- Daspletosaurus
-
Daspletosaurus
?Daspletosaurus
Rango fósil: Cretácico superior
Estado de conservación Fósil Clasificación científica Reino: Animalia Filo: Chordata Clase: Sauropsida Superorden: Dinosauria Orden: Saurischia Suborden: Theropoda Familia: Tyrannosauridae Subfamilia: Tyrannosaurinae Género: Daspletosaurus
Russell, 1970Especie: D. torosus Nombre binomial Daspletosaurus torosus
Russell, 1970Daspletosaurus (gr. "reptil pavoroso") es un género representado por una única especie de dinosaurio terópodo tiranosáurido, que vivió a finales del período Cretácico, hace aproximadamente entre 76 y 74 millones de años, en el Campaniano, en lo que hoy es Norteamérica. Los fósiles de la única especie nombrada, D. torosus, fueron encontrados en Alberta, aunque otras especies posibles de Alberta, Montana y Nuevo México aguardan su descripción. Incluyendo estas especies sin describir, Daspletosaurus es el género de tiranosáuridos con más especies.
Como superdepredador, Daspletosaurus estaba en la cúspide de la cadena alimenticia, cazando probablemente a los dinosaurios grandes como el ceratópsido Centrosaurus y el hadrosáurido Hypacrosaurus. En algunas áreas, Daspletosaurus coexistió junto a otro tiranosáurido, Gorgosaurus, aunque hay cierta evidencia de diferenciación de nicho ecológico entre los dos. Mientras que los fósiles de Daspletosaurus son más raros que otros tiranosáuridos, los especímenes disponibles permiten un cierto análisis de la biología de estos animales, incluyendo su comportamiento social, dieta y historia de vida.
Contenido
Descripción
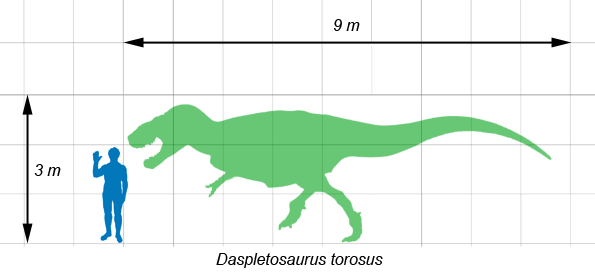
Daspletosaurus medía aproximadamente entre 8 y 9 metros de largo,[1] con un peso estimado de 2,5 toneladas.[1] [2] [3] Pero se ha estimado un rango de peso que va desde las 1,8[4] a 3,8 toneladas.[5] Una de sus principales características era su enorme cabeza, construida muy fuerte, de un metro de largo. Esta estaba preparada para absorber el impacto del golpe sobre la presa cuando atacara. Los huesos nasasales de la superficie superior del morro estaban soldados para darle mayor sólides al conjunto. Igualmente para mantener un peso bajo de la cabeza, esta poseía grandes fenestraciones. La cabeza estaba sostenida por un cuello en forma de S, que le brindaba flexibilidad y fuerza[6] . La boca estaba coronada de 6 docenas de dientes muy largos con una sección de forma oval, menos los dientes desde el premaxilar hacia el final que poseían una sección den forma de D, mostrando la heterodontia, típica de los tiranosáuridos. Las características únicas del cráneo incluyeron la superficie externa rugosa del maxilar (hueso de la mandíbula superior) y las crestas pronunciadas alrededor de los ojos en los huesos lacrimales, postorbitales y yugales. La órbita ocular era un óvalo alto, de alguna manera entre la forma circular del Gorgosaurus y la forma de cerradura del Tyrannosaurus.[7] [8] [6] .
 Primer plano del cráneo de D. torosus.
Primer plano del cráneo de D. torosus.
