- Deinonychus
-
Deinonychus
?Deinonychus
Rango fósil: Cretácico inferior
Estado de conservación Fósil Clasificación científica Reino: Animalia Filo: Chordata Clase: Archosauria Superorden: Dinosauria Orden: Saurischia Suborden: Theropoda Familia: Dromaeosauridae Subfamilia: Velociraptorinae Género: Deinonychus Especie: D. antirrhopus Nombre binomial Deinonychus antirrhopus
Ostrom, 1969Deinonychus (gr. "garra terrible") es un género representado por una única especie de dinosaurio terópodo dromeosáurido, que vivió a mediados del período Cretácico, hace aproximadamente 119 y 97 millones de años, entre el Aptiano y el Albiano, en lo que hoy es Norteamérica. Deinonychus era un corredor muy ágil y veloz, cualidades que se veían favorecidas por su cola larga y rígida, siempre levantada, que le ayudaba a mantener el equilibrio durante la carrera. Aunque estaba estrechamente emparentado con el velocirráptor de Mongolia, el deinonico se diferencia fácilmente a simple vista por su morro más corto, constitución más robusta y mayor tamaño. Alcanzaba longitudes de entre 2,5 y 3,5 metros y un peso de 50 a 70 kilogramos.
El nombre “garra terrible” del griego δεινος, "terrible" y ονυξ/ονυχος, "garra", se refiere a la garra inusualmente grande, en forma de hoz en el segundo dedo de cada del pie trasero, que probablemente se levantaba para que no tocara el suelo mientras que el dinosaurio caminaba apoyando en el tercer y cuarto dedo del pie. Históricamente se pensó que el Deinonychus golpearía con el pie de la garra en hoz para realizar grandes cortes en su presa pero las pruebas recientes en reconstrucciones del similar Velociraptor sugieren que las garras la garra se utilizaban para apuñalar, mas que para desgarrar. Las especie se la nombra D. antirrhopus que significa “equilibrio contrario”, que refiere a la idea de John Ostrom sobre la función de la cola. Como en otros dromeosauridós, las vértebras de la cola tienen una serie de tendones osificados y procesos óseos muy largos. Estas características parecían hacer de la cola en un contrapeso tieso, pero un fósil del muy estrechamente vinculado; Velociraptor mongoliensis (IGM 100/986) presenta un esqueleto articulado en el que la cola se curva lateralmente con una forma de S. Esto sugiere que, en vida, la cola podría haberse curvado a los lados con un alto nivel de flexibilidad.[1] Encontrado en las Formaciones Cloverly y Antlers, los restos de Deinonychus fueron encontrados asociado estrechamente a los del ornitópodo Tenontosaurus. Los dientes descubiertos junto a los especímenes Tenontosaurus implican fueron cazados o por lo menos devorados por Deinonychus.
Contenido
Descripción
El mayor espécimen conocido de Deinonychus alcanzó los 3,4 metros de largo, con un cráneo de alrededor de 410 milímetros, una altura de cadera de 87 centímetros y un peso máximo de 73 kilogramos.[2] Poseía una gran cabeza, con alrededor sesenta dientes curvados hacia atrás, con forma de cuchillos. Los estudios del cráneo han progresado mucho durante las últimas décadas. Ostrom lo reconstruyó a partir de ejemplares parciales preservados imperfectamente, describiéndolos como triangulares, amplios, y bastante similares a los del Allosaurus. Material adicional del cráneo de Deinonychus y géneros estrechamente vinculados encontrados con una buena preservación tridimensional[3] muestran que el paladar era más estrecho que lo que pensó Ostrom, haciendo el hocico más alargado, mientras que el yugal, flanqueado por aberturas, daba una mayor visión estereoscópica. El cráneo de Deinonychus era diferente en la forma al de Velociraptor , con un techo del cráneo como el de Dromaeosaurus , y no tenía los nasales deprimidos de Velociraptor.[3] El cráneo y la mandíbula inferior tenían ventanas para reducir el peso del cráneo. En Deinonychus la ventana anteorbital, una abertura del cráneo entre el ojo y las narinas, eran particularmente grandes.[3]
Su cuello arqueado en forma de S, extremidades largas y fornidas y el segundo dedo del pie provisto de la extraordinaria y terrible garra falciforme, que el animal alzaba del suelo para conservar su aguda punta, y cuya función no era otra sino que desgarrar y destripar. El primer dedo estaba atrofiado, el quinto ausente y el tercero y cuarto se usaban para caminar. En las garras delanteras había sólo tres dedos igual de fuertes y coronados por garras curvas del mismo tamaño, el primer dígito era el más corto y el segundo era el más largo.. Cuando no se usaban para aferrar a la presa, estas garras se recogían bajo el brazo, de forma similar a como un ave recoge sus alas.
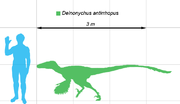 Comparación de tamaño del deinonico con un ser humano.
Comparación de tamaño del deinonico con un ser humano.
Yacimientos con cientos de individuos fosilizados y esqueletos de otros dinosaurios con dientes de deinonico de distinto tamaño incrustados indican claramente que esta especie era un animal social que vivía en manadas. Los deinonicos se valían de su gran número para cooperar en la caza y dar muerte así a animales mucho mayores que ellos. Parece que el gran herbívoro Tenontosaurus de hasta 6,5 metros de longitud (más del doble que un deinonico adulto) era una presa habitual en su dieta.
