- Tyrannosaurus
-
 Tyrannosaurus
TyrannosaurusRango temporal: 67 Ma-65,5 Ma Maastrichtiano (Cretácico Superior) 
Clasificación científica Reino: Animalia Filo: Chordata Clase: Archosauria Superorden: Dinosauria Orden: Saurischia Suborden: Theropoda Infraorden: Coelurosauria Superfamilia: Tyrannosauroidea Familia: Tyrannosauridae Género: Tyrannosaurus
Osborn, 1905Especie: T. rex Nombre binomial Tyrannosaurus rex
Osborn, 1905Sinonimia Manospondylus
Cope, 1892
Dynamosaurus
Osborn, 1905
Stygivenator
Olshevsky, 1995
Dinotyrannus
Olshevsky, 1995Tyrannosaurus (del gr. "lagarto tirano") es un género representado por una única especie de dinosaurio terópodo tiranosáurido, que vivió a finales del período Cretácico, hace aproximadamente 67 y 65 millones de años, en el Maastrichtiano, en lo que es hoy Norteamérica. La especie tipo y única especie válida Tyrannosaurus rex donde rex significa rey en latín, comúnmente abreviado como T. rex, es una figura común en la cultura popular. Vivió a través de lo que es ahora Norteamérica occidental, con una distribución mucho más amplia que otros tiranosáuridos. Es uno de los últimos dinosaurios no avianos en existir antes de la extinción masiva del Cretácico-Terciario.
Como otros tiranosáuridos, Tyrannosaurus fue un carnívoro bípedo con un masivo cráneo balanceado por una larga y pesada cola. En relación con sus largos y poderosos miembros traseros, los miembros superiores del Tyrannosaurus eran pequeños, pero inusualmente poderosos para su tamaño, y terminaban en dos dedos con garras. Aunque otros terópodos rivalizan o superan con Tyrannosaurus rex en tamaño, era el tiranosáurido más grande conocido y uno de los mayores depredadores conocidos de la tierra, midiendo hasta 12,8 metros de largo, y 4 metros de alto a las caderas, y con pesos estimados entre 6 a 10 toneladas. Por mucho fue el más grande carnívoro en su ambiente, Tyrannosaurus rex debió haber sido el superpredador, cazando hadrosáuridos y ceratópsidos, aunque algunos expertos han sugerido que era principalmente carroñero. El debate de si Tyrannosaurus era un depredador dominante o un carroñero es uno de los más largos en la paleontología.
Con más de 30 especímenes de Tyrannosaurus rex identificados, algunos de los cuales son esqueletos casi completos. Tejido conjuntivo y proteínas se ha informado en por lo menos uno de estos especímenes. La abundancia de material fósil ha permitido la investigación significativa en muchos aspectos de su biología, incluyendo historia de vida y biomecánica. Los hábitos de alimentación, fisiología y velocidad potencial de Tyrannosaurus rex son algunos temas de discusión. Su taxonomía es también polémica, con algunos científicos que consideran a Tarbosaurus bataar de Asia como una segunda especie de Tyrannosaurus y otros que mantienen a Tarbosaurus como género separado. Varios otros géneros de tiranosaúridos norteamericanos también han sido sinonimizados con Tyrannosaurus
Contenido
Descripción
Tyrannosaurus podía medir aproximadamente 13 metros de longitud y llegaba hasta los 4 metros de altura, con un peso estimado de entre 6 y 10 toneladas.[1] [2] El tiranosaurio poseía un gran cráneo de 1,60 m provisto de fenestras oculares y nasales. Su cráneo presenta un gran número de huesos fusionados, supliendo la movilidad por una estructura más maciza, cosa inusual en los terópodos, que por lo general tenían huesos ligeros. El cuello era grueso, musculoso y corto. Se dice que el Tyrannosaurus rex era el animal más feroz y fuerte que ha existido en toda la historia.
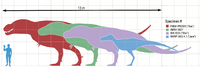 Varios especímenes de Tyrannosaurus rex en comparación con un humano.
Varios especímenes de Tyrannosaurus rex en comparación con un humano.
Tyrannosaurus rex fue uno de los carnívoros más grandes sobre la tierra, de todos los tiempos; el espécimen completo más grande, FMNH PR2081 (apodado "Sue"), midió 12,8 metros de largo, y 4 de alto a las caderas.[3] El estimado de la masa total ha variado a lo largo de los años desde las 7,2 toneladas como máximo,[4] a 4,5 como mínimo,[5] [6] con los últimos estimativos entre 5,4 y 6,8 toneladas,[7] [8] [9] [10] , y algunas estimaciones llegan hasta las 9 toneladas para los mayores especímenes conocidos de Tyrannosaurus.[11] Aunque Tyrannosaurus rex era más largo que el bien conocido terópodo del Jurásico Allosaurus y el Carcharodontosaurus africano, es ligeramente más pequeño que otros terópodos del Cretácico como Spinosaurus y Giganotosaurus.[12] [13]
El cuello de Tyrannosaurus rex forma una curva natural con forma de S como en otros terópodos, pero era corto y muscular para soportar su masiva cabeza. Los miembros superiores solo tenían dos dedos con garras,[14] junto con un pequeño metacarpiano adicional representando el remanente de un tercer dígito.[15] En cambio los miembros traseros estaban entre los más largo en proporción con tamaño de cuerpo de cualquier terópodo. La cola era pesada y larga, formada por más de cuarenta vértebras, para balancear los masivos torso y cabeza . Para compensar el inmenso tamaño del animal, muchos huesos a través del esqueleto eran huecos, reduciendo su peso sin la pérdida significativa de fuerza.[14]
 Perfil del cráneo de Tyrannosaurus (AMNH 5027)
Perfil del cráneo de Tyrannosaurus (AMNH 5027)El más grande cráneo conocido de Tyrannosaurus rex media más de 1,5 metros de largo.[16] Grandes fenestras (aberturas) reducen el peso y proporcionan lugares para la inserción muscular, como se ve en todos los terópodos carnívoros. Pero en otros aspectos el cráneo de Tyrannosaurus era perceptiblemente diferente de los terópodos no tiranosáuridos grandes. Era extremadamente ancho en la parte posterior pero tenía un hocico estrecho, permitiendo una inusualmente buena visión binocular.[17] [18] Los huesos del cráneo eran masivos y los nasales y algunos otros huesos estaban fundidos, no permitiendo movimiento entre ellos, aunque muchos pneumatizados (contenido un "panal de abejas" de espacios aéreos minúsculos) cuál puede haber hecho los huesos más flexibles además de aligerarlos. Éstos y otras características de consolidación del cráneo son parte de la tendencia de los tiranosáuridos hacia una mordedura cada vez más grande, que sobrepasó fácilmente a todos los no tiranosáuridos.[19] [20] [21] El extremo de la quijada superior era en forma de U, a mayoría de los carnívoros del no tiranosáuridos tenían quijadas superiores de forma de V, que aumentaba la cantidad de tejido y hueso que un tiranosaurio podría arrancar con una mordedura, aunque también aumentara las tensiones en los dientes delanteros.
 Dos dientes de la mandíbula inferior del ejemplar MOR 1125, “B-rex”, mostrando la variación en tamaño de los dientes en un mismo individuo
Dos dientes de la mandíbula inferior del ejemplar MOR 1125, “B-rex”, mostrando la variación en tamaño de los dientes en un mismo individuoLos dientes de Tyrannosaurus rex muestran una marcada heterodoncia (diferentes en formas).[22] [23] Los dientes premaxilares en el frente de la mandíbula superior están estrechamente empaquetados, con una sección de corte en forma de D, tenían refuerzo de cantos en la superficie posterior, eran incisiformes (sus extremidades tenían láminas como cinceles) y curvado hacia atrás. La sección de corte en forma de D, los bordes reforzados y la curvatura hacia atrás reducían el riesgo que los dientes se rompieran cuando el Tyrannosaurus mordiera y desgarrara. El resto de los dientes eran robustos, como "bananas letales" similares a dagas; espaciados más extensamente y también tenían refuerzo en los bordes.[24] Los de las quijadas superior eran más grandes que los de la parte trasera de la quijada inferior. El más grande encontrado hasta ahora se estima que medía 30 centímetros de largo incluyendo la raíz cuando el animal estaba vivo, haciéndole el diente más grande de cualquier dinosaurio carnívoro.[3]
Historia
Sus restos fósiles son escasos. Hasta 2006 han sido hallados 30 especímenes,[25] incluyendo sólo tres cráneos completos. Los primeros especímenes encontrados tuvieron un papel importante en la Guerra de los Huesos. Cabe destacar que Tyrannosaurus rex es el dinosaurio carnívoro mejor conocido en la cultura popular humana.
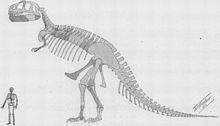 Restauración del esqueleto por William D. Matthew de 1905, una de las primeras reconstrucciones de Tyrannosaurus rex publicadas.[26]
Restauración del esqueleto por William D. Matthew de 1905, una de las primeras reconstrucciones de Tyrannosaurus rex publicadas.[26]Henry Fairfield Osborn, presidente del Museo Americano de Historia Natural, nombró a Tyrannosaurus rex en 1905. El nombre genérico proviene del las palabras en griego τυραννος (tyrannos, que significa "tirano") y σαυρος (sauros, por "lagarto"). Osborn uso la palabra latina rex, que se traduce como "rey", para el nombre binomial. El nombre binomial completa se traduce como "El rey de los lagartos tiranos", poniendo énfasis en el tamaño del dinosaurio, que dominaría sobre todos los otros animales de su tiempo.
