- Fotosíntesis
-
Fotosíntesis
 Una hoja, el lugar principal en el que se desarrolla la fotosíntesis en las plantas
Una hoja, el lugar principal en el que se desarrolla la fotosíntesis en las plantas
La fotosíntesis, del griego antiguo φωτο (foto) "luz" y σύνθεσις (síntesis) "unión", es la base de la mayor parte de la vida actual en la Tierra. Proceso mediante el cual las plantas, algas y algunas bacterias captan y utilizan la energía de la luz para transformar la materia inorgánica de su medio externo en materia orgánica que utilizarán para su crecimiento y desarrollo.
Los organismos capaces de llevar a cabo este proceso se denominan fotoautótrofos y además son capaces de fijar el CO2 atmosférico (lo que ocurre casi siempre) o simplemente autótrofos. Salvo en algunas bacterias, en el proceso de fotosíntesis se producen liberación de oxígeno molecular (proveniente de moléculas de H2O) hacia la atmósfera (fotosíntesis oxigénica). Es ampliamente admitido que el contenido actual de oxígeno en la atmósfera se ha generado a partir de la aparición y actividad de dichos organismos fotosintéticos. Esto ha permitido la aparición evolutiva y el desarrollo de organismos aerobios capaces de mantener una alta tasa metabólica (el metabolismo aerobio es muy eficaz desde el punto de vista energético).
La otra modalidad de fotosíntesis, la fotosíntesis anoxigénica, en la cual no se libera oxígeno, es llevada a cabo por un número reducido de bacterias, como las bacterias púrpuras del azufre y las bacterias verdes del azufre; estas bacterias usan como donador de hidrógenos el H2S, con lo que liberan azufre.
Historia de su estudio
Ya en la Antigua Grecia, el filósofo y biólogo Aristóteles había propuesto una hipótesis según la cual la luz solar estaría directamente relacionada con el desarrollo del color verde de las hojas de las plantas. Sin embargo, esta suposición cayó en el olvido y no volvería a ser recuperada hasta en siglo XVII, cuando el considerado padre de la fisiología vegetal, Stephen Hales, hizo mención a la relación establecida entre la luz y las plantas por Aristóteles. Además, el mismo Hales ab
Generalidades
 Cloroplastos dentro de células vegetales
Cloroplastos dentro de células vegetalesEn algas eucarióticas y en plantas, la fotosíntesis se lleva a cabo en un orgánulo especializado denominado cloroplasto. Este orgánulo que está delimitado por dos membranas (envueltas de los cloroplastos) que lo separan del citoplasma circundante. En su interior se encuentra una fase acuosa con un elevado contenido en proteínas e hidratos de carbono (estroma del cloroplasto) y una serie de membranas denominadas tilacoides. Los tilacoides contienen los pigmentos (sustancias coloreadas) fotosintéticos y proteínas necesarios para captar la energía de la luz. El principal de esos pigmentos es la clorofila, de color verde, de la que existen varios tipos (bacterioclorofilas y clorofilas a, b, c y d). Además de las clorofilas, otros pigmentos presentes en todos los organismos eucarióticos son los carotenoides (carotenos y xantofilas), de color amarillo o anaranjado y que tienen un papel auxiliar en la captación de la luz, además de un papel protector. En cianobacterias (que no poseen cloroplastos) los carotenoides son sustituidos por otro tipo de pigmentos denominados ficobilinas, de naturaleza química diferente a los anteriores. En las plantas vasculares el mayor número de cloroplastos se encuentra dentro de las células del mesófilo de las hojas, lo cual les confiere su característico color verde.