Daspletosaurus esta estrechamente vinculado al mucho más grande y más reciente Tyrannosaurus col el que compartió la misma forma del cuerpo. Como la mayoría de los tiranosáuridos conocidos, era un depredador bípedo de más de una tonelada equipado de docenas de dientes grandes y agudos. Daspletosaurus tenía los pequeños miembros anteriores típicos reducidos de los tiranosáuridos, aunque fueran proporcionalmente más largos que en otros géneros. Como todos los terópodos, tenia una larga cola, y como poseía largas patas traseras que terminaron en pies de cuatro dedos donde solo tres tocabán el suelo con la punta y el primer dígito (hallux) no entraba en contacto con la tierra. Los pequeños brazos solo tenían dos dedos, para quitarle peso a la parte anterior del cuerpo. Sus patas terminaban en tres fuertes dedos y la cola le servia de contrapeso a su enorme cabeza y al torso, colocando su centro de gravedad sobre las caderas.[1] [6]
Historia
El espécimen tipo de Daspletosaurus torosus (CMN 8506) es un esqueleto parcial que incluye el cráneo, el hombro, un brazo, pelvis, un fémur y toda las vértebras del cuello, torso y cadera, así como las primeras once vértebras de la cola. El holotipo fue encontrado en Alberta, Canadá por Charles Mortram Sternberg, en 1921 y descrito bajo el nombre de Gorgosaurus. No fue hasta 1970 que Dale Russell realizó una descripción completa, creando al género Daspletosaurus, del griego; δασπλητo-/daspleto- ("espantoso") y σαυρος/sauros ("lagarto").[9] Designando a la especie tipo como D. torosus, del latín por musculoso. Se conocen unos 6 especímenes en total y aparte del tipo solo se encuentra bien preservcado un esqueleto encontrado en 2001. Los deo mejores especímenes fueron recobrados de la Formación Oldman parte del Grupo Río Judith de Alberta. Un especimen de la más joven Formación Cañón Herradura en Alberta se lo ha reasignado a Albertosaurus sarcophagus.[8] La Formación Oldman fue depositadas durante mediados del Campaniano del Cretácico superior, aproximadamente entre 77 a 76 millones de años atras.[10]
Especies sin nombrar
Dos o tres especies adicionales de han asignado al género Daspletosaurus durante el correr de los años, aunque a la fecha de 2007 ninguna de estas especies habían recibido una descripción apropiada o nombre científico. Mientras tanto, todos se asignan a Daspletosaurus sp. aunque esto no implique que todos son las mismas especies.[8] [6]
Junto con holotipo, Russell señaló un espécimen recogido por Barnum Brown en 1913 como paratipo de D. torosus. Este espécimen (AMNH 5438) consiste en las partes de la pata trasera, pelvis y de algo de vértebras asociadas. Fue descubierto en la parte superior de la Formación Oldman en Alberta.[8] Esta sección superior desde entonces se ha renombrado como Formación Parque del Dinosaurio, que se data durante el Campaniano medio, entre 76 a 74 millones de años.[10] En 1914, Brown recogió un esqueleto y un cráneo casi completos; cuarenta años más tarde el Museo Americano de Historia Natural vendido este espécimen a Museo Field de Historia Natural de Chicago. Fue montada en para su exhibición en Chicago y se etiquetó como Albertosaurus libratus durante muchos años, pero después de que varias características del cráneo fueran encontradas al ser modelado en yeso, incluyendo la mayor parte de los dientes, el espécimen (FMNH PR308) fue reasignado a Daspletosaurus.[7] Un total de ocho especímenes se han recogido de la Formación Parque del Dinosaurio, la mayor parte de ellos dentro de los límites del Parque Provincial de los Dinosaurios. Phil Currie cree que los especímenes del Parque del Dinosaurio representan una nueva especie de Daspletosaurus , diferenciado por ciertas características del cráneo. Los dibujos de esta nueva especie se han publicado, pero todavía aguarda una descripción completa y nombre.[8]