Primeros descubrimiento
La localización de todos los descubrimientos está restringida a Norteamérica. Dientes que hoy son documentados como Tyrannosaurus rex fueron encontrados en 1874 por A. Lakes cerca de Golden, Colorado. A principios de las década de 1890, J. B. Hatcher recolecto elementos post craneales en el este de Wyoming. Estos fósiles se consideraron en un principio, perteneciente a un a especie gigante de Ornithomimus (O. grandis) pero ahora se lo considera un ejemplar de Tyrannosaurus rex. El primer espécimen, una vértebra parcial, fue descubierta por Edward Drinker Cope en 1892 y se describió como Manospondylus gigas. Fue atribuida al Tyrannosaurus rex en 1912 por Henry Fairfield Osborn.[27] Barnum Brown, el conservador auxiliar del Museo Americano de Historia Natural, encontró el segundo esqueleto del T. rex en Wyoming en 1900. Este espécimen fue originalmente nombrado Dynamosaurus imperiosus en el mismo documento en el que el Tyrannosaurus rex fue descrito.[28] Si no hubiera sido por el orden de las páginas, Dynamosaurus se hubiera convertido en el nombre oficial. El material original del "Dynamosaurus" reside en las colecciones del Museo de Historia Natural, Londres.[29]
En total, Barnum Brown encontró cinco esqueletos parciales del T. rex. Brown recolectó su segundo tiranosaurio en 1902 y 1905 en la Formación de Hell Creek, Montana. Este fue el holotipo que se usó para describir al Tyrannosaurus rex por Osborn en 1905. En 1941 se lo vendió al Museo Carnegie de Historia Natural en Pittsburgh, Pensilvania. El cuarto y más grande hallazgo de Brown, también descubierto en la Formación de Hell Creek, se exhibe en el Museo Americano de Historia Natural en Nueva York.[30]
Manospondylus
 Cráneo tipo de Tyrannosaurus rex, del Museo Carnegie de Historia Natural. Esta reconstruido muy pesado, en forma incorrecta tomando como modelo uno de Allosaurus ya que se encontraba desarmado.
Cráneo tipo de Tyrannosaurus rex, del Museo Carnegie de Historia Natural. Esta reconstruido muy pesado, en forma incorrecta tomando como modelo uno de Allosaurus ya que se encontraba desarmado.Los primeros restos fósiles atribuibles a Tyrannosaurus rex consisten en dos vértebras parciales, una de las cuales se encuentra perdida, encontradas por Edward Drinker Cope en 1892 y descrita como Manospondylus gigas. Osborn reconoció/ las similitudes entre M. gigas y T. rex temprano, en 1917 pero, debido a la pobre naturaleza de los restos vertebrales de Manospondylus, no pudo sinonimizarlos.[31]
En junio de 2000, el instituto Black Hills localizo la locación del tipo de M. gigas en Dakota del Sur y desenterraron nuevos huesos de Tyrannosaurus del lugar. Los investigadores consideraron que se trataba del mismo individuo y eran idénticos a los de Tyrannosaurus rex. De acuerdo a las reglas de Código Internacional de Nomenclatura Zoológica (ICZN), el sistema que gobierna los nombres científicos de los animales, Manospondylus gigas tendría prioridad sobre Tyrannosaurus rex, debido a que fue utilizado primero. Sin embargo en la cuarta edición de la ICZN, que empezó a tener efecto el 1 de enero de 2000, establece que " el uso que prevalece debe ser mantenido " cuando "el sinónimo mayor u homónimo no haya sido usado como valido desde 1899" y "el sinónimo menor u homónimo haya sido usado para un taxón en particular, y presumido válido para ese taxón en particular, haya sido usado para ese taxón como presunto nombre válido en al menos 25 trabajos, publicado por al menos 10 autores en inmediatamente antes de los 50 años;..."[32] Tyrannosaurus rex puede calificar como el nombre válido bajo estas condiciones y muy probablemente sería considerado un nomen protectum ("nombre protegido") bajo la ICZN si fuese desafiado, cosa que todavía no ha sido. Manospondylus gigas entonces seria a juzgado nomen oblitum ("nombre olvidado").[33]
Según las reglas de la Comisión Internacional de Nomenclatura Zoológica, el sistema que da a los animales su designación científica, usualmente a partir de raíces latinas y griegas, el nombre de Cope, creado en 1892, Manospondylus gigas, debería tener la prioridad porque su descubrimiento ocurrió primero. Sin embargo, en la 4ta edición del ICZN, que surtió efecto el 1 de enero de 2000, Capítulo 8, Artículo 35.5, declaró que cualquier tal descubrimiento hecho después de 1999 no hace que el más viejo nombre sustituya al más nuevo, prevaleciendo el nombre y que el Tyrannosaurus debe ser un nomen conservandum ("nombre conservado"). Por lo tanto, independientemente del resultado del descubrimiento, el nombre Tyrannosaurus, todavía es usado hoy por los biólogos.[34]
1940-1990
Varios otros esqueletos de Tyrannosaurus rex fueron descubiertos hasta finales de la década de 1980. El cráneo de Nanotyrannus, frecuentemente considerado un T. rex juvenil, fue recobrado de Montana en 1942. En 1966, un grupo de trabajadores del Museo Americano de Historia Natural bajo la dirección de Harley Garbani descubrió otro T. rex (LACM 23844) que presentaba un cráneo completo de un animal maduro. Cuando fue exhibido en Los Ángeles, LACM 23844 se convirtió en el más grande cráneo e exhibición de T. rex e todo el mundo. Garbani siguió descubriendo muchos esqueletos por más de una década, incluyendo LACM 23845, el holotipo de "Albertosaurus" megagracilis, muchos de los cuales son mantenidos en la colección del Museo de Paleontología de la Universidad de California en Berkeley, California. Otros cráneos y esqueletos parciales fueron descubiertos en Dakota del Sur y Alberta, Canadá a principios de la década de 1980.[35]
Antes de 1987, Tyrannosaurus rex era probablemente raro.[35] Sin embargo, en las décadas de 1980 1990 se ha presenciado el descubrimiento y la descripción de alrededor de una docena de especímenes adicionales. El primero fue Tyrannosaurus, apodado "Stan" en honor al paleontólogo amateur Stan Sacrison, se encontró en la Formación Hell Creek cerca de Buffalo, Dakota del Sur, en la primavera de 1987. Después de 30.000 horas de excavación y preparación, surgió un 65% del esqueleto completo. "Stan", BHI 3033, está actualmente en exhibición en la exposición del Museo Black Hills de Historia Natural en Hill City, Dakota del Sur, después de una extensa gira mundial y las reproducciones vendidas por el instituto Black Hills también se encuentran en galerías de museo por todo el mundo. Este Tyrannosaurus fue encontrado también con muchas patologías en sus huesos, incluyendo costillas y un cuello roto que luego se sanaron y un espectacular agujero en la parte trasera de su cabeza, con el tamaño de un diente de Tyrannosaurus.[36]
 Sue el Tyrannosaurus del Field Museum, Chicago.
Sue el Tyrannosaurus del Field Museum, Chicago.Cristóbal Veloso, descubrió el más completo (más del 90%) y más grande esqueleto fósil del T. rex conocido hasta ahora, en la Formación de Hell Creek cerca de Faith, Dakota del Sur, el 12 de agosto de 1990. Sobre la propiedad de ese espécimen de T. rex, ahora llamado Sue en honor a su descubridora, se entabló una enconada batalla legal. En 1997 esta se fijó a favor de Maurice Williams, dueño original del territorio, y la colección fósil se vendió en subasta por $7,6 millones de dólares. Actualmente el esqueleto se ha vuelto a montar y se exhibe en el Museo Field de Historia Natural. Basándose en los huesos fosilizados de Sue, el espécimen alcanzó su tamaño completo a los 19 años de edad y murió 9 años después, viviendo en total 28 años de edad.[25] Los investigadores informan el descubrimiento de dos fósiles de T. rex, uno sub-adulto y otro juvenil (ambos fueron desenterrados en la misma cantera en la que se descubrió Sue); lo cual da evidencia a la posibilidad de que el T. rex corría y cazaba en manadas u otros grupos.Las primeras especulaciones de que "Sue" pudo haber muerto por una mordida en la parte posterior de la cabeza no ha podido ser confirmada. Muchos estudios posteriores han mostrado muchas patologías, pero no se han encontrado marcas de mordidas.[37] El daño en la parte posterior del cráneo pudo haber sido causado por aplastamiento post-mortem. Especulaciones reciente indican que "Sue" puede haber muerto de hambre después de contraer una infección parásitaria por comer la carne putrefacta. La parasitosis resultante habría causado la inflamación en la garganta, impidiendo en última instancia que "Sue" pueda seguir tragando más el alimento. Esta hipótesis es apoyada por los agujeros finos y lisos en su cráneo que son similares a los causados en los pájaros modernos que contraen el mismo parásito.[38]
Últimos hallazgos
 Posible estrategia reproductora en Tyrannosaurus rex. Expuesto en el MUJA.