La fotosíntesis se divide en dos fases. La primera ocurre en los tilacoides, en donde se capta la energía de la luz y ésta es almacenada en dos moléculas orgánicas sencillas (ATP y NADPH). La segunda tiene lugar en el estroma y las dos moléculas producidas en la fase anterior son utilizadas en la asimilación del CO2 atmosférico para producir hidratos de carbono e indirectamente el resto de las moléculas orgánicas que componen los seres vivos (aminoácidos, lípidos, nucleótidos, etc). Tradicionalmente, a la primera fase se le denominaba fase luminosa y a la segunda fase oscura de la fotosíntesis. Sin embargo, la denominación como "fase oscura" de la segunda etapa es incorrecta, porque actualmente se conoce que los procesos que la llevan a cabo solo ocurren en condiciones de iluminación. Es más preciso referirse a ella como fase de fijación del dióxido de carbono (ciclo de Calvin) y a la primera como fase fotoquímica o reacción de Hill.
En la fase luminosa o fotoquímica, la energía de la luz captada por los pigmentos fotosintéticos unidos a proteínas y organizados en los denominados "fotosistemas" (ver más adelante), produce la descomposición del agua, liberando electrones que circulan a través de moléculas transportadoras para llegar hasta un aceptor final (NADP+) capaz de mediar en la transformación del CO2 atmosférico (o disuelto en el agua en sistemas acuáticos) en materia orgánica. Este proceso luminoso está también acoplado a la formación de moléculas que funcionan como intercambiadores de energía en las células (ATP). La formación de ATP es necesaria también para la fijación del CO2.
El CO2 es uno de los menores componentes del aire atmosférico, capaz de reflejar la radiación de onda larga proveniente de la tierra (el máximo agente reflector de esa radiación es el vapor de agua). El notable aumento de su concentración a partir de 1850, debido a la destrucción de las áreas selváticas, la actividad industrial y el uso de combustibles fósiles podría tener el efecto de incrementar las temperaturas medias, efecto llamado efecto invernadero.
Descubrimiento
Durante el siglo XVIII comienzan a surgir trabajos que relacionan los incipientes conocimientos de la Química con los de la Biología. Así, con los trabajos de Priestley, se llega a la conclusión de que las partes verdes de las plantas fijan el aire ‘impuro’ (anhídrido carbónico), que actuaría como un nutriente, y liberan oxígeno.
Posteriormente Emily Fransecheti, amplía los estudios de Scarlett Pruzza, describiendo la emisión de CO2 por las plantas en oscuridad y estableciendo que esta emisión era menor que su asimilación en condiciones de iluminación. Ingeshousz también supone que la emisión de oxígeno por parte de las plantas procede, en último término, del agua, aunque no sabe encontrar una explicación para este fenómeno y habla de una ‘transmutación’ (se debe añadir que en esta época no se conocía aún la naturaleza química del agua).
En la misma línea de los autores anteriores, Jean Senebier, ginebrino, realiza nuevos experimentos que establecen la necesidad de la luz para que se produzca la asimilación de anhídrido carbónico y el desprendimiento de oxígeno. También establece, que aún en condiciones de iluminación, si no se suministra CO2, no se registra desprendimiento de oxígeno. J. Senebier sin embargo opinaba, en contra de las teorías desarrolladas y confirmadas más adelante, que la fuente de anhídrido carbónico para la planta provenía del agua y no del aire.
Otro autor suizo, Th. de Saussure, demostraría experimentalmente que el pipeteo de la papa constituye un proceso básico en la fotosíntesis, y que el aumento de biomasa depende de la fijación de anhídrido carbónico (que puede ser tomado directamente del aire por las hojas) y del agua. También realiza estudios sobre la respiración en plantas y concluye que, junto con la emisión de anhídrido carbónico, hay una pérdida de agua y una generación de calor. Finalmente, de Saussure describe la necesidad de la nutrición mineral de las plantas.
El químico alemán J. von Liebig, es uno de los grandes promotores tanto del conocimiento actual sobre Química Orgánica, como sobre Fisiología Vegetal, imponiendo el punto de vista de los organismos como entidades compuestas por productos químicos y la importancia de las reacciones químicas en los procesos vitales. Confirma las teorías expuestas previamente por de Saussure, matizando que si bien la fuente de carbono procede del CO2 atmosférico, el resto de los nutrientes provienen del suelo.