Tras haber rodeado a la víctima con la ayuda de ingeniosas tácticas de caza, los deinonicos saltaban sobre ella y procedían a desgarrarla con sus uñas afiladas, especialmente la gran garra en forma de hoz de cada pie, que era retráctil y podía golpear bastante fuerte gracias a los potentes músculos que la controlaban. Los dientes los usaban en raras ocasiones para matar a la víctima; los que se han encontrado incrustados sobre los huesos de tenontosaurios seguramente acabaron ahí al devorar a la presa.
Se ignora qué clase de organización habría en las manadas de deinonico, pero al ser depredadores sociales capaces de tramar estrategias de caza, es muy probable que tuviesen una organización jerárquica similar a la de los cánidos o hienas actuales.
Historia
 Antigua visión de Deinonychus sin plumas.
Antigua visión de Deinonychus sin plumas.Los resto fósiles de Deinonychus han sido recobrados de la Formación Cloverly de Montana y Wyoming[4] y de la Formación Antlers de Oklahoma,[5] en Estados Unidos. Adicionalmente, un diente de las capas de arcilla de Cloveryy de la Formación Potomac en la costa atlántica de Maryland ha sido asignado a este género.[6]
Los primeros restos en ser encontrados, se hallaron en 1931 en el sudeste de Montana cerca del pueblo de Bridger. Un grupo liderado por el , paleontólogo Barnum Brown, se concentraron sobre todo en excavar y preparar los restos del ornitópodo Tenontosaurus , pero en su informe de campo sobre el sitio excavación para el Museo Americano de Historia Natural, informo el descubrimiento de un pequeño dinosaurio carnívoro cerca del esqueleto de Tenontosaurus, " pero muy encajado en la roca lo que lo hacia difícil de preparar.[7] Llamó informalmente al animal "Daptosaurus" e hizo preparaciones para describirlo y lo colocó en la exhibición, pero nunca termino este trabajo.[8] Posteriormente B. Brown volvió de la Formación Cloverly con el esqueleto de un terópodo más pequeño con aparentemente los dientes de gran tamaño, al que nombró informalmente como "Megadontosaurus". John Ostrom, reviso el material varias décadas más tarde, identificando a los dientes como pertenecientes a un Deinonychus , el esqueleto proveniente de un animal totalmente distinto al que nombro Microvenator.[8] Un poco más de treinta años más tarde, en agosto de 1964, el paleontólogo John Ostrom llevó a una expedición del Museo Peabody de la Universidad de Yale que descubrió más material esquelético. Las expediciones durante los dos veranos siguientes desenterraron más de 1000 huesos, entre los cuales estaban por lo menos tres individuos. Puesto que la asociación entre los varios huesos recuperados era débil, la fabricación del número exacto de animales individuales se presentó imposible de determinar correctamente, el espécimen tipo (YPM 5205) de Deinonychus fue restringido a un pie izquierdo completo y un pie derecho parcial que pertenecieron definitivamente al mismo individuo.[9] Los especímenes restantes fueron catalogados en cincuenta entradas separadas en el Museo Peabody de Historia Natural deYale .
Posteriores estudios de J. Ostrom y G. E. Meyer analizaron su propio material así como el "Daptosaurus" de B. Brown detalladamente y encontraron que era lo misma especie. J. Ostrom publico su descubrimiento en 1969, dandole a todos los restos referido al nombre de Deinonychus antirrhopus.[9]
Aunque una miríada de huesos estaba disponible antes de 1969, muchos importantes faltaban o eran difíciles de interpretar. Había pocos elementos postorbitales del cráneo, ningún fémures, sacro, espoleta o esternón, vértebras faltantes, y J. Ostrom pensó que solamente un fragmento minúsculo era el coracoides. La reconstrucción esquelética de J. Ostrom de Deinonychus incluyó un hueso pélvico muy inusual, un pubis trapezoidal y plano, diferente al de los otros terópodos, pero que era de la misma longitud que el isquion y que fue encontrada a la derecha de este.
Posteriores descubrimientos
En 1974 J. Ostrom publicó otra monografía sobre el hombro de Deinonychus en cuál corrigió que el pubis que él había descrito era realmente un coracoides, un elemento del hombro.[10] En ese mismo año, otro espécimen de Deinonychus fue excavado en Montana por una expedición de la Universidad de Harvard dirigida por Farish Jenkins. Este descubrimiento agregó varios nuevos elementos; fémures bien preservados, pubis, un sacro, y mejores ilion, así como los elementos del pie y del metatarso.J. Ostrom describió este espécimen y revisó la restauración esquelética de Deinonychus. Esta vez se vieron pubis muy largos, y J. Ostrom comenzó a sospechar que pudieron incluso haber estado orientados hacia cauda poco como en los pájaros.[11]
 Representación del Deinonychus con plumas.
Representación del Deinonychus con plumas.Un esqueleto de Deinonychus que incluye los huesos del especimen original y más completo puede verse en el Museo Americano de Historia Natural,[12] con otro ejemplar en el Museo de Zoología Comparada de la Universidad de Harvard. Los especímenes del Museo Americanos y de Harvard son de un lugar distinto que los de Yale. Incluso estos dos montajes esqueléticos están carecen de elementos incluyendo los esternones, las costillas esternales, la espoleta, y gastralias.