Un nuevo espécimen del tiranosáurido (OMNH 10131), incluyendo fragmentos del cráneo, costillas y las partes de una pata, fue reportado de Nuevo México en 1990 y asignado al género ahora inválido Aublysodon.[11] Muchos autores posteriores han reasignado este espécimen, junto con algunos otros de Nueva México, a otra especie más sin nombre de Daspletosaurus.[8] [6] [12] Sin embargo, la investigación inédita reciente ha sugerido que esta especie, del Miembro Hunter Wash de la Formación Kirtland, es realmente un tiranosáurido temprano, quizás relacionado con Appalachiosaurus.[13] There is currently disagreement over the age of the Kirtland Formation, with some workers claiming a late Campanian age,[14] mientras que otros sugieren una edad más joven en el temprano Maastrichtiano.[15]
En 1992, Jack Horner y colegas publicaron un informe extremadamente preliminar de un tiranosáurido de las partes superiores del Campaniano en la Formación Dos Medicinas en Montana, que fue interpretado como especie transitoria entre Daspletosaurus y el posterior Tyrannosaurus. [16] Otro esqueleto parcial fue comunicado de la parte superior de Dos Medicinas en 2001, preservando los restos de un hadrosáurido juvenil en su cavidad abdominal. Este espécimen fue asignado a Daspletosaurus pero no tiene una especie en particular.[17] Los restos por lo menos de tres Daspletosaurus más también se han descrito en una cama de huesos de Dos Medicinas.[18] Estos especímenes no se han descrito detalladamente, pero Currie cree que todo el material de Dos Medicinas representa una tercera especie todavía sin nombrar de Daspletosaurus[8]
Sistemática
Daspletosaurus es colocado en la subfamilia Tyrannosaurinae dentro de la famila Tyrannosauridae, junto con el Tarbosaurus, Tyrannosaurus y posiblemente el Alioramus. Los animales de esta subfamilia están más estrechamente relacionados con Tyrannosaurus que con Albertosaurus y se conocen por su cráneos con estructura robusta proporcionalmente más grandes y de fémur más largo que los miembros de la otra subfamilia Albertosaurinae.[6] [19]
Carr et al. 2005[20] Tyrannosauridae void Albertosaurinae
Tyrannosaurinae void Daspletosaurus
void - Note: Carr et al. usa el nombre Tyrannosaurus bataar
Daspletosaurus es usualmente considerado muy cercano a Tyrannosaurus rex, o su ancestro directo por anagénesis.[16] Gregory Paul a reasignado a D. torosus al género Tyrannosaurus, creando la nueva combinación Tyrannosaurus torosus,[2] pero no ha sido mayormente aceptado.[7] [6] La mayoria de los investigadores consideran que Tarbosaurus y Tyrannosaurus deben ser considerados taxones hermanos o aun un mismo género, con Daspletosaurus como un relativo basal.[6] [20] Por otro lado, Phil Currie y colegass encuentran a Daspletosaurus más estrechamente emparentado a Tarbosaurus y a otro tiranosáurido asiático Alioramus que al norteamenricano Tyrannosaurus.[19] La sitemática de Daspletosaurus podria clarificarce una vez que se hayan descrito todas las especies.
Currie et al. 2003[19] Tyrannosauridae void Albertosaurinae
Tyrannosaurinae void void Daspletosaurus
void void Paleobiología
Coexistencia junto a Gorgosaurus
Al finales del Campaniano de Norteamérica, Daspletosaurus fue contemporaneo con el tiranosáurido albertosaurino Gorgosaurus. Éste es uno de los pocos ejemplos de dos géneros de tiranosáuridos que coexisten. En los depredadores modernos cuando dos grupos similares ocupan la misma región son separados en diversos nichos ecológicos por diferencias anatómicas, de comportamiento o geográficas que limitan la competición.[21] Varios estudios han intentado explicar la diferenciación de nichos entre Daspletosaurus y Gorgosaurus.
 Cráneo de Gorgosaurus libratus (AMNH 5434)
Cráneo de Gorgosaurus libratus (AMNH 5434) Cráneo de Daspletosaurus, Museo Tyrrell.
Cráneo de Daspletosaurus, Museo Tyrrell.Dale Russell propuso que el más ligeramente construido y más común; Gorgosaurus puede haber cazado hadrosáuridos, más abundantes en ese tiempo, mientras que el más robusto y menos más común Daspletosaurus puede haberse especializado en los ceratópsidos menos frecuentes pero mejor defendidos, que pudieron haber sido más difíciles de cazar.[1] Sin embargo, a un especímen Daspletosaurus (OTM 200) de la Formación Dos Medicinas preserva los restos digeridos de un hadrosáurido juvenil en su región intestinal.[17] Los morros más altos y anchos de los tiranosaurinos como el Daspletosaurus son mecánicamente más fuertes que los hocicos más bajos de albertosaurinos como Gorgosaurus , aunque las fuerzas de los dientes sean similares entre los dos grupos. Esto puede indicar una diferencia en mecánicos de alimentación o dieta.[22]
