados al carácter pueden emplearse en el estudio de la transmisión del mismo en los programas de mejoramiento de especies apomícticas y eventualmente intentar cualquiera de las estrategias de clonado basadas en el mapeo genético para aislar el/los disparadores del fenómeno. El análisis molecular posibilitó la identificación de una región genómica específica de la aposporía (ASGR, del inglés "apospory-specific genomic region" ) en Pennisetum squamulantum que está ausente en individuos sexuales y donde la recombinación está severamente restringida. En Tripsacum dactyloides, Paspalum simplex y Paspalum notatum se detectaron regiones similares en donde numerosos marcadores aparecen completamente ligados al carácter. La ausencia de recombinación bien podría estar asociada a una estrategia para evitar la dispersión de los factores que necesitan cosegregar estrictamente para determinar la apomixis. La existencia de una región ASGR de baja recombinación puede enmascarar la presencia de varios genes que gobiernan al carácter, haciéndolos aparecer y funcionar como una sola unidad genética. Una caracterización molecular detallada de este segmento cromosómico revelará en el futuro la variedad y el número de genes involucrados. Dado que varios elementos diferencian a la apomixis de la sexualidad (apomeiosis, partenogénesis, pseudogamia y a veces desarrollo autónomo del endospermo) la cuestión de si la apomixis es consecuencia de la expresión de un solo gen mayor o de un grupo de factores ligados y coadaptados es uno de los problemas a resolver en el futuro.[27] [28] [29] [30] [31] [32] [33] Debido a que la supresión de la recombinación se verifica tanto en monocotiledóneas como en dicotiledóneas, puede que sea una característica general del locus para la apomixis. Esto puede estar relacioando a su función, como fuera observado en otro loci complejos que estan involucrados en procesos similares (denominados complejos génicos coadaptados), como el supergen para heterostilia en Primula,[34] el locus S de autoincompatibilidad en Brassica,[35] el locus para tipo de apareamiento en Chlamydomonas[36] y el locus de histocompatibilidad en humanos.[37]
Alternativamente, esa supresión de la recombinación puede ser un producto accidental de la reproducción asexual continuada durante largos períodos de tiempo.[38] En las especies de Pennisetum, por ejemplo, los marcadores que están ligados al alelo para apomixis no pueden ser detectados en los individuos que se reproducen sexualmente lo que indicaría que los apomícticos son hemicigóticos para el locus en cuestión («A---»)[39] [40] [41] [42] o bien, que los alelos son altamente divergentes («Aa'a'a'»).[43]
Taxonomía de organismos apomícticos
 Las zarzas europeas constituyen un complejo apomíctico con innumerable cantidad de biotipos.
Las zarzas europeas constituyen un complejo apomíctico con innumerable cantidad de biotipos.La apomixis es un fenómeno muy común en las plantas. Los géneros en los que existe apomixis son muy variables y es muy difícil identificar taxonómicamente las especies pertenecientes a él ya que muchas de ellas a veces aparecen como clones únicos. La situación se hace aún más difícil cuando la apomixis es facultativa ya que la complejidad en la distribución de las características morfológicas es aún mayor.