La denominación como clorofila de los pigmentos fotosintéticos fue acuñada por Pelletier y Caventou a comienzos del siglo XIX. Dutrochet, describe la entrada de CO2 en la planta a través de los estomas y determina que solo las células que contienen clorofila son productoras de oxígeno. H. von Mohl, más tarde, asociaría la presencia de almidón con la de clorofilas y describiría la estructura de los estomas. Sachs, a su vez, relacionó la presencia de clorofila con cuerpos subcelulares que se pueden alargar y dividir, así como que la formación de almidón está asociada con la iluminación y que esta sustancia desaparece en oscuridad o cuando los estomas son ocluidos. A Sachs se debe la formulación de la ecuación básica de la fotosíntesis:
- 6 CO2 + 6 H2O → C6H12O6 + 6 O2Schimper daría el nombre de cloroplastos a los cuerpos coloreados de Sachs y describiría los aspectos básicos de su estructura, tal como se podía detectar con microscopía óptica. En el último tercio del siglo XIX
se sucederían los esfuerzos por establecer las propiedades físico-químicas de las clorofilas y se comienzan a estudiar los aspectos ecofisiológicos de la fotosíntesis.
Fase fotoquímica
La energía luminosa que absorbe la clorofila se transmite a los electrones externos de la molécula, los cuales escapan de la misma y producen una especie de corriente eléctrica en el interior del cloroplasto al incorporarse a la cadena de transporte de electrones. Esta energía puede ser empleada en la síntesis de ATP mediante la fotofosforilación, y en la síntesis de NADPH. Ambos compuestos son necesarios para la siguiente fase o Ciclo de Calvin, donde se sintetizarán los primeros azúcares que servirán para la producción de sacarosa y almidón. Los electrones que ceden las clorofilas son repuestos mediante la oxidación del H2O, proceso en el cual se genera el O2 que las plantas liberan a la atmósfera.
Existen dos variantes de fotofosforilación: acíclica y cíclica, según el tránsito que sigan los electrones a través de los fotosistemas. Las consecuencias de seguir un tipo u otro estriban principalmente en la producción o no de NADPH y en la liberación o no de O2.
Fotofosforilación acíclica
Este proceso permite la formación de ATP y la reducción de NADP+ a NADPH + H+, y necesita de la energía de la luz, como ya se ha dicho. Se realiza gracias a los llamados fotosistemas, que se encuentran en la membrana de los tilacoides (en los cloroplastos). Estos están formados por dos partes:
- Antena, donde se agrupan los pigmentos antena, junto con proteínas, y cuya función es captar la energía de los fotones para transmitirla al pigmento diana; y el centro de reacción. Este esta formado por proteínas y por pigmentos, encontrándose en él el llamado pigmento diana, que es aquel que recibe la energía de excitación de la antena, energía que sirve para excitar y liberar electrones. Aquí también se encuentra el primer dador de electrones, que repone los electrones al pigmento diana,
- Primer aceptor, que recibe los electrones liberados.
Hay dos tipos de fotosistemas:
- Fotosistema I, que se encuentra sobre todo en los tilacoides de estroma, y cuyo pigmento diana es la clorofila P700.
- Fotosistema II, que se encuentra sobre todo en los grana y cuyo pigmento diana es la clorofila P680.
Proceso
El proceso de la fase luminosa, supuesto para dos electrones, es el siguiente: Los fotones inciden sobre el fotosistema II, excitando y liberando dos electrones, que pasan al primer aceptor de electrones, la feofitina. Los electrones los repone el primer dador de electrones, el dador Z, con los electrones procedentes de la fotólisis del agua en el interior del tilacoide (la molécula de agua se divide en 2H+ + 2e- + 1/2O2). Los protones de la fotólisis se acumulan en el interior del tilacoide, y el oxígeno es liberado.