Incluso después de todo el trabajo de J. Ostrom, varios pequeños bloques de material incrustado en la roca seguía estando sin preparación en el almacén del AMNH. Éstos consistieron sobre todo en huesos y fragmentos aislados de huesos, incluidos en la matriz original, o en la roca circundante en la cual los especímenes terminaron enterrados inicialmente. Un examen de estos bloques sin preparación de Gerald Grellet-Tinner y de Peter Makovicky en 2000 reveló una característica interesante, pasada por alto. Varios huesos largos y finos identificados en los bloques como tendones osificados (estructuras que ayudaron a atiesar la cola de Deinonychus) resultado representar realmente gastralias (costillas abdominales). Más perceptiblemente, una gran cantidad de cáscaras de huevo fosilizadas previamente inadvertidas fueron descubiertas en la matriz de la roca que había rodeado el espécimen original de Deinonychus.[13]
En un informe posterior, más detallado sobre las cáscaras de huevo, G. Grellet-Tinner y P. Makovicky concluyeron que los huevos pertenecieron casi ciertamente a Deinonychus , representando el primer huevo del dromeosáurido que se identificará.[7] Por otra parte, la superficie externa de una cáscara de huevo fue encontrada en contacto cercano con las gastralias que sugerriria que ese Deinonychus pudo haber estado incubando sus huevos. Esto implica que Deinonychus realizaba un traspaso térmico del cuerpo como mecanismo para la incubación del huevo, e indica endotermia similar a los pájaros modernos.[14] El estudio adicional de Gregory Erickson y colegas encuentra que este individuo tenia entre 13 o 14 años a la muerte y su crecimiento amesetado. A diferencia de otros de otros terópodos en su estudio de especímenes encontrados asociado con los huevos o nidos, había acabado el crecimiento a la hora de su muerte. [15]
Implicaciones
La descripción de 1969 hecha por J. Ostrom de Deinonychus ha sido descrito como uno de los hechos más importantes de la paleontología del mediados del siglo XX.[16] El hallazgo de los primeros restos de muchos de Deinonychus por los paleontólogos John Ostrom y Grant E. Meyer en 1964, en el sur de Montana, llevaron a Ostrom a suponer que el Deinonychus vivía en manadas, algo impensable para la época, que juzgaba a los dinosaurios tan obligádamente estúpidos que sólo podrían llevar una vida solitaria. También dio con las primeras evidencias de que los deinonico depredaban sobre los Tenontosaurus, algo que, evidentemente, no podrían hacer en solitario.
 Esqueleto de un deinonico.
Esqueleto de un deinonico.Uniendo estas evidencias con su singular anatomía, Ostrom describió al Deinonychus como un animal activo, ágil, veloz y mínimamente inteligente, características que implicaban necesariamente el que fuera un animal de sangre caliente. Tal idea tuvo un fuerte impacto en los círculos académicos y posteriormente en la idea popular sobre los dinosaurios, que llevaron a lo que se ha denominado como Renacimiento de los Dinosaurios, el abandono de los prejuicios decimonónicos sobre los dinosaurios y su estudio sistemático como animales de su tiempo. La avalancha de descubrimientos posteriores que ha dibujado a los dinosaurios como animales bien adaptados, inteligentes, ágiles, de sangre caliente, buenos padres de su prole y capaces de tener vida social, serían impensables si el Deinonychus no hubiese llevado a Ostrom a dar el pistoletazo de salida. Varios años después, J. Ostrom noto las similitudes entre los miembros de Deinonychus con las aves, siendo esta observación lo que le llevo a revivir la hipótesis que las aves descendían de los dinosaurios.[17] Treinta años después esa idea esta mundialmente aceptada.
Deinonychus estaba seguramente emplumado como su cercano pariente Velociraptor, demostrado en 2007 en base de las marcas de las plumas en los brazos. Tales marcas demuestran donde se anclan las plumas en pájaros modernos. Este descubrimiento reciente agregó el peso a las teorías ya fuertes que todos los dromeosáuridos fueron emplumados.[18]
Clasificación
Varios años después, el estudio de los fósiles de Deinonychus permitió a Ostrom apreciar ciertas similitudes entre éste y las aves. Con esto, Ostrom propuso que las aves descendían de los dinosaurios, algo que ya había mencionado Thomas Henry Huxley en el siglo XIX pero que había caído en el olvido desde los años 20 del siglo pasado. Pese a las fuertes críticas de los ornitólogos y paleontólogos de la época, decenas de descubrimientos en los 30 años siguientes han dado la razón a Ostrom. Hoy en día, incluso se conocen numerosos deinonicosaurios y dinosaurios similares con plumas avianas auténticas como Velociraptor o Microraptor, por lo que es muy probable que el propio Deinonychus tuviera también una cubierta plumosa que le ayudase a regular el calor corporal. Deinonicosaurios (Deinonychosauria) y aves (Avialae) están reunidas actualmente en el clado Maniraptora.
 Esqueleto de un D. antirrhopus.