Como una planta apomictica es un ser genético idéntico de una generación a la siguiente, cada individuo tiene el carácter verdadero de la especie, cuando se presenta una distinción de uno a otro congénere apomíctico, esta diferencia es mucho más pequeña que la que se da entre las especies de la mayoría de los géneros. Por lo que se ha dado en llamar a estos seres como microespecies'. En algunos géneros, es posible identificar y nombrar a centenares o aún incluso a millares de microspecies, que se pueden agrupar juntas como un agregado de especies, típicamente enumerados en los libros de Floras con la convención "género de especies agg." _ (e.g., la zarza, Rubus fruticosus agg.). Otros ejemplos de apomixis se pueden encontrar en los géneros Crataegus ("espino albar"), Sorbus ("serbales"), Rubus ("zarza" o "zarzamora"), Hieracium y Taraxacum ("diente de león").[44]
Las zarzas europeas pertenecen a uno de los 12 subgéneros de Rubus y ofrecen un interesante ejemplo de la historia del tratamiento taxonómico de los complejos agámicos. Sólo se conocen seis especies diploides y sexuales dentro del complejo, entre ellas las especies mediterráneas Rubus ulmifolius, R. canescens y R. sanctus. Estas especies han sido denominadas "especies primarias" y se consideran los ancestros, conjuntamente con algunas especies hipotéticas que deben haberse extinguido durante la última glaciación, de toda la flora de zarzas europeas. Casi todas las zarzas europeas pertenecen a un gran complejo agámico, poliploide, que consiste en una innumerable cantidad de biotipos. Solo algunos de ellos (en Europa Central, cerca de 300 taxa) han sido formalmente descriptos como especies. Los restantes, son híbridos derivativos o el producto de cierto grado de sexualidad en los apomícticos facultativos. Tales híbridos surgen espontánea y constantemente en la naturaleza y son fijados indefinidamente mediante reproducción apomíctica. Esto indica que existe un sistema bien balanceado y dinámico entre reproducción apomíctica e hibridación que formó, y continúa formando, la mayor parte de la diversidad de las zarzas europeas.[45] [44] [46] El tratamiento taxonómico de este complejo ha variado con el tiempo. Inicialmente, Linneo dispuso a toda la variabilidad del complejo bajo el nombre Rubus fruticosus.[a] Más tarde, numerosos taxónomos comenzaron a describir parte de la variabilidad mediante la creación de cientos de especies, muchas de ellas simples biotipos con una distribución local sumamente restringida. Otro tratamiento utilizado fue el de disponer todos esos biotipos como subespecies dentro de Rubus fruticosus. Luego de dilucidado el origen y mantenimiento de la variabilidad de este complejo en la naturaleza, se comenzó a tratarlo como una unidad taxonómica especial llamada agamoespecie. En el trabajo taxonómico moderno, solo los clones apomícticos estabilizados y con una distribución geográfica amplia son tratados como especies.[45]
Evolución
La amplia distribución taxonómica de la apomixis y la multiplicidad de mecanismos que subyacen en la producción asexual de semillas sugieren que este modo de reproducción se ha originado independientemente en varias ocasiones durante la evolución de las angiospermas. Ha sido observado que las especies apomícticas son usualmente perennes y derivan de ancestros alógamos. También se ha demostrado que la agamospermia está frecuentemente, aunque no invariablemente, asociada con la hibridación interespecífica y la poliploidía.[47] [48] [49] [50] [51] [52]
Una de las primeras teorías para explicar el valor adaptativo de la apomixis es la que indica que este tipo de reproducción es solamente un escape de la esterilidad en poblaciones que, de otro modo, serían estériles. Esta teoría fue propuesta por Cyril Dean Darlington en 1939[53] y apoyada por George Ledyard Stebbins.[49] [50] Esta teoría ganó adhesión entre muchos botánicos debido a que era compatible con la visión de que la reproducción sexual acelera las tasas de evolución adaptativa,[54] y además, porque explicaba la estrecha asociación existente entre apomixis, poliploidía e hibridación interespecífica.[55] De hecho, buena parte de las especies apomícticas de las compuestas son estériles debido a autopoliploidía (las especies triploides Taraxacum officinale y Erigeron annuus, por ejemplo) y a hibridación interespecífica (tal es el caso de Arnica gracilis, una especie híbrida entre A. latifolia y A. cordifolia). Los híbridos interespecíficos frecuentemente presentan problemas de segregación cromosómica durante la meiosis debido a un inadecuado apareamiento cromosómico. Debido a esto son altamente estériles. La ventaja selectiva de la reproducción apomíctica se hace evidente considerando que en estos individuos, de otro modo estériles, se asegura su reproducción y dispersión por medio de semillas. Desde un punto de vista ecológico, las especies apomícticas se distribuyen frecuentemente en hábitats disturbados y son muy exitosos colonizadores, ya que no dependen de la presencia de individuos de la misma especie en las inmediaciones para dejar descendencia.[9]
Jens Christian Clausen fue uno de los primeros en reconocer formalmente que la apomixis, particularmente la apomixis facultativa, no necesariamente conduce a una pérdida de variabilidad genética y de potencial evolutivo. Utilizando una analogía entre el proceso adaptativo y la producción a gran escala de automóviles, Clausen arguyó que una combinación de sexualidad (que permite la producción de nuevos genotipos) y de apomixis (que permite la producción ilimitada de los genotipos más adaptados) podría incrementar, más que disminuir, la capacidad de una especie para el cambio adaptativo.[56]
Véase también
- Partenogénesis, proceso de reproducción asexual que se da en algunos animales.