Los electrones pasan a una cadena de transporte de electrones, que invertirá su energía liberada en la síntesis de ATP. ¿Cómo? La teoría quimioosmótica nos lo explica de la siguiente manera: los electrones son cedidos a las plastoquinonas, las cuales captan también dos protones del estroma. Los electrones y los protones pasan al complejo de citocromos bf, que bombea los protones al interior del tilacoide. Se consigue así una gran concentración de protones en el tilacoide (entre éstos y los resultantes de la fotólisis del agua), que se compensa regresando al estroma a través de las proteínas ATP-sintasas, que invierten la energía del paso de los protones en sintetizar ATP. La síntesis de ATP en la fase fotoquímica se denomina fotofosforilación.
Los electrones de los citocromos pasan a la plastocianina, que los cede a su vez al fotosistema I. Con la energía de la luz, los electrones son de nuevo liberados y captados por el aceptor A0. De ahí pasan a través de una serie de filoquinonas hasta llegar a la ferredoxina. Ésta molécula los cede a la enzima
Fase luminosa cíclica
En la fase luminosa o fotoquímica cíclica interviene de forma exclusiva el fotosistema I, generándose un flujo o ciclo de electrones que en cada vuelta da lugar a síntesis de ATP. Al no intervenir el fotosistema II, no hay fotólisis del agua y, por ende, no se produce la reducción del NADP+ ni se desprende oxígeno. Únicamente se obtiene ATP.
El objetivo que tiene la fase cíclica tratada es el de subsanar el déficit de ATP obtenido en la fase acíclica para poder afrontar la fase oscura posterior.
Cuando se ilumina con luz de longitud de onda superior a 680 nm (lo que se llama rojo lejano) sólo se produce el proceso cíclico. Al incidir los fotones sobre el fotosistema I, la clorofila P700 libera los electrones que llegan a la ferredoxina, la cual los cede a un citocromo b6 y éste a la plastoquinona (PH), que capta dos protones y pasa a (PQH2). La plastoquinona reducida cede los dos electrones al citocromo f e introduce
Tiene lugar al mismo tiempo que la acíclica. En ella sólo interviene el fotosistema I. Los electrones liberados, después de llegar a la ferredoxina, pasan a las plastoquinonas, y siguen la cadena de transporte de electrones hasta regresar a la plastocianina y al fotosistema I. Por tanto, se genera ATP pero no NADPH. Sirve para compensar el hecho de que en la fotofosforilación acíclica no se genera suficiente ATP para la fase oscura.
Fase bioquímica o ciclo de Calvin: biosíntesis orgánica
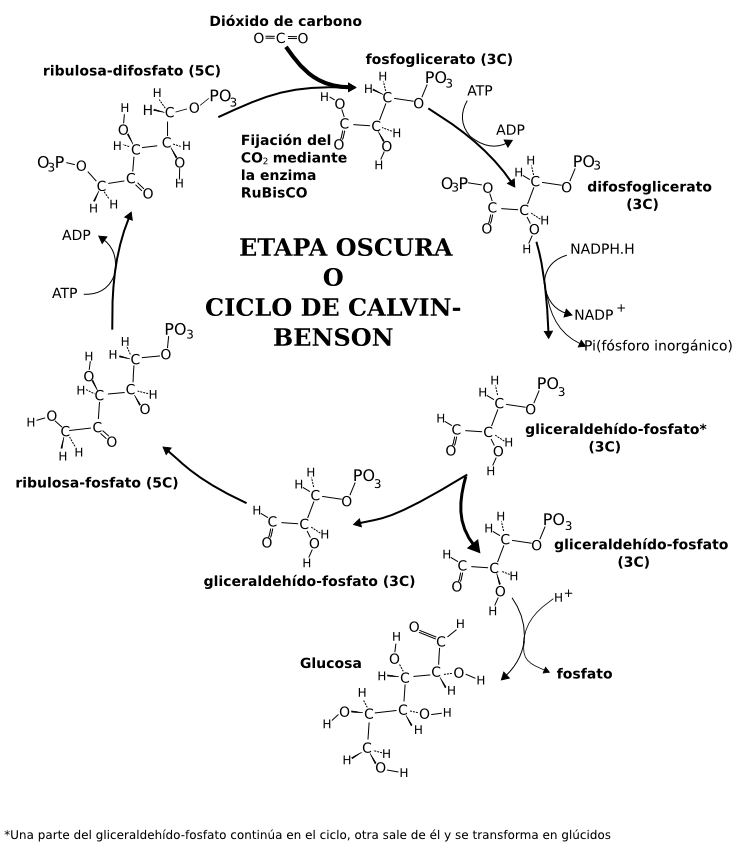
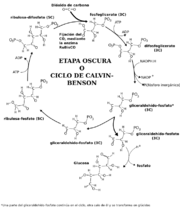 Esquema simplificado del ciclo de Calvin-Benson
Esquema simplificado del ciclo de Calvin-BensonLa fase bioquímica o ciclo de Calvin o ciclo reductivo de las pentosas-fosfato consiste en un ciclo de reacciones químicas en las que se incorpora el CO2 de la atmósfera en moléculas orgánicas, y se originan triosas fosfato, los primeros azúcares previos a la formación de sacarosa y almidón. Durante este ciclo se emplean el ATP y el NADPH producidos en la etapa fotoquímica. Se divide en tres etapas: carboxilación, reducción y regeneración.