Otros autores han sugerido que la competición fue limitada por la separación geográfica. A diferencia de algunos otros grupos de dinosaurios, no parece haber una correlación con la distancia del mar. Ninguno, ni Daspletosaurus ni Gorgosaurus eran más comunes en terrenos más elevados o más llanos que el otro. [21] Sin embargo, mientras que hay una cierta diferenciación en la distribución, Gorgosaurus parece ser más común en las latitudes norteñas, con las especies de Daspletosaurus más abundante al sur. El mismo patrón se considera en otros grupos de dinosaurios. Los ceratopsianos chasmosaurinos y los hadrosáuridoss del hadrosaurinos son también mas comunes en la Formación Dos Medicinas y en Norteamérica al sudoeste durante el Campaniano. Thomas Holtz ha sugerido que este patrón indica preferencias ecológicas compartidas entre los tiranosaurinos, los chasmosaurinos y los hadrosaurinos. Holtz observa que, a finales del Mastrichtiano, la etapa posterior, los tiranosaurinos como el Tyrannosaurus rex , los hadrosaurinos y los chasmosaurinos como Triceratops se habían extendido por Norteamérica occidental, mientras que los albertosaurinos y los centrosaurinos se habían extinguido, y los lambeosaurinos eran muy raros. [6]
Comportamiento social
Un especímen juvenil de la especie de Daspletosaurus del Parque de los Dinosaurios (TMP 94.143.1) muestra marcas de mordeduras en la cara que fueron infligidas por otro tiranosáurido. Las marcas de la mordedura estan curadas, indicando que el animal sobrevivió la mordedura. Un ejemplar del mismo sito completamente crecido Daspletosaurus (TMP 85.62.1) también exhibe las marcas de mordeduras de tiranosáurido, demostrando que los ataques a la cara no fueron limitados a animales más jóvenes. Mientras que es posible que las mordeduras sean atribuibles a otra especie, la agresión intraespecífica, incluyendo mordeduras faciales, es muy común entre depredadores. Las mordeduras faciales se consideran en otros tiranosáuridos como Gorgosaurus y Tyrannosaurus , así como en el otros géneros de terópodos como Sinraptor y Saurornitholestes. Darren Tanke y Phil Currie presumen que las mordeduras son debido a competición intraespecífica por el territorio o recursos, o para la dominación dentro de un grupo social.[23]
Evidencia de que Daspletosaurus haya vivido en grupos sociales viene de la cama de huesos encontrada en la Formación Dos Medicinas de Montana. La cama de huesos incluye los restos de tres Daspletosaurus , incluyendo un adulto grande, un joven pequeño, y otro individuo del tamaño intermedio. Por lo menos cinco hadrosáuridos se preservan en la misma locación. La evidencia geológica indica que los restos no fueron reunidos por corrientes de agua pero todos los animales fueron enterrados simultáneamente en el mismo lugar. Los restos del hadrosáuridos están dispersos y tienen numerosas marcas de los dientes un tiranosáurido, indicando que el Daspletosaurus se alimentaban en los hadrosáuridos a la hora de muerte. La causa de la muerte es desconocida. Currie especula que los daspletosaurios formaron un grupo de caza, aunque esto no se pueda asegurar con certeza.[18] Otros científicos son escépticos de la evidencia de grupos sociales en Daspletosaurus y otros terópodos grandes.[24] Brian Roach y Daniel Brinkman han sugerido que la interacción entre daspletosaurios se asemejaría mas a lo que ocurre entre dragones de Komodo, donde individuos se reuen para alimentarse de una carcasa pero sin colaborar entre ellos, frecuentemente atacados entre ellos y llegando al canibalismo, durante el proceso de alimentación.[25]
Historia de vida
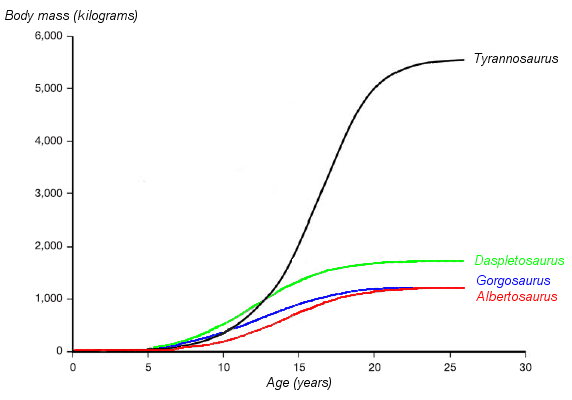
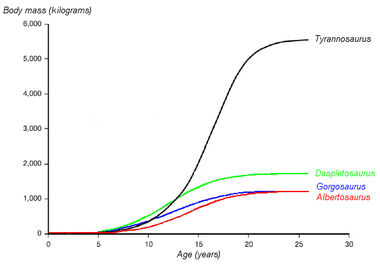 Un gráfico sobre la hipotética curva de crecimiento (masa corporal versus edad) de cuatro tiranosáuridos. Daspletosaurus es mostrado en verde. Basado en Erickson et al. 2004.