Esqueleto de un D. antirrhopus.Deinonychus es uno de los mejor conocidos entre los Dromaeosauridae,[19] junto al más pequeño y relativamente cercano Velociraptor, del Campaniano de Asia Central.[20] [21] El clado que ellos forman se llama Velociraptorinae. El nombre Velociraptorinae fue por primera vez acuñado por Rinchen Barsbold en 1983[22] y originalmente contenía solo al Velociraptor. Posteriormante Phil Currie incluyo a más dromeosáuridos.[23] Del Cretácico superior a Tsaagan de Mongolia[20] y al norteaméricano Saurornitholestes,[2] pueden también estar entre los familiares cercanos, pero este último es mal conocido y difícil de clasificar.[20] Velociraptor y miran a sus parientes usaron sus garras más que sus cráneos como herramientas de caza, en comparación con dromeosáuridos como Dromaeosaurus de cráneos mas masivos.[24] Junto con Troodontidae, los dromeosáuridos del clado Deinonychosauria son taxones hermanos de las aves. Filogenícamente, Deinonychosauria al grupo de dinosaurios no aviares más cercano a las aves.[25]
Paleoecología
La evidencia geologica sugiere que Deinonychus habitaba una llanura aluvional o zonas pantanosas.[19] El paleoambiente de ambas formaciones, la Cloverly y la de Antlers, en dondes los restos de Deinonychus han sido encontrados, consisten de bosques, deltas fluviales y lagos, no muy diferente a lo que es hoy Louisiana.[26] Otros animales que compartieron este mundo con Deinonychus al herbívoro armado Sauropelta y los ornitópodos Zephyrosaurus y Tenontosaurus. En Oklahoma, el ecosistema de Deinonychus también incluyo al terópodo Acrocanthosaurus, y el gigante Sauroposeidon, el cocodriliano Goniopholis, y el Lepisosteiformes Lepisosteus.[26] Si los dientes encontrados en Maryland son de ' ' Deinonychus' ' , entonces sus vecinos incluirían Astrodon , Dryptosaurus y un nodosáurido, llamado Priconodon conocido solamente por los dientes.
Paleobiología
Comportamiento predador
 cabeza de Deinonychus basado en la reciente reconstrucción del cráneo.
cabeza de Deinonychus basado en la reciente reconstrucción del cráneo.Los dientes de Deinonychus encontrados en asociación con fósiles del dinosaurio ornitópodo Tenontosaurus son muy comunes en la Formación Cloverly. Se han descubierto dos sitios que preservan fósiles bastantes completos de Deinonychus cerca a Tenontosaurus. El primero, el sitio de Yale en Cloverly, Montana, incluye numerosos dientes, cuatro fósiles de Deinonychus adultos y un juvenil. La asociación de este número de esqueletos de Deinonychus en una solo lugar sugiere que Deinonychus puede haberse alimentado de ese animal, y quizás haberlo cazado. J. Ostrom y W. Maxwell incluso han utilizado esta información para especular que Deinonychus pudo haber vivido y cazado en manadas.[27] El segundo Sitio es en la Formación Antlers de Oklahoma. El sitio contiene seis esqueletos parciales de Tenontosaurus de varios tamaños, junto con un esquelético parcial y muchos dientes de Deinonychus. Un húmero del Tenontosaurus incluso tiene lo qué pudo ser marcas de dientes de Deinonychus. D. Brinkman et al. . (1998) precisan que Deinonychus tenía un peso adulto de 70 a 100 kilogramos, mientras que los Tenontosaurus adultos pesaban entre 1 a 4 toneladas. Un solitario Deinonychus no podría matar un Tenontosaurus adulto, sugiriendo que la caza en manadas sería posible.[28]
Un estudio reciente por B. T. Roach y D. L. Brinkman han aportado en la pregunta sobre el comportamiento cooperativo durante la caza en manada de Deinonychus , basados en lo que se sabe de la caza de carnívoros modernos y tafonomía de sitios de Tenontosaurus. Los arcosaurios modernos (pájaros y cocodrilos) y el Dragón de Komodo exhiben poco de caza cooperativa, en su lugar, son generalmente cazadores solitarios, o se agrupan en carcasas muertas anteriormente, donde ocurre mucho conflicto entre los individuos de la misma especie. Por ejemplo, en las situaciones donde los grupos de dragones de Komodo están comiendo juntos, los individuos más grandes comen primero y atacarán a Dragones de Komodo más pequeños que intenten alimentarse. Si se mata al animal más pequeño, éste es canibalizado. Cuando esta información se aplica a los sitios de Tenontosaurus, parece que es consistente con lo encontrado entre los Dragones de Komodo y cocodrilos. Los restos esqueléticos de Deinonychus encontrados en estos sitios son de subadultos y las piezas que faltan en ellos son consistentes con la predación por otro Deinonychus.[29]
Función de los miembros
A pesar de ser la característica más distintiva de Deinonychus, la forma y la curvatura en hoz de la garra varía entre los especímenes. El espécimen tipo descrito por J. Ostrom en 1969 tiene una garra fuertemente curvada en hoz, mientras que un espécimen más nuevo descrito en 1976 tenía una garra con una curvatura mucho más débil, más similar en perfil con el de las garras normales en los restantes dedos del pie.[30] J. Ostrom ha sugerido que esta diferencia en el tamaño y la forma de las garras en hoz podría ser debido a la variación individual, sexual, o relativa a la edad.
 Modelo del los huesos del Deinonychus mostrando la garra falciforme, en el Museo de Zoología de Copenhagen.