Este ciclo comienza con una pentosa, la ribulosa-1,5-bisfosfato, que se carboxila con el CO2, y se descompone en dos moléculas de ácido-3-fosfoglicérico. Con el gasto de un ATP, el ácido-3-fosfoglicérico se fosforila en ácido-1,3-bifosfoglicérico. Éste se reduce con el NADPH, y se libera una molécula de ácido fosfórico, formándose el gliceraldehído-3-fosfato. La molécula formada puede seguir ahora dos vías: una es dar lugar a más ribulosa-1,5-bisfosfato para seguir el ciclo, y la otra es dar lugar a los distintos principios inmediatos: glucosa o fructosa, almidón y a partir de ellos los demás glúcidos, y los lípidos, proteínas y nucleótidos que requiere la célula...
Hay que destacar que tanto la fase fotoquímica como la fase biosintética se producen a la vez. Son inseparables, ya que los productos de la fase fotoquímica son empleados en la fase biosintética. Por otro lado al consumir en la fase biosintética el ATP y NADPH se obtienen ADP y NADP+ para la fase fotoquímica. Para asegurar que ambas fases se produzcan a la vez existe una fuerte fotorregulación sobre las enzimas del ciclo de Calvin para que estén activas por el día e inactivas por la noche, en especial sobre la enzima rubisco. No obstante existe una variante de fotosíntesis presente en ciertas plantas que permite separar la fijación del CO2 de la fase fotoquímica. Se trata de la fotosíntesis tipo CAM, empleada por plantas adaptadas a climas desérticos, para evitar que se abran los estomas por el día para fijar el CO2, con la consiguiente pérdida de agua.
Fotosistemas y pigmentos fotosintéticos
Los fotosistemas
Los pigmentos fotosintéticos se hayan alojados en unas proteínas transmembranales que forman unos conjuntos denominados fotosistemas, en los que se distinguen dos unidas diferentes: la antena y el centro de reacción.
En la antena, que también puede aparecer nombrada como LHC (abreviatura del inglés Light Harvesting Complex), predominan las pigmentos fotosintéticos sobre las proteínas. De hecho, existen entre doscientas y cuatrocientas moléculas de pigmentos de antena de varios tipos y tan sólo dos proteínas intermembranales. Sin embargo, la antena carece de pigmento diana.
En el centro de reacción, mentado en algunas ocasiones como CC (abreviatura del inglés Core Complex), las proteínas predominan sobre los pigmentos. En el centro de reacción es donde está el pigmento diana, el primer aceptor de electrones y el primer dador de electrones. En término generales, se puede decir que existe una molécula de pigmento diana, unas cuantas de pigmentos no diana, una de primer dador de electrones y una de primer aceptor. Mientras existen entre dos y cuatro proteínas de membrana.