Un gráfico sobre la hipotética curva de crecimiento (masa corporal versus edad) de cuatro tiranosáuridos. Daspletosaurus es mostrado en verde. Basado en Erickson et al. 2004.El paleontólogo Gregory Erickson y colegas han estudiado la curva del crecimiento y de vida de los tiranosáuridos. El análisis de la histología de los huesos puede determinar la edad de un espécimen en el momento de su muerte. Las tasas de crecimiento pueden ser examinadas cuando la edad de varios individuos se compara contra su tamaño en un gráfico. Erickson ha demostrado que después de un tiempo largo como jóvenes, los tranosáuridos experimentaron enormes arranques de crecimiento por cerca de cuatro años a la mitad del camino de sus vidas. Después de la fase de crecimiento rápido terminara con la madurez sexual, el crecimiento retrasaba considerablemente en animales adultos. Erickson examinó solamente Daspletosaurus de la Formación Parque del Dinosaurio, pero de estos especímenes muestran el mismo patrón. Comparado con los albertosaurinos, Daspletosaurus mostró una tasa de crecimiento más rápida durante el período del crecimiento rápido debido a su peso adulto más alto. La tasa de crecimiento máxima en Daspletosaurus era de 180 kilogramos por el año, basado en una estimación de un peso adulto de 1800 kilogramos. Otros autores han sugerido pesos adultos más altos para Daspletosaurus , esto cambiaría la magnitud de la tasa de crecimiento pero no del patrón total.[4]
Por tabulación del número de especímenes de cada categoría de edad, Erickson y sus colegas pudieron extraer conclusiones sobre historia de vida en una población de Albertosaurus. Su análisis demostró que mientras que los jóvenes eran raros en el expediente del fósil, los subadultos en la fase de crecimiento rápido y los adultos eran más comunes. Mientras que esto podría ser debido a la preservación o a la polarización de la muestra, Erickson propuso que la diferencia era debido a la mortalidad baja entre jóvenes de cierto tamaño, que también se ve en algunos mamíferos grandes modernos como el elefante. Esta mortalidad baja pudo haber resultado de una carencia de la depredación, puesto que los tiranosáuridos sobrepasaron en tamaño a todos los depredadores contemporáneos a los dos años de vida. Los paleontólogos no han encontrado bastante Daspletosaurus para un análisis similar, pero Erickson observa que la misma tendencia general parece aplicarse.[26]
Paleoecología
Todo los fósiles conocidos de Daspletosaurus se han encontrado en las formaciones que se fechan en el centro a la última etapa del Campaniano del Cretácico superior, entre hace 80 y 73 millones de años. Desde mediados del Cretácico, Norteamérica había sido dividida por la mitad por la vía marítima interior occidental, con mucho de Montana y Alberta debajo de la superficie. Sin embargo, el levantamiento del Montañas Rocosas en la Orogenia Laramide al oeste, que comenzó durante la época del Daspletosaurus , forzo a la vía marítima a retirarse hacia el este y sur. Los ríos fluyeron de las montañas y drenaron en la vía marítima, llevando el sedimento junto con ellas que formaron la Formación Dos Medicinas, el Grupo Río Judith, y otro formaciones sedimentarias de la región. Hace cerca de 73 millones de años, la vía marítima comenzó a avanzar hacia el oeste y hacia el norte otra vez, y la región entera fue cubierta por el mar de Bearpaw, representado en los Estados Unidos y el Canadá occidentales por la masiva pizarra de Bearpaw.[27] [28] [29]
Daspletosaurus vivío en una llanura aluvional extensa a lo largo de la orilla occidental de la vía marítima interior. Los grandes ríos regaron la tierra, inundando y cubriendo de vez en cuando la región con nuevo sedimento. Como el agua era abundante, la región podría soportar mucha de vida vegetal y animal, pero las sequías periódicas también golpeaban la región, dando por resultado mortalidad total según lo preservado en las muchas cama de hueso encontrados en Dos Medicina y sedimentos del río Judith, incluyendo el lecho de huesos del Daspletosaurus.[30] Similares condiciones existen hoy en el este de África.