Notas
Referencias
- ↑ a b c Richards, A.J. (2003), "Apomixis in flowering plants: an overview", Philosophical Transactions of the Royal Society B: Biological Sciences 358 (1434): 1085–1093, doi:, http://journals.royalsociety.org/content/qk3fnvdnr67paudj/fulltext.pdf
- ↑ Sven E. Asker, Lenn Jerling: Apomixis in Plants. CRC Press, Boca Raton 1992, S. 50f. ISBN 0-8493-4545-6
- ↑ a b Whitton, J.; Sears, C.J.; Baack, E.J.; Otto, S.P. (2008), "The Dynamic Nature of Apomixis in the Angiosperms", International Journal of Plant Sciences 169 (1): 169, doi:
- ↑ a b c d Nogler, G.A. (1984a). Gametophytic apomixis. In: Embryology of Angiosperms, B.M. Johri, ed (Berlin: Springer-Verlag), pp. 475–518.
- ↑ Pichot, C., Fady, B., & Hochu, I. 2000. Lack of mother tree alleles in zymograms of Cupressus dupreziana A. Camus embryos. Ann. For. Sci. 57: 17–22.
- ↑ Pichot, C., El Maátaoui, M., Raddi, S. & Raddi, P. 2001. Surrogate mother for endangered Cupressus. Nature 412: 39
- ↑ Richards, A. J. 1997a. Plant breeding systems, 2nd edn. London: Chapman & Hall.
- ↑ a b Richards, A. J. 1997b Why is gametophytic apomixis almost restricted to polyploids? The gametophyte-expressed model. Apomixis News 9, 3–4.
- ↑ a b Universidad de Hamburgo (2001 en adelante). «Types of Propagation. Systems of Recombination» (en inglés). Botanic on line. Consultado el 2008-28-04.
- ↑ Davis, G.L. 1967: Apomixis in the Compositae. Phytomorphology 77:270-7.
- ↑ Bertasso-Borges Maristela Sanches,; Coleman James Robert (2005). «Cytogenetics and embryology of Eupatorium laevigatum (Compositae)» Genet. Mol. Biol. [serial on the Internet].. Vol. 28. n.º (1):. pp. 123-128. ISSN: 00656275. Consultado el 25/05/2008.
- ↑ Rozenblum, E., Sara Maldonado and Celina E. Waisman. 1988. Apomixis in Eupatorium tanacetifolium (Compositae). American Journal of Botany, Vol. 75, No. 3 pp. 311-322
- ↑ a b Noyes, Richard D., Rieseberg, Loren H. 2000. Two Independent Loci Control Agamospermy (Apomixis) in the Triploid Flowering Plant Erigeron annuus. Genetics 155: 379-390.
- ↑ Davis, G.L. 1964. Embryological studies in the compositae. IV. Sporogenesis, gametogenesis, and embryogeny in Brachycome ciliaris (Labill.) Less. Australian Journal of Botany 12, 142–151.
- ↑ Ray, D.T., D.A. Dierig y A.E. Thompson (1990). «Facultative apomixis in guayule as a source of genetic diversity» pp. 245-247. Ed. J. Janick and J.E. Simon (eds.). Portland, Oregón: Timber Press.
- ↑ Wolf, S. y Keith E. (1984). «Arnica gracilis (Compositae), a Natural Hybrid between A. latifolia and A. cordifolia» Systematic Botany. Vol. 9. n.º 1. pp. 12-16.
- ↑ Connor, H.E. 1979. Breeding systems in the grasses: a survey. New Zealand Journal of Botany 17:547-74.
- ↑ Savidan, Y. 2000. Apomixis: Genetics and Breeding. Plant Breeding Reviews, Volume 18, Jules Janick (ed). John Wiley & Sons, Inc. ISBN 0-471-35567-4. Artículo en inglés
- ↑ Kellogg, E. 1990. Variation and Species Limits in Agamospermous Grasses. Systematic Botany 15: 112-123 Resumen en inglés
- ↑ Barcaccia, G., Mazzucato, A., Albertini, E., Zethof, J., Gerats, A., Pezotti, M., and Falcinelli, M. (1998). Inheritance of parthenogenesis in Poa pratensis L.: Auxin test and AFLP linkage analysis support monogenic control. Theor. Appl. Genet. 97, 74–82
- ↑ Petrov, D.F., Belousova, N.I., and Fokina, E.S. (1979). Inheritance of apomixis and its elements in corn-Tripsacum hybrids. Genetika 15, 1827–1836.