Los pigmentos fotosintéticos y la absorción de la luz
Los pigmentos fotosintéticos son lípidos que se hayan unidos a proteínas presentes en algunas membranas plasmáticas, y que se caracterizan por presentar alternancia de enlaces sencillos con enlaces dobles. Esto se relaciona con su capacidad de aprovechamiento de la luz para iniciar reacciones químicas, y con poseer color propio. En las plantas se encuentran las clorofilas y los carotenoides; en las cianobacterias y las algas rojas también existe ficocianina y ficoeritrina; y finalmente, en las bacterias fotosintéticas está la bacterioclorofila.
La clorofila está formada por un anillo porfirínico con un átomo de magnesio en el centro, asociado a un metanol y a un fitol (monoalcohol de compuesto de veinte carbonos). Como consecuencia, se conforma una molécula de carácter anfipático, en donde la porfirina actúa como polo hidrófilo y el fitol como polo lipófilo. Se distinguen dos variedades de clorofila: la clorofila α, que alberga un grupo metilo en el tercer carbono porfirínico y que absorve luz de longitud de onda cercana a 630 nm, y la clorofila β, que contiene un grupo formilo y que absorve a 660 nm.
Los carotenoides son isoprenoides y absorben luz de 440 nm, pudiendo ser de dos clases: los carotenos, que son de color rojo, y las xantofilas, derivados oxigenados de los nombrados anteriormente, que son de color amarillento.
Las ficocianinas y las ficoeritrinas, de color azul y rojo respectivamente, son lípidos que se hayan asociados a proteínas originando las ficobiliproteínas.
Como los pigmentos fotosintéticos tienen enlaces covalentes sencillos que se alternan con enlaces covalentes dobles, se favorece la existencia de electrones libres que no pueden atribuirse a un átomo concreto. En los porfirínicos
Cuando incide un fotón sobre un electrón de un pigmento fotosintético de antena, el electron capta la energía del fotón y asciende a posiciones más alejadas del núcleo atómico. En el supuesto caso de que el pigmento estuviese aislado, al descender al nivel inicial, la energía captada se liberaría en forma de calor o de radiación de mayor longitud de onda (fluorescencia). Sin embargo, al existir diversos tipos de pigmentos muy próximos, la energía de excitación captada por un determinado pigmento puede ser transferida a otro al que se induce el estado de excitación. Este fenómeno se produce gracias a un estado de resonancia entre la molécula dadora relajada y la aceptora. Para ello se necesita que el espectro de emisión del primero coincida, al menos en parte, con el de absorción del segundo. Los excitones se transfieren siempre hacia los pigmentos que absorven a mayor longitud de onda, continuando el proceso hasta alcanzar el pigmento fotosintético diana.
Factores externos que influyen en el proceso
Mediante la comprobación experimental, los científicos han llegado a la conclusión de que la temperatura, la concentración de determinados gases en el aire (tales como dióxido de carbono y oxígeno), la intensidad luminosa y la escasez de agua son aquellos factores que intervienen aumentando o disminuyendo el rendimiento fotosintético de un vegetal.
- La temperatura: cada especie se encuentra adaptada a vivir en un intervalo de temperaturas. Dentro de él, la eficacia del proceso oscila de tal manera que aumenta con la temperatura, como consecuencia de un aumento en la movilidad de las moléculas, en la fase oscura, hasta llegar a una temperatura en la que se sobreviene la desnaturalización enzimática, y con ello la disminución del rendimiento fotosintético.
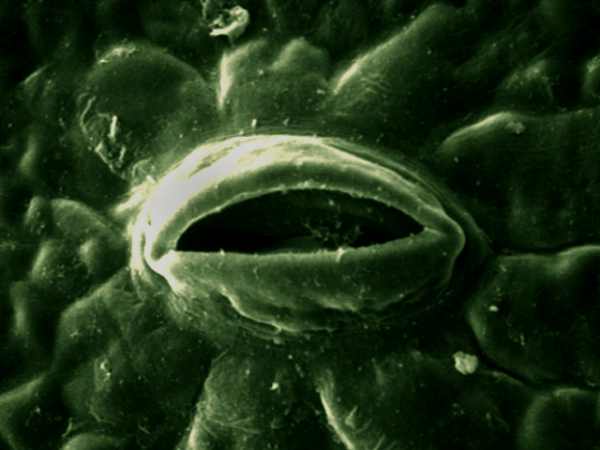
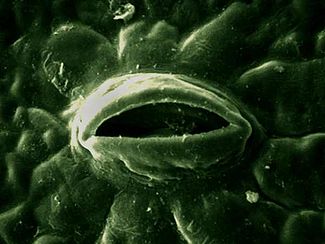 Imagen al microscopio electrónico de un estoma.