[31] Erupciones volcánicas en el oeste cubrían periódicamente la región con ceniza, también dando por resultado mortalidad en grande número, mientras que simultáneamente enriquecía el suelo para el crecimiento vegetal futuro. Son estas camas de ceniza las que permiten la exacta datación radiométrica. Los niveles del mar que fluctuaban también dieron lugar a una variedad de otros ambientes en diversos momentoss y a lugares dentro del Grupo Río Judith, incluyendo costa afuera y de los hábitat marinos costeros, humedales costeros, deltas y lagunas, además de los terrenos de aluvión interiores.[28] La Formación de Dos Medicinas fue depositada en tierra más altas y más lejos del mar que las otras dos formaciones.[29]
El excelente expediente fósil de vertebrados de Dos Medicina y de las rocas del río Judith resultó de una combinación de vida animal abundante, de desastres naturales periódicos, y de la deposición de granes cantidades de sedimento. Muchos tipos de peces de agua dulce y de estuarios] se han encotrados, incluyendo tiburónes, rayas, esturiones, lepisosteus y otros. El grupo del río de Judith preserva los restos de muchos anfibios acuático y reptiles, incluyendo ranas, salamandra s, tortugas, Champsosaurus y cocodrilos. Lagartos terrestres, incluyendo teíidos, escíncidos, varanidos y lagartos de cristal también se han descubierto. Grandes pterosaurios Azhdarchidae, y neornithes como Apatornis volaron por encima, mientras que Enantiornithes como Avisaurus y varias variedades de mamíferos multituberculado, marsupiales y placentarios se escurrían debajo de los pies de Daspletosaurus y otros dinosaurios.[28]
En la Formación Oldman, Daspletosaurus torosus habría podido cazar hadrosáuridos como Brachylophosaurus y Hypacrosaurus , pequeño ornitópodos como Orodromeus, los ceratopsianos como Centrosaurus, paquicefalosaurianoo, ornitomímidos, tericinosáuridos y posiblemente anquilosáuridos. Otros depredadores incluyen troodóntidos, oviraptorosaurianos, dromeosáurido como Saurornitholestes y posiblemente un albertosaurino (de género actualmente desconocido). En el Parque de los Dinosaurios y en Dos Medicinas tienen faunas comparable a Oldman, con el Parque de los Dinosaurio particularmente preservando un arsenal incomparable de dinosaurios.[28] El albertosaurino Gorgosaurus vivio junto con el Daspletosaurus en las zonas de Parque de Dinosaurio y Dos Medicinas.[21] Los tiranosáuridos jovenes pudieron haber llenado los nichos ecologicos entre los adultos y los otros terópodos, entre los cuales habia una gran diferencia de masa.[1] [6] [22] [32]
Referencias
- ↑ a b c d e Russell, Dale A. (1970). «Tyrannosaurs from the Late Cretaceous of western Canada» National Museum of Natural Sciences Publications in Paleontology. Vol. 1. pp. 1–34.
- ↑ a b Paul, Gregory S. (1988). Predatory Dinosaurs of the World. New York: Simon & Schuster, pp. 464pp. ISBN 978-0671619466.
- ↑ Christiansen, Per; & Fariña, Richard A. (2004). «Mass prediction in theropod dinosaurs» Historical Biology. Vol. 16. n.º 2-4. pp. 85–92. DOI 10.1080/08912960412331284313.
- ↑ a b Erickson, Gregory M.; Makovicky, Peter J.; Currie, Philip J.; Norell, Mark A.; Yerby, Scott A.; & Brochu, Christopher A. (2004). «Gigantism and comparative life-history parameters of tyrannosaurid dinosaurs» Nature. Vol. 430. pp. 772–775. DOI 10.1038/nature02699.
- ↑ Therrien, François; & Henderson, Donald M. (2007). «My theropod is bigger than yours... or not: estimating body size from skull length in theropods» Journal of Vertebrate Paleontology. Vol. 27. n.º 1. pp. 108–115. DOI [108:MTIBTY2.0.CO;2 10.1671/0272-4634(2007)27[108:MTIBTY]2.0.CO;2].
- ↑ a b c d e f g h i j Holtz, Thomas R. (2004). «Tyrannosauroidea», Weishampel, David B.; Dodson, Peter; & Osmólska, Halska (eds.). (ed.). The Dinosauria, Second Edition edición, Berkeley: University of California Press, pp. 111-136. ISBN 0-520-24209-2.
- ↑ a b c Carr, Thomas D. (1999). «Craniofacial ontogeny in Tyrannosauridae (Dinosauria, Coelurosauria)» Journal of Vertebrate Paleontology. Vol. 19. n.º 3. pp. 497-520.
- ↑ a b c d e f g Currie, Philip J. (2003). «Cranial anatomy of tyrannosaurids from the Late Cretaceous of Alberta» Acta Palaeontologica Polonica. Vol. 48. n.º 2. pp. 191–226.