- ↑ Bicknell, R.A., Borst, N.K., and Koltunow, A.M. (2000). Monogenic inheritance of apomixis in two Hieracium species with distinct developmental mechanisms. Heredity 84, 228–237.
- ↑ Savidan, Y. (2000). Apomixis: Genetics and Breeding. In: Plant Preeding Reviews, volume 18. J. Janick (Ed.). John Wiley & Sons, Inc. London.
- ↑ Vielle-Calzada, J.P.; Crane, C.F.; Stelly, D.M. (1996) Apomixis: The Asexual Revolution. Science 274:1322-1323
- ↑ Nogler, G.A. (1984b). Genetics of apospory in apomictic Ranunculus auricomus. V. Conclusion. Bot. Helv. 94, 411–422.
- ↑ Bicknell, R.A., Borst, N.K., and Koltunow, A.M. (2000). Monogenic inheritance of apomixis in two Hieracium species with distinct developmental mechanisms. Heredity 84, 228–237.
- ↑ Martínez, E. J.; Hopp, E.; Sein, J.; Ortiz, J.P. y Quarín, C.L. (2003) Genetic characterization of apospory in tetraploid Paspalum notatum based on the identification of linked molecular markers. Molecular Breeding 12: 319-327.
- ↑ NOGLER, G.A. (1984) Gametophytic Apomixis. In: Johri BM (ed) Embryology of Angiosperms. Springer-Verlag, Berlin.
- ↑ Pessino, S.C.; Ortiz, J.P.A.; Hayward, M.D. y Quarín, C.L. (1999). «The molecular genetics of gametophytic apomixis» Hereditas. Vol. 130. n.º 1. pp. 1-11.
- ↑ Ortiz JPA, Pessino SC, Leblanc O, Hayward MD, Quarín CL (1997). «Genetic fingerprinting for determining the mode of reproduction in Paspalum notatum, a subtropical apomictic forage grass» Theor. Appl. Genet.. Vol. 95.
- ↑ Ortiz JPA, SC Pessino, V Bhat, MD Hayward, CL Quarin (2001). A genetic linkage map of diploid Paspalum notatum. Crop Sci. 41: 823-830.
- ↑ Pessino SC, Espinoza F, Martínez EJ, Ortiz JPA, Valle EM, Quarín CL (2001). «Isolation of cDNA clones differentially expressed in flowers of apomictic and sexual Paspalum notatum» Hereditas. Vol. 134. pp. 35-42.
- ↑ Pessino SC, Evans C, Ortiz JPA, Armstead I, do Valle CB, Hayward MD (1998) A genetic map of the apospory region in Brachiaria hybrids: identification of two markers closely associated with the trait. Hereditas 128: 153-158.
- ↑ Ernst, A. (1936). Heterostylie-Forschung: Versuche zur genetischen Analyse eines Organisations und "Anpassungs" Merkmales. Z. Indukt. Abstammungs. Vererbungsl. 71, 156–230.
- ↑ Awadalla, P. y Charlesworth, D. (1999). «Recombination and selection at Brassica self-incompatibility loci» 152. pp. 413–425.
- ↑ Ferris, P.J., and Goodenough, U. (1994). The mating-type locus of Chlamydomonas reinhardtii contains highly rearranged DNA sequences. Cell 76, 1135–1145.
- ↑ O'hUigin, C., Satta, Y., Hausmann, A., Dawkins, R.L. y Klein, J. (2000). «The implications of intergenic polymorphism for major histocompatibility complex evolution» Genetics. Vol. 156. pp. 867–877.
- ↑ Welch, D.M., and Meselson, M. (2000). Evidence for the evolution of bdelloid rotifers without sexual reproduction or genetic exchange. Science 288, 1211–1215.