Modelo del los huesos del Deinonychus mostrando la garra falciforme, en el Museo de Zoología de Copenhagen.J. Ostrom especuló originalmente que Deinonychus agarraba sus presas con las garras de las manos mientras que pateaba desgarrando con sus garras falciformes.[9] Posteriores estudios, sin embargo, han demostrado que las garras falciformes no fueron utilizadas para desgarrar pero si para asestar pequeñas puñaladas a la víctima,[31] posiblemente indicando su uso como crampones para escalar.[32] Estudios biomecánicos Ken Carpenter en 2002 confirmaron que la función más probable de los brazos era la de agarrar la presa, pues sus grandes longitudes habrían permitido un alcance más largo que para la mayoría de los otros terópodos. El grande y alargado coracoides hace suponer la existencia de músculos de gran alcance en los brazos, lo que consolida esta teoría.[33] Los estudios biomecánicos de K. Carpenter usando moldes de los huesos también demostraron que los Deinonychus no podían doblar sus brazos contra su cuerpo como un pájaro, contrariamente lo qué fue deducido en 1985 de las descripciones anteriores por Jacques Gauthier[34] y Gregory S. Paul en 1988.[2]
 Ilustración de Deinonychus antirrhopus en descanso con los brazos doblados.
Ilustración de Deinonychus antirrhopus en descanso con los brazos doblados.
Plumaje basado en géneros relacionados.Los estudios de Phil Senter en 2006 indicaron que los brazos de Deinonychus se podían utilizar no sólo para agarrar sino también para llevar los objetos hacia el pecho. La presencia probable de un ala grande emplumada en Deinonychus , sin embargo, habría limitado en un cierto grado la gama de movimiento de los brazos. Por ejemplo, cuando Deinonychus llevaba sus brazos hacia adelante, la palma de la mano giraba automáticamente a una posición hacia arriba. Esto habría hecho que un ala bloqueara a la otra si ambos brazos estuviesen extendidos al mismo tiempo, llevando P. Senter a concluir eso que lo atraer objetos al pecho habría sido lograda solamente con un brazo a la vez. La función de los dedos también habría estado limitada por las plumas; por ejemplo, solamente el tercer dígito de la mano se habría podido emplear en actividades tales escarbar en las grietas para buscar pequeñas presa, y solamente en perpendicular a la posición del ala.[35] Alan Gishlick, en un 2001 estudio de la mecánica del brazo de Deinonychus, encontró que incluso si las plumas grandes del ala estaban presentes, la capacidad de atrape de la mano no habría sido obstaculizada perceptiblemente. Esta habría sido perpendicular al ala, y habría sido sostenida probablemente por ambas manos simultáneamente en un " abrazo de oso" de manera, hecho que ha sido apoyado por los estudios posteriores del brazo por Carpenter y Senter.[36]
W. Parsons ha demostrado que los especímenes juveniles y subadultos de Deinonychus exhiban algunas diferencias morfológicas con los adultos. Por ejemplo, los brazos de los especímenes más jóvenes eran proporcional más largos que los de los adultos, una indicación posible de la diferencia en comportamiento entre los jóvenes y adultos.[37]
Velocidad
 Buitreraptor y Deinonychus montados en posición de correr.
Buitreraptor y Deinonychus montados en posición de correr.Los dromeosáuridos, especialmente Deinonychus, se representan a menudo como animales inusualmente de rápidos en los medios masivos, y J. Ostrom mismo especuló que Deinonychus era veloz en su descripción original.[9] Sin embargo, cuando el primer ' ' Deinonychus' ' fue descrito, carecia de una pierna completa, y J. Ostrom especulo sobre la longitud del fémur, que más adelante se comprobo que resulto una sobrestimación. En un estudio posterior, J. Ostrom observó que cociente del fémur a tibia no es tan importante en la determinación de velocidad como la longitud relativa del pie y de la parte inferior de la pierna. En los pájaros modernos, como el avestruz, el cociente entre el pie y la tibia es 0,95. En dinosaurios inusualmente de rápidos como el Struthiomimus , el cociente es 0,68, pero en Deinonychus , el cociente es de 0,48. J. Ostrom indicó que "solamente una conclusión es razonable, el Deinonychus no era particularmente rápido comparado con otros dinosaurios, ni con las aves corredoras modernas".[30]
El bajo cociente del miembro posterior distal en Deinonychus es debido en parte a un inusuálmente corto metatarso. El cociente es realmente más grande en individuos más pequeños que en los más grandes. J. Ostrom sugirió que el metatarso corto se pueda relacionar con la función de la garra falciforme, y utilizó el hecho de que parece hacerse más corto mientras que los individuos envejecen como soporte de esto, Interpretó todas estas características, el segundo dedo del pie corto con la garra agrandada, el metatarso corto, etc. como ayuda para el uso de la pierna trasera como arma ofensiva, donde la garra giraría hacia abajo y atras, y de la pierna empujaba hacia abajo y detrás al mismo tiempo, desgarrando en la presa. J. Ostrom sugirió que el metatarso corto redujera la tensión total en los huesos de la pierna durante tal ataque, e interpretó el arreglo inusual de las inserciones musculares en la pierna del Deinonychus como apoyo para su idea que un sistema de músculos distinto fuera utilizado en el movimiento rapaz y en caminar. Por lo tanto, J. Ostrom concluyó que las piernas de Deinonychus representaban un equilibrio entre las adaptaciones corrientes necesarias para un depredador ágil, y las características para compensar su arma única del pie.[30]