Posible estrategia reproductora en Tyrannosaurus rex. Expuesto en el MUJA.En el verano boreal de 2000, Jack Horner descubrió 5 especímenes de Tyrannosaurus cerca de la Reservación de Fort Peck en Montana. Uno de estos esqueletos, apodado "C. rex," fue reportado como el más largo de los Tyrannosaurus jamás encontrado.[39]
En 2001, el 50% del esqueleto de un juvenil tiranosaurio fue descubierto en la Formación de Hell Creek en Montana por un equipo de investigadores del Museo Burpee de Historia Natural de Rockford, Illinois. Apodado "Jane", el hallazgo fue inicialmente considerado el primer esqueleto conocido del pequeño tiranosáurido nanotirano pero una investigación subsecuente ha revelado que el fósil fue de un tiranosaurio juvenil. Este espécimen es el más completo y mejor preservado juvenil hasta la fecha. Jane ha sido examinada por Jack Horner, Peter Larson, Robert Bakker, Gregorio Erikson y varios otros paleontólogos renombrados, debido a la unicidad de su edad. Jane está actualmente en exposición en el Museo Burpee de Historia Natural en Rockford, Illinois.
En marzo de 2005 en la revista Science, Mary Higby Schweitzer de la Universidad Estatal de Carolina del Norte y sus colegas anunciaron la recuperación del tejido blando de la cavidad medular de un hueso de la pata fosilizada de un T. rex, que databa aproximadamente 68 millones de años. El hueso había sido roto, intencionalmente aunque con renuencia, para ser enviado y no fue conservado de la manera usual porque Schweitzer estaba deseando investigar el tejido blando. Designado como el espécimen 1125, o MOR 1125 del Museo de los Rocosas, el dinosaurio fue desenterrado previamente en la Formación de Hell Creek. Vasos sanguíneos (flexibles y bifurcados) y el tejido (de la fibrosa pero elástica matriz del hueso) fueron reconocidos. Además, se encontraron microestructuras parecidas a las células de la sangre dentro de la matriz y los vasos sanguíneos. Las estructuras son semejantes a las células y vasos sanguíneos del avestruz actual. Sin embargo, ya que un proceso desconocido y distinto al de la fosilización normal parece haber conservado el material, los investigadores son cuidadosos de no afirmar que este es el material original del dinosaurio.[40] La presencia de huesos medulares en este espécimen también es interesante.[41]
Si resulta ser el material original, cualquier proteína sobreviviente puede usarse como medio de estimar indirectamente algunos de los contenidos del ADN (ácido desoxirribonucleico) de los dinosaurios involucrados, porque cada proteína se crea típicamente por un gen específico. La ausencia de hallazgos anteriores puede ser meramente el resultado de personas que asumen que la conservación del tejido era imposible, y simplemente no lo observaron; desde el primero, dos tiranosaurios más y un hadrosaurio también han sido encontrados teniendo estas estructuras similares a tejidos. [2][3]
En un boletín de prensa del 7 de abril de 2006, la Universidad Estatal de Montana reveló que el cráneo de tiranosaurio más grande hasta ahora hallado también poseía esta característica. Descubierto en los años 1960 y recientemente reconstruido, el cráneo mide 149,8 centímetros (59 pulgadas) de largo, comparado con el cráneo de "Sue" (140,7 cm, es decir 55,4 pulgadas) hay una diferencia de 6,5%. [4] & [5]
A pesar de que se conocen numerosos esqueletos, solo un rastro de huellas ha sido bien documentado, en el Rancho Philmont Scout al noeste de Nuevo México. Fueron descubiertas en 1983 e identificadas y documentadas en 1994.[42]
Clasificación
 Modelo del T. rex apodado Stan, expuesto en el Museo de Manchester.
Modelo del T. rex apodado Stan, expuesto en el Museo de Manchester.Tyrannosaurus es, el género tipo, mundialmente aceptado, de la superfamilia Tyrannosauroidea, la familia Tyrannosauridae y la subfamilia Tyrannosaurinae. Otros miembros de la subfamilia Tyrannosaurinae, incluyen al Daspletosaurus de América del Norte y a Tarbosaurus de Asia;[43] [44] los cuales son, ocasionalmente, clasificados dentro del género del Tyrannosaurus.[45] Los tiranosáuridos fueron vistos por mucho tiempo como los descendienes de grandes terópodos anteriores como los megalosáuridos y carnosaurios, aunque actualmente se los considera como parte de los habitualmente pequeños celurosaurios.[46]
En 1955, el paleontólogo soviético Evgeny Maleev, nombró al Tyrannosaurus bataar como una nueva especie de Mongolia.[47] Para 1965, esta especie fue renombrada como Tarbosaurus bataar.[48] El Tarbosaurus, de Mongolia, es a veces colocado en el género Tyrannosaurus como T. bataar, aunque la mayor parte de investigadores de tiranosaurios, como Tom Holtz, ven suficientes diferencias entre esas dos especies como para asegurar que se trata de géneros separados.[44] mientras que otros lo consideran la especie asiática de Tyrannosaurus.[46] [49] [50] Una reciente descripción del cráneo de Tarbosaurus bataar ha mostrado que es más estrecho que el de Tyrannosaurus rex y que durante la mordida, la distribución de las tensiones en los huesos de la calavera eran muy distintas, siendo más cercana a la de Alioramus, otro tiranosáurido asiático.[51] Un reciente análisis cladistico encontró que Alioramus, y no Tyrannosaurus, es el taxón hermano de Tarbosaurus, lo que sugiere que Tarbosaurus y Tyrannosaurus deben permanecer separados.[43]
Otros fósiles de tiranosáuridos encontrados en las mismas formaciones que Tyrannosaurus rex han sido originalmente atribuidos a diferentes taxones, incluyendo a Aublysodon y Albertosaurus megagracilis,[45] que fue llamado posteriormente llamado Dinotyrannus megagracilis en 1995.[52] Sin embargo, estos fósiles son universalmente considerados un ejemplar juvenil de Tyrannosaurus rex.[53] Un pequeño pero muy completo cráneo encontrado en Montana, de 60 centímetros de largo, puede ser una excepción. Este cráneo fue originalmente clasificado como una especie de Gorgosaurus (G. lancensis) por Charles W. Gilmore en 1946,[54] pero posteriormente le fue asignado a su propio género, Nanotyrannus.[55] Las opiniones sobre la validez de N. lancensis están divididas. Muchos paleontólogos consideran al cráneo como perteneciente a un juvenil Tyrannosaurus rex.[56] Existen menores diferencias entre los dos, incluyendo un mayor número de dientes en N. lancensis, lo que ha llevado a los científicos en recomendar que ambos géneros se mantengan separados, hasta que nuevos descubrimientos ayuden a clarificar estas cuestiones.[44] [57]
Un gran número de especies inválidas de Tyrannosaurus ha sido clasificado como T. rex y Tarbosaurus bataar. La lista es la siguiente:
- T. amplus (Marsh, 1892) nomen dubium (originalmente Aublysodon) especie no válida, ahora Aublysodon amplus.
- T. bataar (Maleev, 1955) especie no válida, ahora Tarbosaurus bataar.
- T. efremovi (Maleev, 1955) (originalmente Tarbosaurus) especie no válida, ahora Tarbosaurus efremovi.
- T. gigantus (1990) ("reptil tirano gigante") nomen nudum, especie no válida, ahora Tyrannosaurus rex.
- T. imperiosus (Osborn, 1905) (originalmente Dynamosaurus) especie no válida, ahora Tyrannosaurus rex.
- T. lancensis (Gilmore, 1946) (originalmente Gorgosaurus) = Tyrannosaurus rex?
- T. lancinator (Maleev, 1955) (originalmente Gorgosaurus) especie no válida, ahora Tarbosaurus bataar.
- T. lanpingensis (Yeh, 1975) nomen dubium especie no válida, ahora Tarbosaurus lanpingensis.
- T. luanchuanensis (Dong, 1979) nomen dubium especie no válida, ahora Tarbosaurus luanchuanensis.
- T. megagracilis (Paul, 1988) (originalmente Albertosaurus) = Tyrannosaurus rex?
- T. novojilovi (Maleev, 1955) (originalmente Gorgosaurus) = Tarbosaurus bataar?
- T. stanwinstonorum (Pickering, 1995) nomen nudum especie no válida, ahora Tyrannosaurus rex.
- T. torosus (D. A. Russell, 1970) (originalmente Daspletosaurus) especie no válida, ahora Daspletosaurus torosus.
- T. turpanensis (Zhai, Zheng & Tong, 1978) especie no válida, ahora Tarbosaurus bataar.
- Refiérase a Tyrannosauridae para una lista completa de tiranosaurios avanzados, como Gorgosaurus, Albertosaurus y Alectrosaurus. Refiérase a Tyrannosauroidea para una discusión de tiranosaurios primitivos, tales como el Dilong, Guanlong y Eotyrannus.
Paleoecología
 Antigua representación del T. rex (con una postura incorrecta, ver más arriba) en su hábitat natural.
Antigua representación del T. rex (con una postura incorrecta, ver más arriba) en su hábitat natural.El tiranosaurio vivía en todo el occidente de Norteamérica, desde Alberta (Canadá), hasta Coahuila en México,[58] justo antes de que los dinosaurios se extinguieran. Normalmente el T. rex habitaba en planicies de inundación y bosques subtropicales donde acechaba a sus presas, en zonas demarcadas por ríos, lagos y bosques exuberantes llenos de cicadáceas, helechos, plantas florecidas y árboles como las coníferas, sicomoros y araucarias.