- ↑ Liddell, Henry G.; & Scott, Robert (1980). Greek-English Lexicon, Abridged Edition edición, Oxford: Oxford University Press,. ISBN 0-19-910207-4.
- ↑ a b Eberth, David A.; & Hamblin, Anthony P. (1993). «Tectonic, stratigraphic, and sedimentologic significance of a regional discontinuity in the Upper Judith River Group (Belly River wedge) of southern Alberta, Saskatchewan, and northern Montana» Canadian Journal of Earth Sciences. Vol. 30. pp. 174–200.
- ↑ Lehman, Thomas M.; & Carpenter, Kenneth (1990). «A partial skeleton of the tyrannosaurid dinosaur Aublysodon from the Upper Cretaceous of New Mexico» Journal of Paleontology. Vol. 64. n.º 6. pp. 1026–1032.
- ↑ Carr, Thomas D.; & Williamson, Thomas E. (2000). «A review of Tyrannosauridae (Dinosauria: Coelurosauria) from New Mexico», Lucas, Spencer G.; & Heckert, Andrew B. (eds.). (ed.). Dinosaurs of New Mexico, pp. 113–146.
- ↑ Mortimer, Michael (2002-10-16). «Details on SVP 2002 Part 2». Dinosaur Mailing List. Consultado el 2007-06-18.
- ↑ Sullivan, Robert M.; & Lucas, Spencer G. (2006). «The Kirtlandian land-vertebrate "age" - faunal composition, temporal position and biostratigraphic correlation in the nonmarine Upper Cretaceous of North America», Lucas, Spencer G.; & Sullivan, Robert M. (eds.). (ed.). Late Cretaceous vertebrates from the Western Interior, pp. 7–29.
- ↑ Ryan, Michael J. (1997). «Kirtland Formation», Currie, Philip J. & Padian, Kevin (eds.). (ed.). Encyclopedia of Dinosaurs. San Diego: Academic Press, pp. 390–391. ISBN 0-12-226810-5.
- ↑ a b Horner, John R.; Varricchio, David J.; & Goodwin, Mark B. (1992). «Marine transgressions and the evolution of Cretaceous dinosaurs» Nature. Vol. 358. pp. 59–61. DOI 10.1038/358059a0.
- ↑ a b Varricchio, David J. (2001). «Gut contents from a Cretaceous tyrannosaurid: implications for theropod dinosaur digestive tracts» Journal of Paleontology. Vol. 75. n.º 2. pp. 401–406. DOI <0401:GCFACT>2.0.CO;2 10.1666/0022-3360(2001)075<0401:GCFACT>2.0.CO;2. DOI: 10.1666/0022-3360(2001)075<0401:GCFACT>2.0.CO;2
- ↑ a b Currie, Philip J.; Trexler, David; Koppelhus, Eva B.; Wicks, Kelly; & Murphy, Nate. (2005). «An unusual multi-individual tyrannosaurid bonebed in the Two Medicine Formation (Late Cretaceous, Campanian) of Montana (USA)», Carpenter, Kenneth (ed.). (ed.). The Carnivorous Dinosaurs. Bloomington: Indiana University Press, pp. 313–324. ISBN 978-0253345394.
- ↑ a b c Currie, Philip J.; Hurum, Jørn H; & Sabath, Karol. (2003). «Skull structure and evolution in tyrannosaurid phylogeny» Acta Palaeontologica Polonica. Vol. 48. n.º 2. pp. 227–234.
- ↑ a b Carr, Thomas D.; Williamson, Thomas E.; & Schwimmer, David R. (2005). «A new genus and species of tyrannosauroid from the Late Cretaceous (middle Campanian) Demopolis Formation of Alabama» Journal of Vertebrate Paleontology. Vol. 25. n.º 1. pp. 119–143. DOI [0119:ANGASO2.0.CO;2 10.1671/0272-4634(2005)025[0119:ANGASO]2.0.CO;2].
- ↑ a b c Farlow, James O.; & Pianka, Eric R. (2002). «Body size overlap, habitat partitioning and living space requirements of terrestrial vertebrate predators: implications for the paleoecology of large theropod dinosaurs» Historical Biology. Vol. 16. n.º 1. pp. 21–40. DOI 10.1080/0891296031000154687.
- ↑ a b Snively, Eric; Henderson, Donald M.; & Phillips, Doug S. (2006). «Fused and vaulted nasals of tyrannosaurid dinosaurs: implications for cranial strength and feeding mechanics» Acta Palaeontologica Polonica. Vol. 51. n.º 3. pp. 435–454.