En su estudio de 1981 de las huellas de dinosaurio encontrada en Canadá, Richard Kool realizo estimaciones groseras sobre la velocidad que caminan basadas en varios rastros de huellas hechas por diversas especie en la Formación Gething de la Columbia Británica. R. Kool estimo para una de estos rastros de icnospecies, Irenichnites gracilis (que pudo haber sido de hecho Deinonychus), una velocidad al caminar de 2,8 metros por segundo (alrededor de 10,1 km por hora).[38]
Huevos
La identificación en 2000 de un probable huevo de Deinonychus asociado a uno de los especímenes originales permitió la comparación con otros dinosaurios terópodos en términos de estructura del huevo, nido, y reproducción. En su estudio de 2006 del espécimen, G. Grellet-Tinner y P. Makovicky examinaron la posibilidad que los dromeosáuridos se había estado alimentando de los huevos, o que fragmentos había sido asociado al esqueleto de Deinonychus por coincidencia. Descartaron la idea que el huevo había sido una comida para el terópodo, observando que los fragmentos fueron intercalados entre las gastralias y los huesos del brazo, haciéndolo imposible que representaran el contenido estomacal del animal. Además, la manera de la cual el huevo había sido machacado y hecho fragmento indicando que había estado intacto a la hora del entierro, y estaba quebrado por el proceso de fosilización. La idea que el huevo fuera asociado aleatoriamente al dinosaurio también fue encontrada como inverosímil, los huesos que rodeaban el huevo no habían sido dispersados ni desarticulado, sino seguían estando bastante intactos concerniente a sus posiciones en vida, indicando que el área alrededor al huevo no fueron disturbadas durante la preservación. El hecho de que estos huesos fueran gastralias, que muy raramente se encuentran articulados, apoyó esta interpretación. Toda la evidencia, según G. Grellet-Tinner y P. Makovicky, indican que el huevo estaba intacto debajo del cuerpo del Deinonychus cuando fue enterrado. Es posible que éste representa el empollamiento o comportamiento de nido en Deinonychus similar a lo visto en troodóntidos relacionados y oviraptóridos, o que el huevo estaba de hecho en el interior del oviducto cuando murió el animal.[7]
 Deinonychus echado en el suelo
Deinonychus echado en el sueloEl examen de la microestructura del huevo de Deinonychus confirma que perteneció a un terópodo, puesto que comparte características con otros huevos conocidos de terópodos y muestra que diferencias con huevos de ornitisquios y saurópodos. Comparado a otro terópodos maniraptores, el huevo de Deinonychus es más similar a los de oviraptórido que a los de troodóntidos, a pesar de los estudios que demuestran que estos últimos son lo más estrechamente vinculados a los dromeosáuridos como ' ' Deinonychus' '. Mientras que el huevo estaba muy roto para determinar exactamente su tamaño, G. Grellet-Tinner y P. Makovicky estimaban un diámetro alrededor de 7 cm. basado en la anchura del canal pélvico a través del cual el huevo tuvo que haber pasado. Este tamaño es similar al diámetro de 7.2 cm. del más grande huevo de Citipati huevos de un oviraptórido. Citipati y Deinonychus también compartieron el mismo tamaño de cuerpo total, apoyando esta estimación. Además, el grosor de las cáscaras de huevo de Citipati y Deinonychus son casi idénticas, y puesto que el grueso de cáscara se correlaciona con el volumen del huevo, estos datos apoyan la idea que los huevos de estos dos animales eran de tamaño casi igual.[7]
Cultura Popular
El deinonychus aparece en la serie de itv Primeval, en su primera aparición aparece una familia en un centro comercial, mas tarde vemos una pareja cautiva por un villano.
En la tercera temporada de la serie Primeval se le ve en el ultimo episodio, cuando un grupo persigue a los protagonistas.
En la serie de películas y novelas de Parque Jurásico, la especie de dinosaurio conocida como Velocirraptor es en realidad el Deinonychus, ya que el verdadero Velocirraptor es más pequeño y solo vivió en Asia, mientras que en Parque Jurásico encuentran un fósil en Norteamérica, donde vivía el Deinonychus. En el caso de que se filmase "Parque Jurásico 4", se cree que el Deinonico sería llamado por su verdadero nombre en lugar de Velocirraptor y estaría totalmente cubierto de plumas.
También fue visto en la serie transmitida en Disney XD Dino Rey aunque ahí el trío de Deinonicos ninguno tenía plumas.
Referencias
- ↑ Norell, M. A.; & Makovicky, P. J. (1999). «Important features of the dromaeosaurid skeleton II: information from newly collected specimens of Velociraptor mongoliensis» American Museum Novitates. Vol. 3282. pp. 1–45.
- ↑ a b c Paul, G.S. (1988). Predatory Dinosaurs of the World. New York: Simon and Schuster, pp. 366–369.
- ↑ a b c Maxwell, W.D.; &Witmer, L.M. (1996). «New Material of Deinonychus (Dinosauria, Theropoda)» Journal of Vertebrate Paleontology. Vol. 16(3). pp. 51A.
- ↑ Ostrom, J. H. (1970). «Stratigraphy and paleontology of the Cloverly Formation(Lower Cretaceous) of the Bighorn Basin area, Wyoming and Montana» Bulletin of the Peabody Museum of Natural History. Vol. 35. pp. 1–234.