En la época del T. rex, Norteamérica presentaba un paisaje natural con elementos familiares y extraños. Las tortugas de cuero, los cocodrilos, los lucios (Esocidae), y los peces pipa (Lepisosteidae) que vivieron en esa época eran bastante similares a los que se pueden encontrar hoy. Las ranas y los lagartos varanos eran otros animales familiares. Los helechos, colas de caballo, palmas, magnolias, álamos y arbustos eran algunas de las plantas dominantes; los pastos y hierbas ya se habían desarrollado, pero no estaban aún extendidas. Las coníferas como sequoias, araucarias, pinos, y cipreses eran comunes. El T. rex probablemente vivió en muchos hábitats diferentes debido a su amplio rango, pero muchos de los yacimientos fósiles en donde normalmente se encuentran sus esqueletos parecen haber sido bosques subtropicales y húmedos. Otros habitantes del paisaje son menos familiares y carecen de semejanza con la fauna actual. Los pterosaurios gigantes, como el Quetzalcoatlus, planeaban y volaban en los cielos, con envergaduras de alas de más de 12 metros. Otros terópodos, incluyendo a los dromeosáuridos, troodóntidos y ornitomímidos, parecen haber medido menos de 4 ó 5 metros de largo. Las manadas de ceratopsianos como los tricerátopos y torosaurios, y de hadrosaurios como los hadrosaurios y edmontosaurios, vagaban por la tierra, mientras aves dentadas volaban en los bosques (Ichthyornis) y nadaban en las orillas de los mares (Hesperornis). Otros dinosaurios herbívoros fueron el armado anquilosaurios, los "cabeza-duras" paquicefalosaurios y estigimoloch y pequeños ornitópodos como Bugenasaura y tescelosaurio. Antiguos primates también pudieron haber existido junto a los dinosaurios (esta publicación está abierta a la discusión). Los mamíferos (predominantemente multituberculados y marsupiales) eran todavía pequeños, animales nocturnos que se asemejaban mucho a las ratas y musarañas de hoy, como el Ptilodus y Meniscoessus; aunque había géneros excepcionales que parecían ya un poco más grandes y desarrollados, como el Taeniolabis.
Se cree que el tiranosaurio necesitaba extensos rangos geográficos de alimentación, debido a la retirada de la Vía Marítima Interior Occidental de Norteamérica, hace 69 millones de años, lo cual incrementó el tamaño del rango de alimento.[59]
Paleobiología
Postura
Al igual que muchos dinosaurios bípedos, el Tyrannosaurus rex fue descrito históricamente como un trípode vivo, con el cuerpo a 45 grados o menos de la vertical y la cola arrastrando por el suelo, similar a un canguro. Este concepto data de 1865 cuando Joseph Leidy realizo la reconstrucción de Hadrosaurus, el primero en describir un dinosaurio en una postura bípeda.[60] Henry Fairfield Osborn, ex presidente del Museo Americano Historia Natural (AMNH) en Nueva York, creyo que la criatura podría colocarse en posición vertical y refuerza aún más la idea después de exhibir el primer esqueleto completo de Tyrannosaurus rex en 1915. Se mantuvo en esta posición vertical durante casi un siglo, hasta que fue desmantelado en 1992.[61] En 1970, los científicos se dieron cuenta de esta postura era incorrecta y no podría haber sido mantenida por un animal vivo, ya que habría dado lugar a la dislocación o debilitamiento de varias articulaciones, incluyendo las caderas y la articulación entre la cabeza y la columna vertebral.[62] El montaje inexacto ha inspirado muchas representaciones similares en películas y pinturas, como el famoso mural Rudolph Zallinger "La Edad de los Reptiles" en el Museo Peabody de la Universidad de Yale.[63] Esto sucedió hasta la década de 1990, cuando las películas como Jurassic Park presentaron una postura más exacta al público en general. Representaciones modernas en los museos, arte y cine de Tyrannosaurus Rex muestran a su cuerpo aproximadamente paralelo al suelo y la cola extendida detrás del cuerpo para balancear la cabeza.[45]
El tiranosaurio, al igual que todos los terópodos, era bípedo. Sus patas estaban dotadas de un tejido almohadillado. Este tejido también funcionaba como un resorte. Los huesos largos de las patas se fusionan entre sí para transferir las fuerzas mayores generadas por sus pisadas fuertes, por las piernas y hacia el resto del cuerpo.
Los estudios han demostrado que el tiranosaurio tenía unas patas bastante largas. Todavía existen desacuerdos sobre cuan rápido se desplazaba el tiranosaurio. Los cálculos oscilan entre una velocidad pausada de 18 km/h y una muy rápida de 72 km/h. Los científicos que consideran que el T. rex se movía rápidamente indican que sus patas eran semejantes a las de ornitomímidos tan veloces como el estrutiomimo. Un estudio reciente concluyó que el T. rex no tenía suficiente masa muscular en las piernas como para ser tan veloz, lo que hacía era caminar dando zancadas de 4 m por cada paso dándole una velocidad de 42km/h.
Algunos científicos indican que los animales pesados tienen las patas ubicadas debajo del cuerpo como pilares, con huesos grandes para soportar el peso, lo que no les permite correr. Quizá el T. rex era lento. Sólo se ha descubierto una huella en México que podría ser de un T. rex y que mide casi 1 m de longitud. Si se encontrara toda la serie de las huellas se conocería mejor que tan rápido pudo andar. Los bípedos tienen mayor riesgo de caerse, si durante un arranque se tropiezan, y no pueden acomodar sus patas debajo del cuerpo. Estudios recientes sugieren que los adultos no eran animales especialmente veloces, aunque en una persecución podían dar zancadas de más de 4 m.
Las caídas eran muy peligrosas para el T. rex porque la cabeza recorría más de 3 m en el desplome, y los brazos no podían detener su caída. Las avestruces tienen un problema similar, pero el riesgo de caída de un avestruz o de otras aves corredoras es muchísimo menor que el que habría tenido un tiranosauro incluso siendo éste depredador (si cazaba presas vivas). Si el T. rex se caía duramente podía lesionarse e incluso morir. Unos investigadores calcularon que si un T. rex de 6 toneladas corriera a una velocidad de 72 km/h y tropezara, golpearía el suelo con mucha fuerza y se deslizaría con tanta presión que no podría soportar el dolor de sus huesos y músculos rotos, lo que le causaría la muerte. Pero otro equipo de investigadores dio una propuesta más aceptable, trata de la posibilidad que el T. rex se mueva entre los 18 y 54 km/h, parecida a la velocidad máxima de un elefante africano. Si el T. rex se cayera a dicha velocidad, se lesionaría y quizás se rompería algunos huesos pero podría sobrevivir.
El cerebro del tiranosaurio era tan grande como el de un gorila y tenía aproximadamente un volumen de 1000 cm³, aunque la mayor parte del mismo estaba dedicado a su sentido del olfato, que, según se cree, pudo haber sido el más agudo de todos.[cita requerida]
Extremidades Superiores
 Brazo de tiranosaurio.
Brazo de tiranosaurio.Cuando Tyrannosaurus rex fue descubierto, no se hallaron las extremidades superiores.[64] Para completar el esqueleto original, que fue montado para ser exhibido al público, Osborn sustituyó esa parte restante por los «brazos» con tres dedos de un Allosaurus.[31] Sin embargo, en 1914, Lawrence Lambe describió un corto brazo de 2 dedos para el estrechamente relacionado Gorgosaurus.[65] Por lo que le atribuyó 2 dedos en lugar de 3, conjetura que se confirmó en 1989, con el hallazgo de brazos relativamente completos de Tyrannosaurus rex, perteneciente a MOR 555, el "Wankel rex".[30] Los restos de «Sue» también incluyen brazos completos.[14] Estos eran relativamente pequeños en comparación del cuerpo, de aproximadamente 1 metro de largo, sin embargo no eran órganos vestigiales, ya que presentaban grandes áreas para la inserción de los músculos, los que le brindaban considerable fuerza. Así lo reconoció ya en 1906 Osborn, quien especuló que los miembros anteriores pueden haber sido usados para atrapar a la pareja durante la copulación.[66] También se ha sugerido que los miembros anteriores se utilizaron para ayudar a los animales a levantarse de una posición de decúbito esternal.[62] Otra posibilidad es que los miembros anteriores detuvieran a la presa durante la lucha, mientras las enormes mandíbulas del tiranosaurio la mataban. Esta hipótesis puede ser apoyada por el análisis biomecánico.
Los huesos de las extremidades anteriores de Tyrannosaurus rex presentan muy grueso hueso cortical, que indica que fueron desarrollados para soportar cargas pesadas. El músculo bíceps braquial de un adultoTyrannosaurus rex era capaz de levantar 199 kg por sí solo, número que aumentaría al actuar con otros músculos, como el músculo braquial. El antebrazo de Tyrannosaurus también sufría una disminución en el rango del movimiento, con el hombro y codo que permitían sólo 40 y 45 grados de movimiento, respectivamente. Por el contrario, las mismos dos articulaciones en Deinonychus permitían hasta 88 y 130 grados de movimiento, respectivamente, mientras que un brazo humano puede girar 360 grados en el hombro y moverse alrededor de 165 grados a nivel del codo. El pesado cuerpo de los huesos del brazo, la fuerza extrema de los músculos, y el rango limitado de movimiento puede indicar un sistema evolucionado que se mantenga firme a pesar de las tensiones de una presa que lucha.[67]
Crecimiento
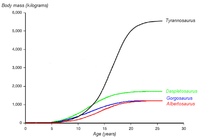 Curva de crecimiento de T.rex comparada con la de otros tiranosáuridos. Basada en Erickson et al. 2004.