- ↑ Roche, D., Cong, P., Chen, Z., Hanna, W.W., Gustine, D.L., Sherwood, R.T. y Ozias-Akins, P. (1999). «An apospory-specific genomic region is conserved between buffelgrass (Cenchrus ciliaris L.) and Pennisetum squamulatum Fresen» The Plant Journal. Vol. 19. n.º 2. pp. 203–208.
- ↑ Goel, Shailendra, Chen, Zhenbang, Conner, Joann A., Akiyama, Yukio, Hanna, Wayne W., Ozias-Akins, Peggy Delineation by Fluorescence in Situ Hybridization of a Single Hemizygous Chromosomal Region Associated With Aposporous Embryo Sac Formation in Pennisetum squamulatum and Cenchrus ciliaris. Genetics 2003 163: 1069-1082
- ↑ Goel, Shailendra, Chen, Zhenbang, Akiyama, Yukio, Conner, Joann A., Basu, Manojit, Gualtieri, Gustavo, Hanna, Wayne W., Ozias-Akins, Peggy. Comparative Physical Mapping of the Apospory-Specific Genomic Region in Two Apomictic Grasses: Pennisetum squamulatum and Cenchrus ciliaris. Genetics 2006 173: 389-400
- ↑ Joann A. Conner, Shailendra Goel, Gunawati Gunawan, Marie-Michele Cordonnier-Pratt, Virgil Ed Johnson, Chun Liang, Haiming Wang, Lee H. Pratt, John E. Mullet, Jeremy DeBarry, Lixing Yang, Jeffrey L. Bennetzen, Patricia E. Klein, and Peggy Ozias-Akins. Sequence Analysis of Bacterial Artificial Chromosome Clones from the Apospory-Specific Genomic Region of Pennisetum and Cenchrus. Plant Physiol. 147: 1396-1411; Plant Physiol.
- ↑ Suzuki, G., Kai, N., Hirose, T., Fukui, K., Nishio, T., Takayama, S., Isogai, A., Watanabe, M. y Hinata, K. (1999). «Genomic organization of the S locus: Identification and characterization of genes in SLG/SRK region of S9 haplotype of Brassica campestris (syn. rapa)» Genetics. Vol. 153. pp. 391–400.
- ↑ a b Richards, A.J.; Kirschner, J.; Stepanek, J.; Marhold, K. (1996), "Apomixis and taxonomy: an introduction", Folia Geobotanica 31 (3): 281–282, doi:, http://www.springerlink.com/index/5722475036555077.pdf
- ↑ a b c Weber, H.E. (1996), "Former and modern taxonomic treatment of the apomictic Rubus complex", Folia Geobotanica 31 (3): 373–380, doi:, http://www.springerlink.com/index/E24Q712516338163.pdf
- ↑ Evans KJ, Symon DE, Roush RT. 1998. Taxonomy and genotypes of the Rubus fruticosus L. aggregate in Australia. Plant Protection Quarterly. 13: 152-156.
- ↑ Gustaffson, A., 1947. Apomixis in higher plants. III. Biotype and species formation. Lunds, Univ. Arsskr., 44, 183-370.
- ↑ Knox. R. B. 1967. Apomixis. Seasonal and population differences in a grass. Science, 157,325—326.
- ↑ a b Stebbins, G. L. 1941. Apomixis in the angiosperms. Bot Rev., 10, 507-542.
- ↑ a b Stebbins, G. L. 1950. Variation and Evolution in Plants, 643 pp. Columbia University Press, New York.
- ↑ Grant, V. 1971. Plant Speciation, 435 pp. Columbia University Press, New York.
- ↑ de Wet, J. M., & Stalker, H. T. 1974. Gametophytic apomixis and evolution in plants. Taxon, 23, 689-697.
- ↑ Darlington, C. D. 1939. The Evolution of Genetic Systems. Cambridge University Press,Cambridge.
- ↑ Crow, J. F., & Kimura, M. 1965. Evolution in sexual and asexual populations. Amer.Nat., 99, 439-450.
- ↑ de Wet. J. M., & Stalker, H. T. 1974. Gametophytic apomixis and evolution in plants. Taxon, 23, 689-697.
- ↑ Clausen, J. 1954. Partial apomixis as an equilibrium system in evolution. Caryologia Suppl.,6, 469-479.
Enlaces externos
Categoría: Reproducción vegetal


Wikimedia foundation. 2010.