- ↑ Brinkman, D. L.; R. L. Cifelli, and N. J. Czaplewski (1998). «First occurrence of Deinonychus antirrhopus (Dinosauria: Theropoda) from the Antlers Formation (Lower Cretaceous: Aptian-Albian) of Oklahoma» Oklahoma Geological Survey Bulletin. Vol. 146. pp. 1–27.
- ↑ Lipka, T. R. (1998). «The Affinities of the Enigmatic Theropods of the Arundel Clay Facies (Aptian), Potomac Formation, Atlantic Coastal Plain of Maryland», Lucas, S.G., Kirkland, J.I. and Estep, J.W. (ed.). Lower and Middle Cretaceous Terrestrial Ecosystems. Albuquerque: New Mexico Museum of Natural History and Science, pp. 229–234.
- ↑ a b c d Grellet-Tinner, G.; and Makovicky, P. (2006). «A possible egg of the dromaeosaur Deinonychus antirrhopus: phylogenetic and biological implications» Canadian Journal of Earth Sciences. Vol. 43. n.º 6. pp. 705–719. DOI 10.1139/E06-033.
- ↑ a b Norell, M. A.; Gaffney, E. S.; and Dingus, L. (1995). Discovering Dinosaurs in the American Museum of Natural History. New York: Knopf, pp. 126–130. ISBN 0-679-43386-4.
- ↑ a b c d Ostrom, J. H. (1969). «Osteology of Deinonychus antirrhopus, an unusual theropod from the Lower Cretaceous of Montana» Peabody Museum of Natural History Bulletin. Vol. 30. pp. 1–165.
- ↑ Ostrom, John H.,. (1974). " The Pectoral Girdle and Forelimb Function of Deinonychus (Reptilia: Saurischia) : A Correction". Postilla, Peabody Museum of Natural History Bulletin , 165:1-11.
- ↑ Ostrom, J.H. (1976). «On a new specimen of the Lower Cretaceous theropod dinosaur Deinonychus antirrhopus» Breviora. Vol. 439. pp. 1–21.
- ↑ American Museum of Natural History (2007). «Deinonychus». http://www.amnh.org. American Museum of Natural History. Consultado el 2007-07-13.
- ↑ Makovicky, P. J.; Grellet-Tinner, G. (2000). «Association between a specimen of Deinonychus antirrhopus and theropod eggshell», Bravo, A.M. and T. Reyes (ed.). First international symposium on dinosaur eggs and babies,Isona i Conca Dellà Catalonia, Spain, 23–26 September 1999, pp. 123–128.
- ↑ Grellet-Tinner, Gerard (2006). «Oology And The Evolution Of Thermophysiology In Saurischian Dinosaurs: Homeotherm And Endotherm Deinonychosaurians?» Papeis Avulsos de Zoologia. Vol. 46. n.º 1. pp. 1–10.
- ↑ Erickson, Gregory M.; Curry Rogers, Kristina; Varricchio, David J.; Norell, Mark A.; and Xu, Xing (2007). «Growth patterns in brooding dinosaurs reveals the timing of sexual maturity in non-avian dinosaurs and genesis of the avian condition» Biology Letters. Vol. published online. pp. 558. DOI 10.1098/rsbl.2007.0254.
- ↑ Fastovsky, D.E., Weishampel, D.B. (2005). «Theropoda I: Nature Red in Tooth and Claw», Fastovsky, D.E., Weishampel, D.B. (ed.). The Evolution and Extinction of the Dinosaurs, 2nd edition edición, Cambridge: Cambridge University Press, pp. 265–299. ISBN 0-521-81172-4.
- ↑ Ostrom, J. H. (1976). «Archaeopteryx and the origin of birds» Biological Journal of the Linnean Society. Vol. 8. pp. 91–182. DOI 10.1111/j.1095-8312.1976.tb00244.x.
- ↑ Turner, A.H.; Makovicky, P.J.; and Norell, M.A. (2007). «Feather quill knobs in the dinosaur Velociraptor» Science. Vol. 317. n.º 5845. pp. 1721. DOI 10.1126/science.1145076. PMID 17885130.
- ↑ a b Norell, M.A., Makovicky, P.J. (2004). «Dromaeosauridae», Weishampel, D.B., Dodson, P., Osmólska, H. (ed.). The Dinosauria, 2nd edition edición, Berkeley: University of California Press, pp. 196–210. ISBN 0-520-24209-2.
- ↑ a b c Norell, M.A.; Clark, J.M., Turner, A.H., Makovicky, P.J., Barsbold, R., and Rowe, T. (2006). «A new dromaeosaurid theropod from Ukhaa Tolgod (Ömnögov, Mongolia)» American Museum Novitates. Vol. 3545. pp. 1–51. DOI [1:ANDTFU2.0.CO;2 10.1206/0003-0082(2006)3545[1:ANDTFU]2.0.CO;2].
- ↑ Turner, A.S.; Hwang, S.H.; and Norell, M.A. (2007). «A small derived theropod from Öösh, Early Cretaceous, Baykhangor Mongolia» American Museum Novitates. Vol. 3557. pp. 1–27. DOI [1:ASDTFS2.0.CO;2 10.1206/0003-0082(2007)3557[1:ASDTFS]2.0.CO;2].
- ↑ Barsbold, R. (1983). «Carnivorous Dinosaurs from the Cretaceous of Mongolia» The Joint Soviet-Mongolian Palaeontological Expedition, Transactions. Vol. 19. pp. 1–117.