Curva de crecimiento de T.rex comparada con la de otros tiranosáuridos. Basada en Erickson et al. 2004.La identificación de varios ejemplares juveniles como de tiranosaurio ha permitido a los científicos documentar los cambios ontogénicos en la especie, estimar la vida útil, y determinar con qué rapidez estos animales habrían crecido. El ejemplar más pequeño conocido, LACM 28471, el "terópodo Jordan", se estima que sólo pesaba 30 kg, mientras que el más grande, como FMNH PR2081, "Sue", lo más probable es que pesaba más de 5400 kg . El análisis histológico de los huesos de LACM 28471 mostró sólo tenía dos años cuando murió, mientras que "Sue" tenía 28 años, una edad que puediendo haber estado cerca del máximo de la especie.[7]
La histología también ha permitido calcular la edad de otros ejemplares. Las curvas de crecimiento puede desarrollarse cuando las edades de diferentes especímenes se trazan en un gráfico junto con su masa. Una curva de crecimiento para Tyrannosaurus rex tiene forma de S, con los menores que permanecen bajo 1800 kg hasta aproximadamente los 14 años de edad, cuando el tamaño del cuerpo comienza a aumentar dramáticamente. Durante esta fase de crecimiento rápido, un joven tiranosaurio ganaría un promedio de 600 kg al año durante los próximos cuatro años. A los 18 años de edad, la curva se ameseta de nuevo, lo que indica que el crecimiento se redujo drásticamente. Por ejemplo, sólo 600 kg separados los 28 años de edad, "Sue" deun ejemplar canadiense de 22 años de edad, RTMP 81.12.1.[7] Otro estudio histológico reciente realizado por cientificos diferentes corrobora estos resultados, encontrando que el rápido crecimiento comenzó a disminuir en alrededor de 16 años de edad.[68] Este repentino cambio en la tasa de crecimiento pueden indicar la madurez física, una hipótesis que es apoyada por el descubrimiento de tejido medular en el fémur de un tiranosaurio de 16 a 20 años de edad, de Montana, MOR 1125, también conocida como "B-rex". El tejido medular se encuentra sólo en las hembras durante la ovulación, lo que indica que "B-rex" estaba en la edad reproductiva.[69] Un estudio adicional indica una edad de 18 años para esta muestra.[70] Otros tiranosáuridos presentan curvas de crecimiento muy similares, aunque con menores tasas de crecimiento correspondientes a las tallas de adulto más baja.[71]
Más de la mitad de los ejemplares conocidos de tiranosaurio parece que han muerto a los seis años de alcanzar la madurez sexual, un patrón que se observa también en otros tiranosáuridos y en algunas aves grandes de larga vida y mamíferos en la actualidad. Estas especies se caracterizan por las altas tasas de mortalidad infantil, seguido de una mortalidad relativamente baja entre los jóvenes. La mortalidad aumenta de nuevo después de la madurez sexual, en parte debido a las tensiones de la reproducción. Un estudio sugiere que la escasez de fósiles de de menores Tyrannosaurus rex se debe en parte a las bajas tasas de mortalidad de menores, los animales no estaban muriendo en gran número a estas edades, por lo que no eran a menudo fosilizados. Sin embargo, esta rareza también puede ser debido a lo incompleto del registro fósil o el sesgo de los recolectores hacia fósiles ejemplares más grandes y espectaculares.[71]
 Esqueleto fósil de Tyrannosaurus rex en el Museo Nacional de Historia Natural, Washington, DC.
Esqueleto fósil de Tyrannosaurus rex en el Museo Nacional de Historia Natural, Washington, DC.El Tiranosaurus, crecía a un ritmo de 2,1 kilos diarios, lo que le permitía, en sus casi 30 años de vida, alcanzar las seis toneladas de peso, según revela un novedoso estudio realizado por la Universidad Estatal de Florida (Estados Unidos) y publicado en la revista Nature.
El estudio, dirigido por los científicos Greg Erickson y Peter Makovicky, de la Universidad de Tallahassee (Florida), demuestra que ciertos dinosaurios crecían a un ritmo extremadamente rápido, lo que les permitía alcanzar un inmenso tamaño en poco tiempo. Sus autores han desarrollado un análisis completo del crecimiento de este dinosaurio, un carnívoro voraz que vivió hace 65 millones de años, y han concluido que este animal engordaba 2,1 kilos diarios durante su adolescencia.
Con este ritmo de crecimiento, el saurio alcanzaba su madurez ósea en dos décadas y podía vivir hasta 28 años. A partir de los 14 a 18 años de edad, el tiranosaurio -al igual que sus familiares como el albertosaurio, el gorgosaurio y el daspletosaurio- adquiría alrededor del 70% de su masa adulta, y pasaba de ser un lagarto carnívoro de una tonelada a convertirse en un dinosaurio devorador de huesos de seis tonedadas de peso, con pocos rivales en la cadena alimenticia el mundo prehistórico.
Ese ritmo de crecimiento puede compararse con el de los elefantes, aunque con la diferencia de que éstos llegan a vivir 70 años, mientras que los dinosaurios tan sólo alcanzaban los 30. "Sabemos que el 'Tiranosaurus Rex' vivía rápido y moría joven", indica el doctor Erickson en Nature. Erickson y Makovicky han basado su descubrimiento en el análisis de más de 60 huesos de 20 fósiles diferentes de la familia del Tiranosaurus. Entre los especímenes examinados por estos cienfíticos se encuentra la Tiranosaurus conocida por el nombre de Sue, cuyo esqueleto -el mayor y más completo hallado hasta ahora- se conserva en un 90% en el Museo Field de Chicago.
Hasta ahora se sabía que al igual que los árboles, cuyo crecimiento queda grabado en los anillos de su tronco, los principales huesos de los dinosaurios también poseen estas estrías, lo que permite determinar su crecimiento. Sin embargo, con el paso del tiempo estos huesos desarrollaban unas cavidades que dificultaban su lectura. El nuevo estudio se ha centrado en huesos de menor grosor, como las costillas, y se ha descubierto que con los años no cambiaban, sino que se mantienen en perfecto estado y conservan las líneas de crecimiento.
Dimorfismo sexual
Con el número de ejemplares en aumento, los científicos comenzaron a analizar la variación entre los individuos y descubrieron lo que parecían ser dos tipos de conformaciones distintas, al igual que algunas especies de otros terópodos. Como uno de estas conformaciones estaba más sólidamente construida, que se denominó el "robusta" mientras que el otro se denomina "grácil". Varias diferencias morfológicas asociadas a los dos morfotipos fueron utilizadas para analizar el dimorfismo sexual en el Tyrannosaurus, siento el tipo el "robusto" por lo general considerado hembra. Por ejemplo, la pelvis de varios especímenes "robustos" parecía ser más amplia, tal vez para permitir el paso de los huevos.[72] Además, se consideró que la morfología "robusta" estaba correlacionada con un cheuron reducido en la primera vértebra de la cola, también para permitir que los huevos salen del aparato reproductor, como se había informado erróneamente de cocodrilos.[73]
En los últimos años, las pruebas sobre el dimorfismo sexual se ha debilitado. Un estudio de 2005 informó de que las afirmaciones previas del dimorfismo sexual en la anatomía del cheuron cocodrilo fue un error, arrojando dudas sobre la existencia de dimorfismo similar entre sexos de Tyrannosaurus rex.[74] Un cheuron de tamaño natural fue descubierto en la primera vértebra de la cola de "Sue", un individuo muy robusto, lo que indica que esta característica no puede ser utilizado para diferenciar los dos morfotipos de todos modos. Como los ejemplares de Tyrannosaurus rex se han encontrado de Saskatchewan a Nuevo México, por lo que las diferencias entre los individuos puede ser indicativo de la variación geográfica en lugar de dimorfismo sexual. Las diferencias también podrían ser relacionada con la edad, con los ejemplares robustos como los animales más viejos.[14]
Sólo un único ejemplar de Tyrannosaurus se ha demostrado en forma concluyente a pertenecer a un género específico. En el examen de "B-rex" se demostró la preservación de tejido blando dentro de varios huesos. Parte de este tejido ha sido identificado como tejido medular, un tejido especializado encontrado exclusivamente en las aves modernas, como fuente de calcio para la producción de la cáscara de huevo durante la ovulación. Puesto que sólo las hembras ponen huevos, el tejido medular sólo se encuentra naturalmente en las hembras, aunque los machos son capaces de producir cuando son inyectados con hormonas reproductivas femeninas como el estrógeno. Esto sugiere fuertemente que "B-rex" era una hembra, y que murió durante la ovulación.[69] La investigación reciente ha demostrado que el tejido medular no se encuentra en cocodrilos, que se cree que son los parientes vivos más cercanos de los dinosaurios, además de las aves . La presencia compartida de tejido medular en las aves y los dinosaurios terópodos es una prueba más de la estrecha relación evolutiva entre los dos.[75]
Tejidos Blandos