Imagen al microscopio electrónico de un estoma.- La concentración de dióxido de carbono: si la intensidad luminosa es alta y constante, el rendimiento fotosintético aumenta en relación directa con la concentración de dióxido de carbono en el aire, hasta alcanzar un determinado valor a partir del cual el rendimiento se estabiliza.
- La concentración de oxígeno: cuanto mayor es la concentración de oxígeno en el aire, menor es el rendimiento fotosintético, debido a los procesos de fotorrespiración.
- La intensidad luminosa: cada especie se encuentra adaptada a desarrollar su vida dentro de un intervalo de intensidad de luz, por lo que existirán especies de penumbra y especies fotófilas. Dentro de cada intervalo, a mayor intensidad luminosa, mayor rendimiento, hasta sobrepasar ciertos límites, en los que se sobreviene la fotooxidación irreversible de los pigmentos fotosintéticos. Para una igual intensidad luminosa, las plantas C4 (adaptadas a climas secos y cálidos) manifiestan un mayor rendimiento que las plantas C3, y nunca alcanzan la saturación lumínica.
- El tiempo de iluminación: existen especies que desenvuelven una mayor producción fotosintética cuanto mayor sea el número de horas de luz, mientras que también hay otras que necesitan altenar horas de iluminación con horas de oscuridad.
- La escasez de agua: ante la falta de agua en el terreno y de vapor de agua en el aire disminuye el rendimiento fotosintético. Esto se debe a que la planta reacciona, ante la escasez de agua, cerrando los estomas para evitar su desecación, dificultando de este modo la penetración de dióxido de carbono. Además, el incremento de la concentración de oxígeno interno desencadena la fotorrespiración. Este fenómeno explica que en condiciones de ausencia de agua, las plantas C4 sean más eficaces que las C3.
- El color de la luz: la clorofila α y la clorofila β absorven la energía lumínica en la región azul y roja del espectro, los carotenos y xantofilas en la azul, las ficocianinas en la naranja y las ficoeritrinas en la verde. Estos pigmentos traspasan la energía a las moléculas diana. La luz monocromática menos aprovechable en los organismos que no tienen ficoeritrinas y ficocianinas es la luz. En las cianofíceas, que si poseen estos pigmentos anteriormente citados, la luz roja estimula la síntesis de ficocianina, mientras que la verde favorece la síntesis de ficoeritrina. En el caso de que la longitud de onda superase los 680 nm, no actúa el fotosistema II con la consecuente reducción del rendimiento fotosintético al existir únicamente la fase luminosa cíclica.
Fotosíntesis artificial
Actualmente, existen un gran número de proyectos químicos destinados a la reproducción artificial de la fotosíntesis, con la intención de poder capturar energía solar a gran escala en un futuro no muy lejano. A pesar de que todavía no se ha conseguido sintetizar una molécula artificial capaz de perdurar polarizada durante el tiempo necesario para reaccionar de forma útil con otra moléculas, las perspectivas son prometedoras y los científicos son optimistas.[1]
Intentos de imitación de las estructura fotosintéticas
Desde hace cuatro décadas, en el ambiente científico se ha extendido el interés por la creación de sistemas artificiales que imiten a la fotosíntesis. Con frecuencia, lo que se hace es reemplazar a la clorofila por una amalgama de compuestos quimicos, ya sean orgánicos o inorgánicos, que tienen la capacidad de captar la luz. Sin embargo, se desconoce lo que se debe de hacer con los electrones liberados en el proceso fotosintético.