- ↑ Currie, P. J. (1995). «New information on the anatomy and relationships of Dromaeosaurus albertensis (Dinosauria: Theropoda)» Journal of Vertebrate Paleontology. Vol. 15. n.º 3. pp. 576–591. (abstract)
- ↑ Paul, G.S. (1988). Predatory dinosaurs of the world : a complete illustrated guide. New York: Simon and Schuster, pp. 358. ISBN 0-671-61946-2.
- ↑ Benton, M.J. (2004). Vertebrate Palaeontology (Third Edition). Blackwell Publishing, pp. 472. ISBN 9780632056378.
- ↑ a b Wedel, M. J.; Cifelli, R. L. (2005). «Sauroposeidon: Oklahoma’s Native Giant» Oklahoma Geology Notes. Vol. 65. n.º 2. pp. 40–57.
- ↑ Maxwell, W. D.; Ostrom, J.H. (1995). «Taphonomy and paleobiological implications of Tenontosaurus-Deinonychus associations» Journal of Vertebrate Paleontology. Vol. 15. n.º 4. pp. 707–712. (abstract)
- ↑ Brinkman, Daniel L.; Cifelli, Richard L., Czaplewski, Nicholas J. (1998). «"First Occurrence of Deinonychus antirrhopus (Dinosauria: Theropoda) from the Antlers Formation (Lower Cretaceous: Aptain-Albian) of Oklahoma"» Oklahoma Geological Survey. n.º 164. pp. 27.
- ↑ Roach, B. T.; D. L. Brinkman (2007). «A reevaluation of cooperative pack hunting and gregariousness in Deinonychus antirrhopus and other nonavian theropod dinosaurs» Bulletin of the Peabody Museum of Natural History. Vol. 48. n.º 1. pp. 103–138. DOI [103:AROCPH2.0.CO;2 10.3374/0079-032X(2007)48[103:AROCPH]2.0.CO;2].
- ↑ a b c Ostrom, J. H. (1976). «On a new specimen of the Lower Cretaceous theropod dinosaur Deinonychus antirrhopus» Breviora. Vol. 439. pp. 1–21.
- ↑ Carpenter, K. (1998). «Evidence of predatory behavior by carnivorous dinosaurs» Gaia. Vol. 15. pp. 135–144.
- ↑ Manning, P. L.; D. Payne, J. Pennicott, P. M. Barrett, and R. A. Ennos (2006). «Dinosaur killer claws or climbing crampons?» Biology Letters. Vol. 2. n.º 1. pp. 110–112. DOI 10.1098/rsbl.2005.0395. PMID 17148340.
- ↑ Carpenter, K. (2002). «Forelimb biomechanics of nonavian theropod dinosaurs in predation» Senckenbergiana Lethaea. Vol. 82. pp. 59–76.
- ↑ Gauthier, J.; Padian, K. (1985). "Phylogenetic, Functional, And Aerodynamic Analyses Of The Origin Of Birds And Their Flight" in The Beginnings Of Birds. M.K. Hecht, J.H. Ostrom, G. Viohl, & P. Wellnhofer (eds) proceedings of the International Archaeopteryx Conference, Eichstätt, 1984: 185-197, Eischtatt: Freunde des Jura-Museums Eichstätt. isbn 3-9801178-0-4.
- ↑ Senter, P. (2006). «Comparison of Forelimb Function Between Deinonychus And Bambiraptor (Theropoda: Dromaeosauridae)» Journal of Vertebrate Paleontology. Vol. 26. pp. 897–906. DOI [897:COFFBD2.0.CO;2 10.1671/0272-4634(2006)26[897:COFFBD]2.0.CO;2]. DOI 10.1671/0272-4634(2006)26[897:COFFBD]2.0.CO;2.}
- ↑ Gishlick, A.D. (2001). «The function of the manus and forelimb of Deinonychus antirrhopus and its importance for the origin of avian flight», Gauthier, J. and Gall, L.F. (ed.). New Perspectives on the Origin and Early Evolution of Birds. New Haven: Yale Peabody Museum, pp. 301–318.
- ↑ Parsons, W.; Parsons, K. (2006). «Morphology And Size Of An Adult Specimen Of Deinonychus antirrhopus, (Saurischia, Theropoda)» Journal of Vertebrate Paleontology. Vol. 26. n.º 3 sup.. pp. 109A.
- ↑ Kool, R. (1981). "The walking speed of dinosaurs from the Peace River Canyon, British Columbia, Canada." Canadian Journal of Earth Science, 18: 823-825.
Véase también
- Anexo: Géneros válidos de dinosaurios
Enlaces externos
 Portal:Dinosaurios Contenido relacionado con Dinosaurios.
Portal:Dinosaurios Contenido relacionado con Dinosaurios.
 Wikimedia Commons alberga contenido multimedia sobre Deinonychus.
Wikimedia Commons alberga contenido multimedia sobre Deinonychus.
 Wikiespecies tiene un artículo sobre Deinonychus.
Wikiespecies tiene un artículo sobre Deinonychus.- Deinonychus en Dinodata (en inglés, se requiere registrarse en forma gratuita)
- Deinonychus dentro de Dromaeosauridae en The Thescelsosaurus (en inglés)
- Significado y pronunciación del nombre (en inglés)
Categorías: Fósiles | Dromaeosauridae | Dinosaurios del Cretácico | Dinosaurios norteamericanos










Wikimedia foundation. 2010.