En el ejemplar de marzo de 2005 de la revista Science, Mary Higby Schweitzer y colegas de la Universidad Estatal de Carolina del Norte anunciaron la recuperación de tejidos blandos de la cavidad medular de un hueso de la pierna fosilizado de un Tyrannosaurus. El hueso había sido intencionalmente, aunque a regañadientes, roto por el envío y luego no conservado en las condiciones normales, específicamente porque Schweitzer tenía la esperanza de encontrar pruebas de los tejidos blandos.[76] Designado como MOR 1125, este dinosaurio fue excavado con anterioridad de la Formación Hell Creek. El flexible con bifurcación de los vasos sanguíneos matriz de tejido óseo fibrosa pero elástica fueron reconocidas. Además, microestructuras parecidas células de la sangre se encuentra dentro de la matriz y los vasos. Las estructuras se parecen a las células de sangre y vasos de avestruz. Ya sea un por proceso desconocido, distinto de la fosilización normal, u otro proceso fortuito es que se haya conservado el ejemplar, por lo que los ineestigadores tienen cuidado de no hacer ninguna aseveración sobre la preservación.[77] Si se comprueba que el material original posee cualquier proteína sobreviviente puede usarse como un medio de forma indirecta adivinar algo sobre el contenido de ADN de los dinosaurios involucrados, porque cada proteína se crea normalmente por un gen específico. La ausencia de hallazgos anteriores puede ser meramente el resultado de que se asumia que la preservación del tejido blando era imposible, por tanto, simplemente no se buscaba. Desde el primer descubrimiento, en dos tiranosaurios más y en un hadrosaurio también se han encontrado que tales estructuras.[76] La investigación sobre algunos de los tejidos involucrados ha sugerido que las aves están más cercanas en el árbol evolutivo de los tiranosaurios que otros animales modernos.[78]
En los estudios reportados en la revista Science en abril de 2007, Asara y sus colegas concluyeron que siete trazas de proteínas de colágeno se encontraron en los huesos purificados de Tyrannosaurus rex se acercan más a los reportados en los pollos, seguido de las ranas y tritones. El descubrimiento de las proteínas de una criatura de decenas de millones de años de antigüedad, junto con restos similares que el equipo encontró en un hueso de mastodonte de al menos 160.000 años de edad, voltea la visión convencional de los fósiles y paleontólogos pudiendo cambiar el foco "de la caza del hueso a la bioquímica". Hasta estos hallazgos, la mayoría de los científicos presumía que la fosilización sustituye todos los tejidos vivos con minerales inertes. El paleontólogo Hans Larsson, de la Universidad McGill en Montreal, que no formaba parte de los estudios, denominado encuentra "un hito", y sugirió que los dinosaurios podrían "entrar en el campo de la biología molecular y la paleontología en el mundo moderno."[79]
Estudios posteriores en abril de 2008 confirmaron la estrecha relación de Tyrannosaurus con las aves modernas. Investigador postdoctoral biología Chris Organ la Universidad de Harvard anunció: "Con más datos, que probablemente sería capaz de colocar el T. rex en el árbol evolutivo entre los cocodrilos y las gallinas y los avestruces". Co-autor John M. Asara agregó, "También muestran que los grupos mejor con las aves de los reptiles modernos, como los caimanes y lagartos anolis verdes."[80]
El presunto tejido blando fue puesto en duda por Kaye Thomas de la Universidad de Washington y sus colaboradores en 2008. Sostienen que lo que realmente estaba en el interior del hueso de tiranosaurio fue biopelícula pegajosa creado por las bacterias que cubría los huecos una vez ocupado por los vasos sanguíneos y las células.[81] Los investigadores encontraron que lo que previamente se había identificado como los restos de las células de la sangre, debido a la presencia de hierro, eran en realidad framboides, mineral de hierro microscópicas esferas. Encontraron esferas similares en una variedad de otros fósiles de distintas épocas, incluyendo un ammonites. En los ammonites que se encuentran las esferas en un lugar donde el hierro que contienen no podría haber tenido alguna relación con la presencia de sangre.[82] Sin embargo, Schweitzer ha criticado duramente las reclamaciones de Hayes y sostiene que ella realmente se encontraron células de la sangre, y argumenta que no hay evidencia informada que los biopelícula puedan producir ramificación, tubos huecos, como las mencionadas en su estudio.[83]
Plumas
Dada la posición filogenética de Tyrannosaurus, se ha propuesto que pudo haber tenido plumas. Pequeños celurosaurianos, el grupo de dinosaurios al que pertenece, de la Formación Yixian en Liaoning, China, han sido descubiertos presentando plumas penáceas o con el antiguo pelaje de "protoplumas", lo que sugiere la posibilidad de que los tiranosáuridos también pudieron haber tenido plumas. En 2004, la revista científica Nature, publicó un informe que describe al temprano tiranosauroide Dilong paradoxus, fuera descubierto en la misma formación y mostró pruebas de filamentos de protoplumas en la cola. Sin embargo, impresiones de la piel de tiranosaurios adultos de Alberta y Mongolia aparecen mostrando las escamas escaladas típicas de otros dinosaurios.[84] Esto sería compatible con el tamaño de la criatura, ya que, con un peso de 8 toneladas, la dispersión de calor del tiranosaurio de sangre caliente en realidad habría sido impedida por una cubierta de plumas. Es posible que el T. rex presentara plumas o protoplumas en otras regiones del cuerpo pero, tal como ocurre con el pelo de los elefantes y rinocerontes modernos, en áreas reducidas. Como un objeto que aumenta de tamaño, su capacidad para retener calor aumenta debido a su decreciente relación área-volumen. Por lo tanto, como animales de gran tamaño, en climas cálidos, un abrigo de pieles o plumas pierde su ventaja evolutiva como aislamiento térmico y puede convertirse a la vez en una situación de desventaja, ya que el aislamiento de calor el exceso no le permitiria refrescarse, ocasionando el sobrecalentamiento del animal. Las protoplumas puede haberse perdido secundariamente durante la evolución de los tiranosáuridos grandes como Tyrannosaurus, especialmente en climas cálidos del Cretácico.[85]
Termorregulación
A los tiranosaurios, como a la mayoría de los dinosaurios, por mucho tiempo se los pensó como ectotérmicos, de "sangre fría", con un metabolismo de reptil. La idea de la ectotermia de los dinosaurio fue rechazada por los científicos como Robert T. Bakker y John Ostrom en los primeros años del "Renacimiento de los Dinosaurios", a partir de la década de 1960.[86] [87] Alegando que los tiranosaurios debieron haber sido endotérmicos, de "sangre caliente", lo que implica un estilo de vida muy activo.[6] Desde entonces, varios paleontólogos han tratado de determinar la capacidad de Tyrannosaurus para regular su temperatura corporal. A través de evidencia histológica de altas tasas de crecimiento en los jóvenes Tyrannosaurus rex, comparables a los de los mamíferos y las aves, apoyando la hipótesis de un metabolismo alto. Las curvas de crecimiento indican que, como en los mamíferos y las aves, el crecimiento del Tyrannosaurus rex se limitó sobre todo a los animales inmaduros, más que el crecimiento indeterminado visto en la mayoría de otros vertebrados.[68]
Las proporciones de isótopos de oxígeno en los huesos fosilizados se utilizan a veces para determinar la temperatura a la que se deposita en el hueso, ya que la relación entre ciertos isótopos se correlaciona con la temperatura. En una muestra, las proporciones de isótopos en los huesos de diferentes partes del cuerpo indica una diferencia de temperatura de no más de 4 a 5 °C entre las vértebras del tronco y la tibia de la pierna. Este rango pequeño de temperatura entre el núcleo del cuerpo y las extremidades fue usado por el paleontólogo Reese Barrick y geoquímico William Showers para indicar que el Tyrannosaurus rex mantienia una temperatura constante del cuerpo, homeotermia y que disfrutaron de un metabolismo en algún lugar entre los reptiles ectotérmicos y los mamíferos endotérmicos.[88] Otros científicos han señalado que la proporción de isótopos de oxígeno en los fósiles de hoy no representa necesariamente la misma relación en el pasado distante, y puede haber sido alterada durante o después de fosilización, diagénesis.[89] Barrick y Showers han defendido sus conclusiones en trabajos posteriores, encontrando resultados similares en otro dinosaurio terópodo de un continente diferente y a decenas de millones de años de separación, el Giganotosaurus.[90] Los dinosaurios ornitisquios también mostraron evidencia de homeotermia, mientras que los lagartos varanidos de la misma formación no.[91] Aunque el Tyrannosaurus rex no muestren indicios de homeotermia, no significa necesariamente que sea endotérmico. La termorregulación pueden también explicarse por gigantotermia, como en algunas especies de tortugas marinas que viven hoy.[92] [93]
Huellas
 Posible huella en Nuevo México.
Posible huella en Nuevo México.Dos huellas aisladas fosilizadas han sido asignadas provisionalmente a Tyrannosaurus. La primera fue descubierta en Philmont Scout Ranch, Nuevo Méxic o, en 1983 por el geólogo estadounidense Charles Pillmore. Originalmente se pensó que pertenecen a un hadrosáurido, el examen de la huella reveló una gran " huella de talón" desconocida de dinosaurio ornitópodo, y los rastros de lo que pudo haber sido un dedo gordo, el espolón, como cuarto dígito del pie de tiranosaurio. La huella fue publicado bajo el título icnogénero Tyrannosauripus pillmorei en 1994, por Martin Lockley y Adrian Hunt. Lockley y Hunt sugirieron que era muy probable que fuera hecha por un Tyrannosaurus rex, lo que la convierte en la primera huella conocida de esta especie. La huella se imprimio en lo que fue una vez barro de los humedales con vegetación. Mide 83 centímetros de largo por 71 centímetros de ancho.[94]
Una segunda huella que pudo haber sido hecha por un Tyrannosaurus fue reportada por primera vez en 2007 por el paleontólogo británico Phil Manning, de la Formación Hell Creek de Montana. Esta segunda huella de 76 centímetros de largo, más corta que la descrita por Lockley y Hunt. Sea o no la huella fue hecha por Tyrannosaurus no está claro, aunque Tyrannosaurus y Nanotyrannus son los únicos grandes terópodos conocidos en la Formación Hell Creek. El estudio adicional de la huella, a descripción completa aún no ha sido publicada, compara la huella de Montana con la que se encuentra en Nuevo México.[95]
Locomoción
 La pata derecha del T. rex (lateral) fotografiada en el Museo de Historia Natural de la Universidad de Oxford.