- ↑ Tanke, Darren H.; & Currie, Philip J. (1998). «Head-biting behavior in theropod dinosaurs: paleopathological evidence» Gaia. Vol. 15. pp. 167–184. [not printed until 2000]
- ↑ Eberth, David A.; & McCrea, Richard T. (2001). «Were large theropods gregarious?» Journal of Vertebrate Paleontology. Vol. 21. n.º (Supplement to 3 - Abstracts of Papers, 61st Annual Meeting of the Society of Vertebrate Paleontology). pp. 46A.[resumen unicamente publicado]
- ↑ Roach, Brian T.; & Brinkman, Daniel L. (2007). «A reevaluation of cooperative pack hunting and gregariousness in Deinonychus antirrhopus and other nonavian theropod dinosaurs» Bulletin of the Peabody Museum of Natural History. Vol. 48. n.º 1. pp. 103–138. DOI [103:AROCPH2.0.CO;2 10.3374/0079-032X(2007)48[103:AROCPH]2.0.CO;2].
- ↑ Erickson, Gregory M.; Currie, Philip. J.; Inouye, Brian D.; & Wynn, Alice A. (2006). «Tyrannosaur life tables: an example of nonavian dinosaur population biology» Science. Vol. 313. pp. 213–217. DOI 10.1126/science.1125721. PMID 16840697.
- ↑ English, Joseph M.; & Johnston, Stephen T. (2004). «The Laramide Orogeny: what were the driving forces?» International Geology Review. Vol. 46. n.º 9. pp. 833–838. DOI 10.2747/0020-6814.46.9.833.
- ↑ a b c d Eberth, David A. (1997). «Judith River Wedge», Currie, Philip J. & Padian, Kevin (eds.). (ed.). Encyclopedia of Dinosaurs. San Diego: Academic Press, pp. 199–204. ISBN 0-12-226810-5.
- ↑ a b Rogers, Raymond R. (1997). «Two Medicine Formation», Currie, Philip J. & Padian, Kevin (eds.). (ed.). Encyclopedia of Dinosaurs. San Diego: Academic Press, pp. 199–204. ISBN 0-12-226810-5.
- ↑ Rogers, Raymond R. (1990). «Taphonomy of three dinosaur bonebeds in the Upper Cretaceous Two Medicine Formation of northwestern Montana: evidence for drought-induced mortality» Palaios. Vol. 5. n.º 5. pp. 394–413. DOI 10.2307/3514834.
- ↑ Falcon-Lang, Howard J. (2003). «Growth interruptions in silicified conifer woods from the Upper Cretaceous Two Medicine Formation, Montana, USA: implications for palaeoclimate and dinosaur palaeoecology» Palaeogeography, Palaeoclimatology, Palaeoecology. Vol. 199. pp. 299–314. DOI 10.1016/S0031-0182(03)00539-X.
- ↑ Farlow, James O. (1976). «Speculations about the diet and foraging behavior of large carnivorous dinosaurs» American Midland Naturalist. Vol. 95. n.º 1. pp. 186–191. DOI 10.2307/2424244.
Cultura popular
Ha sido visto en varias series como Dino Rey donde se activa una carta en la isla de Bali (Indonesia). Lo extraño era que le encantaba bailar y de acuerdo con la paleontología no se descubrió en Indonesia sino que en Norteamérica
Véase también
- Anexo: Géneros válidos de dinosaurios
Enlaces externos
 Portal:Dinosaurios Contenido relacionado con Dinosaurios.
Portal:Dinosaurios Contenido relacionado con Dinosaurios.
 Wikimedia Commons alberga contenido multimedia sobre Daspletosaurus.
Wikimedia Commons alberga contenido multimedia sobre Daspletosaurus.
 Wikiespecies tiene un artículo sobre Daspletosaurus.
Wikiespecies tiene un artículo sobre Daspletosaurus.- Daspletosaurus en Dinodata (en inglés, se requiere registrarse en forma gratuita)
- Daspletosaurus en duiops
- Daspletosaurus dentro de Tyrannosauroidea en The Thescelsosaurus (en inglés)
- Significado y pronunciación del nombre (en inglés)
- Daspletosaurus en The Theropod Database (en inglés)
Categorías: Fósiles | Tyrannosauroidea | Dinosaurios del Cretácico | Dinosaurios norteamericanos



Wikimedia foundation. 2010.