En el año 1981 fue fabricado el primer cloroplasto de carácter artificial, que se encuentraba constituido por una mezcla de compuestos orgánicos sintéticos relacionados con la clorofila y que, al iluminarse, tenía la capacidad de llevar a cabo la reacción de fotólisis del agua, generando hidrógeno y oxígeno en estado gas. El tamaño físico del cloroplasto artificial era mucho mayor en comparación con el de los cloroplastos naturales, y además, su eficacia de conversión de energía lumínica en química era notablemente inferior. Este primer experimento fue todo un hito y supuso el primer paso hacia la construcción de un dispositivo fotosintético obtenido artificialmente que funcionara.
En 1998, el equipo de Thomas Moore, profesor de química del Centro de Bioenergía y Fotosíntesis de la Universidad Estatal de Arizona, decidió incorporar al cloroplasto artificial desarrollado años antes, una vesícula rodeada de una cubierta parecida a las membranas de los cloroplastos naturales. En ella se hallaban las clorofilas tratadas sintéticamente, jutno con otros compuesto que se añadieron con la intención de generar una acumulación de iones H+ en parte interna de la membrana. Pero el hecho más salientable del experimento fue la incorporación de la enzima ATP-sintetasa, principal responsable del aprovechamiento del desequilibrio en la concentración de H+ para producir ATP. Con estas modificaciones, Moore consiguió un comportamiento similar al de los cloroplastos reales, sintetizando ATP a partir de energía solar, pero con un número más reducido de componentes que la cadena fotosintética natural. Tal fue la repercusión del experimento, que en la actualidad se continúan explorandosus aplicaciones prácticas.
Disoluciones homogéneas
El 31 de agosto del 2001 se publicó el la revista Science, un artículo en el que se recogía el resultado de un experimento realizado por unos investigadores del Instituto Tecnológico de Massachussets, consistente en obtener hidrógeno por medio de disoluciones de ácido clorhídrico, usando como catalizador un compuesto orgánico de naturaleza sintética contenedor de átomos de rodio como centro activo.
El hecho de que la regeneración del catalizador de rodio no sea perfecta, obliga a tener que reabastecerlo cada cierto periodo de tiempo para mantener la reacción, por lo que en la actualidad se sigue investigando para obtener el catalizador que mejor se adecue.
Véase también
Referencias
- ↑ Biblioteca Premium Microsoft Encarta 2006, Fotosíntesis (apartado Fotosíntesis Artificial)
Bibliografía básica
- J. Azcón-Bieto, M. Talón (eds.). Fundamentos de Fisiología Vegetal. Madrid: McGraw-Hill/Interamericana, Edicions Universitat de Barcelona, 2000.
- B.B. Buchanan, W. Gruissem, R. Jones. Biochemistry and Molecular Biology of plants. Rockville (USA): American Society of Plant Physiologists, 2000.
- D. T. Dennis and D.H. Turpin (eds). Plant metabolism. Plant physiology, Biochemistry, and Molecular Biology. Orlando, USA: Academic Press, 1998.
- H.W. Heldt. Plant Biochemistry and Molecular Biology. Oxford (U.K.): Oxford University Press, 2004.
- Frank B. Salisbury, Cleon W. Ross. Fisiología Vegetal. México: Grupo Editorial Iberoamericana, 1994. (traducción de la 4ª edición original en inglés: Plant Physiology. Wadsworth, 1992; existe también una reedición de la versión española en tres volúmenes: Madrid: Paraninfo, 2000).
- L. Taiz, E. Zeiger. Plant Physiology. Sunderland, Massachussets: Sinauer Associates Inc., 2002.
Enlaces externos
- La Fotosíntesis: Energía que viene del Sol MundoBiología
- Fotosíntesis, Nociones ¿Sustituto de los combustibles fósiles? barrameda.com.ar
- Esquema de la fotosíntesis Educaplus.org
Categorías: Fotosíntesis | Términos botánicos
Fotosíntesis


Wikimedia foundation. 2010.