La pata derecha del T. rex (lateral) fotografiada en el Museo de Historia Natural de la Universidad de Oxford.Christiansen (1998) estima que los huesos de la pierna de Tyrannosaurus no fueron significativamente más fuertes que los de los elefantes, que son relativamente limitadas en su velocidad máxima y nunca corren ya que no presenta la fase de aire, y por lo tanto propone que la velocidad máxima de los dinosaurios han sido cerca de 11 metros por segundo (40 km/h), lo que se refiere a la velocidad de un velocista humano. Pero también señaló que estas estimaciones dependen de muchos supuestos dudosos.[96]
Farlow y sus colegas (1995) han argumentado que un Tyrannosaurus peso 5,4 toneladas a 7,3 toneladas que han sido gravemente o incluso fatalmente herido si había caído mientras se mueve con rapidez, ya que su torso se habría estrellado contra el suelo a una desaceleración de 6 g (seis veces la aceleración de la gravedad, o alrededor de 60 m/s y los brazos pequeños no podrían haber reducido el impacto.[8] Sin embargo, las jirafas se han conocido a galopar a 50 kilómetros por hora, a pesar del riesgo de que puedan romper una pierna o algo peor, llegando a ser mortal, incluso en un ambiente "seguro" como un zoológico.[97] [98] Por lo tanto, es muy posible que también Tyrannosaurus se trasladaba rápidamente cuando sea necesario y tuvo que aceptar tales riesgos.[99] [100]
La mayoría de las investigaciones recientes sobre la locomoción de Tyrannosaurus no es compatible con una velocidad de 40 kilómetros por hora, es decir, ejecutando una moderada velocidad. Por ejemplo, un trabajo de 2002 en la revista Nature, utilizó un modelo matemático, validado por aplicación a tres animales que vivos, caimanes, aves y seres humanos; además, más adelante otros ocho especies, incluyendo emúes y avestruces para medir la masa del músculo de la pierna necesarios para una rápida ejecución más de 40 km/h.[101] [102] Encontraron que propone una velocidad máxima de más de 40 kilómetros por hora eran inviables, porque requeriría músculos de las piernas muy grandes de aproximadamente más del 40-86% de la masa total del cuerpo. Incluso moderada altas velocidades habría requerido grandes músculos de la pierna. Esta discusión es difícil de resolver, ya que no se sabe cuán grande es el músculos de las piernas en realidad estaban en Tyrannosaurus. Si fueran más pequeñas, sólo 18 kilómetros por hora para caminar o trotar posiblemente.[102]
 Un pollo de 6 toneladas habría necesitado músculos de las piernas que constituyeran casi el 100% de su masa corporal para correr. Siendo realistas, el T. rex tenía los músculos para correr a unos 5 metros por segundo (18 km/h, 11 mph).[102]
Un pollo de 6 toneladas habría necesitado músculos de las piernas que constituyeran casi el 100% de su masa corporal para correr. Siendo realistas, el T. rex tenía los músculos para correr a unos 5 metros por segundo (18 km/h, 11 mph).[102]Un estudio realizado en 2007 los modelos informáticos utilizados para calcular la velocidad de marcha, con base en datos obtenidos directamente de los fósiles, y afirmó que el Tyrannosaurus tenía una velocidad máxima de funcionamiento de 8 metros por segundo. El promedio de un fútbolista profesional sería un poco más lento, mientras que un velocista humano puede alcanzar los 12 metros por segundo (43 km/h). Tenga en cuenta que estos modelos de computadora predicen una velocidad máxima de 17,8 metros por segundo (64 km/h) para un pequeño Compsognathus de 3 kg probablemente un individuo juvenil.[103] [104] (probably a juvenile individual).[105] Aquellos que argumentan que el Tyrannosaurus era incapaz de correr estimando la velocidad máxima en alrededor de 17 kilómetros por hora. Esto es todavía más rápido que su más probable especies presa, hadrosáuridos y ceratopsianos.[102] Además, algunos defensores de la idea de que el Tyrannosaurus era un depredador afirman que la velocidad de los tiranosaurios en persecución no es importante, ya que puede haber sido lento, pero aún más rápido que su presa probable.[106] Sin embargo, Paul y Christiansen (2000) argumentó que al menos los ceratopsianos más tardi tuvieron patas delanteras en posición vertical y las especies más grandes pueden haber sido tan rápido como los rinocerontes. ]].[107] Heridas de mordedura curadas en los fósiles de ceratopsianos se interpretan como evidencia de ataques de tiranosaurios a ceratopsianos durante la vida. Si los ceratopsianos que vivieron junto a Tyrannosaurus eran rápidos, que pone en duda el argumento de que el Tyrannosaurus no tenía que ser rápido para atrapar a su presa.[100]
Estrategias de alimentación
 Mandíbula de tiranosaurio.
Mandíbula de tiranosaurio.Las grandes mandíbulas del tiranosaurio medían 1,4 m y estaban llenas de afilados dientes curvos de 19 cm. Estudios recientes de finales de 2007 y principios de 2008, sugieren que este dinosaurio tuvo la mordedura más poderosa de todos los depredadores, con una fuerza de presión de más de 4 toneladas,[108] aunque un pliosaurio, denominado depredador X encontrado en 2009 en el ártico podría tener haber ejercido una presión con su mordida 4 veces mayor, además de haber sido más grande.[109] [110]
 Un Allosaurus devorando carroña de un saurópodo.
Un Allosaurus devorando carroña de un saurópodo.¿Carroñero o Cazador?
El debate sobre si el Tyrannosaurus era un depredador o un carroñero puro es tan antiguo como el debate sobre su locomoción. Lambe (1917) describieron un buen esqueleto de un pariente cercano de Tyrannosaurus, Gorgosaurus y concluyó que era un carroñero puro, por lo que también Tyrannosaurus, ya que Gorgosaurus mostraba muy poco desgaste en los dientes.[111] Este argumento ya no es tomado en serio, porque los terópodos sustituyen los dientes continuamente. Desde el primer descubrimiento de la mayoría de los científicos han especulado que el Tyrannosaurus era un depredador;. Los grandes depredadores modernos limpian cadáveres que encuentran fácilmente o roban a otro depredador si tienen la oportunidad[112] Jack Horner experto en hadrosáuridos es actualmente el principal defensor de la idea de que Tyrannosaurus era exclusivamente carroñero y no realizaba caza activa.[30] [113] [114] Horner ha presentado varios argumentos para apoyar su hipótesis:
- Los brazos de Tyrannosaurus son cortos en comparación con otros depredadores conocidos. Horner afirma que los brazos eran demasiado cortos para que la fuerza de agarre necesario para aferrarse a su presa.[115]
- Tyrannosaurus tenía grandes bulbos olfatorios y nervios olfativos en relación con su tamaño cerebral. Esto sugiere un sentido del olfato muy desarrollado con lo que podría detectar cadáveres a grandes distancias, como los buitres modernos lo hacen. La investigación sobre los bulbos olfatorios de los dinosaurios ha demostrado que el Tyrannosaurus tenía el más sentido desarrollado del olfato entre los 21 dinosaurios en la muestra.[116] Los opositores de la hipótesis del carroñero puro han usado el ejemplo de los buitres en el sentido contrario, argumentando que la hipótesis del carroñero es inverosímil, porque los carroñeros puros modernas son sólo grandes aves planeadoras, que utilizan sus aguzados sentidos y la energía eficiente del planeo para cubrir amplias zonas económicamente.[117] Sin embargo, investigadores de Glasgow llegaron a la conclusión de que un ecosistema tan productivo como el actual Serengeti proporcionaría suficiente carroña para un gran terópodos carroñero, aunque los terópodos deberían de haber sido de sangre fría con el fin de obtener más calorías de la comida de lo que gasta en alimentarse. También sugirieron que en los ecosistemas modernos como Serengeti no hay carroñeros terrestres grandes, porque las aves planeadoras ahora hacen el trabajo mucho más eficientemente, mientras que los terópodos grandes no se enfrentaban a la competencia por el nicho ecológico de las aves carroñeras planeadoras.[118]
- Los dientes de Tyrannosaurus podrían aplastar huesos, y por lo tanto podría extraer tanta comida, médula ósea, como sea posible de los restos de un animal, por lo general las partes menos nutritivas. Karen Chin y sus colegas han encontrado fragmentos de hueso en coprolitos que atribuyen a tiranosaurios, pero señalan que los dientes de un Tyrannosaurus no se adaptaron bien a la manera sistemática de la masticación de los huesos de las hienas para extraer la médula.[119]
- Dado que al menos algunas de presas potenciales de Tyrannosaurus podrían haberse movido rápidamente, las pruebas de que caminaba en lugar de correr podrían indicar que era un carroñero.[113] [120] Por otro lado, los análisis recientes sugieren que Tyrannosaurus, aunque era más lento que los grandes depredadores terrestres modernos, bien pudo haber sido lo suficientemente rápido como para cazar hadrosáuridos grandes y ceratopsianos.[102] [106]
Otra evidencia sugiere el comportamiento de caza en Tyrannosaurus. Las órbitas de Tyrannosaurus se colocan de manera que los ojos miren hacia adelante, dándoles una visión binocular ligeramente mejor que la de los halcones modernos. También señaló que el linaje de Tyrannosaurus tenía una historia de constante mejora de la visión binocular. No es obvio por qué la selección natural habría favorecido esta tendencia a largo plazo si los Tyrannosaurus habrían sido carroñeros puros, que no habría necesitado la percepción de la profundidad avanzada que proporciona la visión estereoscópica.[17] [18] En los animales modernos, la visión binocular se encuentra principalmente en los depredadores.
 Restauración basada en el ejemplar MOR 980, con infecciones parasitarias, que causaron las cicatrices encontradas en muchos cráneos anteriormente explicadas como señales de luchas interespecificas.
Restauración basada en el ejemplar MOR 980, con infecciones parasitarias, que causaron las cicatrices encontradas en muchos cráneos anteriormente explicadas como señales de luchas interespecificas.Un esqueleto del hadrosáurido Edmontosaurus annectens descrito de Montana aparece con una lesión infligida por un Tyrannosaurus y curada en sus vértebras de la cola. El hecho de que el daño parece haber sanado sugiere que el Edmontosaurus sobrevivió al ataque de un Tyrannosaurus durante su vida, es decir, el Tyrannosaurus había intentado depredación activa.[121] También hay evidencia de una interacción agresiva entre Triceratops y Tyrannosaurus, marcas parcialmente curadas de dientes de Tyrannosaurus aparecen en el cuerno frontal y escamoso, un hueso del volante del cuello, de un Triceratops; el cuerno mordido estaba roto, con un crecimiento de hueso nuevo en la fractura. No se sabe cuál fue la naturaleza exacta de la interacción, sin embargo el animal podría haber sido el agresor.[122] Al examinara a Sue, el paleontólogo Peter Larson encontró una fractura sanada en el peroné y las vértebras de la cola, cicatrices en los huesos de la cara y un diente de otro Tyrannosaurus incrustado en una vértebra del cuello. Si es correcto, estos podrían ser una fuerte evidencia de comportamiento agresivo entre Tyrannosaurus, pero si fue competencia por el alimento o compañeros o canibalismo activo no está claro.[123] Sin embargo, la investigación más reciente de estas supuestas heridas ha demostrado que la mayoría son infecciones en lugar de lesiones o simplemente daños en los fósiles después de la muerte y las pocas lesiones son demasiado generales para ser indicativo de un conflicto intraespecífico.[113] Un estudio de 2009 demostró que los agujeros en los cráneos de varios especímenes pueden haber sido causadas por parásitos como las Trichomonas que normalmente infectan a las aves. Algunos investigadores sostienen que si el Tyrannosaurus era un carroñero, otro dinosaurio tenía que ser el mayor depredador en el Cretácico superior en Laurasia. Las presas más grandes eran los marginocéfalos y ornitopódos. Los tiranosáuridos compartieron tantas características que sólo los pequeños dromeosáuridos son los posibles principales depredadores. En este sentido, los partidarios de la hipótesis del carroñero puro han sugerido que el tamaño y el poder de Tyrannosaurus era suficiente para robar las presas de los pequeños depredadores.[120] La mayoría de los paleontólogos aceptan que Tyrannosaurus era a la vez un activo depredador y un carroñero como la mayoría de los grandes carnívoros.
Muchos científicos que han publicado sobre el tema insisten en que el tiranosaurio fue tanto depredador como carroñero, alimentándose de cualquier carne que pudiera conseguir, dependiendo de la oportunidad que se le presentara.[124] Los carnívoros modernos raras veces son estrictos depredadores o carroñeros. Los leones, por ejemplo, a veces comen hienas muertas y viceversa. El comportamiento depende de la disponibilidad de la presa, entre otros factores. Si los tiranosauros eran carroñeros que practicaban cleptoparasitismo (robo de las presas cazadas por auténticos depredadores) su masa corporal habría sido un factor intimidante para ahuyentar a los depredadores; los depredadores coetáneos indiscutibles (por ejemplo los raptores) eran mucho más pequeños y veloces, por lo que la presencia de un carroñero gigante dotado de grandes dientes les habría hecho huir o retroceder.
 Reconstrucción de la cabeza del T. rex, expuesta en el Museo de Historia Natural de la Universidad de Oxford.
Reconstrucción de la cabeza del T. rex, expuesta en el Museo de Historia Natural de la Universidad de Oxford.No obstante, se suponen pruebas de comportamiento cazador en el tiranosaurio, como por ejemplo la visión binocular mencionada más arriba. Debido a que los carroñeros no necesitan la compleja percepción de profundidad que brinda la visión estereoscópica, en los animales modernos la visión binocular se halla principalmente en los depredadores. Sin embargo, no es exclusiva de los mismos (lemúridos y primates, entre otros no depredadores, poseen visión binocular estereoscópica). Además, el descubrimiento de marcas de mordidas en otros animales, e incluso en otros tiranosaurios, ha sugerido un comportamiento predatorio.
Además la teoría de que fuera un exclusivo carroñero no es del todo posible, ya que si era completamente carroñero, no podría haber sobrevivido, ya que no existían carnívoros suficientemente grandes en su ecosistema que pudieran cazar presas tan grandes para abastecerse ellos mismos y a una población saludable de tiranosaurios, además de que no hay animales muriendo todos los días para la misma causa. Por otra parte, recientes estudios de la mandíbula y la presión que esta ejercía, refutan que fuera débil y sin fuerza suficiente para abatir grandes presas. De hecho era el más fuerte de todos los depredadores terrestres conocidos. Además, no necesitaba ser rápido, por que sus presas tampoco lo eran, sino extremadamente poderoso para derribarlas. Así que el Tyrannosaurus fue un depredador que evolucionó a la par con sus presas, y bien podría acechar a sus víctimas oculto y usando el olfato, que además debió haber servido, en parte para ayudarlo a localizar animales muertos que presentaban una fuente de comida fácil, o para localizar a sus presas a distancia para evitar ser descubierto. Finalmente, en cuanto a los brazos extremadamente pequeños, la evolución de los tiranosáuridos llevó a la reducción de éstos para equilibrar el peso de la cabeza con el del resto del cuerpo, por lo que si no se hubieran extinguido, los brazos de los tiranosaurios se hubieran reducido por completo y sus cuerpos se hubieran vuelto aún más grandes. Estas características convirtieron al Tyrannosaurus, en lugar de un carroñero, en la máxima máquina de matar de artillería pesada del mundo de los dinosaurios, más eficiente que los anteriores megalosaurios, alosaurios, carcarodontosaurios y espinosaurios.
Por lo que en conclusión, el Tyrannosaurus era un formidable depredador y un carroñero cuando tuviera la oportunidad.
Canibalismo
Un estudio de Currie, Horner, Erickson y Longrich en 2010 se ha presentado como prueba de canibalismo en el género Tyrannosaurus.[125] Ellos estudiaron algunos ejemplares de Tyrannosaurus con marcas de dientes en los huesos, atribuibles al mismo género. Las marcas de dientes fueron identificados en el húmero, huesos del pie y metatarsos, y esto fue visto como una prueba de carroñeros oportunistas, en lugar de las heridas causadas por el combate intraespecífico. En una pelea, que propone que sería difícil llegar a bajar a morder en los pies de un rival, por lo que es más probable que las marcas de dientes se hicieron en un cadáver. Debido a que las marcas se realizaron en otras partes del cuerpo con relativamente escasamente cantidades de carne, se sugiere que el Tyrannosaurus se alimentaba de un cadáver en el que las partes más carnosas ya se había comido.[125] También fueron abiertos a la posibilidad de que otros tiranosáuridos practicaba el canibalismo.
Saliva Infecciosa
Tyrannosaurus puede haber tenido la saliva infecciosa para matar a sus presas. Esta teoría fue propuesta por primera vez por William Abler[126] Abler examinar los dientes de los tiranosáuridos entre cada denticulo de la sierra de los dientes queda un espacio que podría haber alojado parte de carne de la alimentación en estado de putrefacción con bacterias, dando al Tyrannosaurus una mordedura infecciosa mortal, muy parecido al dragón de Komodo. Sin embargo, Jack Horner se refiere estrías Tyrannosaurus diente más como cubos en forma de las estrías en los dientes de un monitor de Komodo, que se redondean.[127] Horner argumenta que el diente es sólido comparado con el diente con canales del dragón de komodo.[128]
En la cultura popular
Desde que fuera descrito por primera vez en 1905, el Tyrannosaurus rex se ha convertido en la especie lo más reconocida de dinosaurio en la cultura popular. Es el único dinosaurio que es conocido comúnmente por público en general por su nombre científico completo (nombre binomial) (Tyrannosaurus rex), y su abreviatura científica T. rex también ha encontrado un uso amplio.[14] Robert Bakker nota esto en su libro Herejías de dinosaurio y explica que un nombre como el de Tyrannosaurus rex "es irresistible a pronunciar".[6]
Referencias
- ↑ Unearthing T. rex : T.rex In-Depth : Traits. Retrieved on December 11, 2005.
- ↑ Erickson, G.M., Makovicky, P.J., Currie, P.J., Norell, M.A., Yerby, S.A., & Brochu, C.A. 2004. Gigantism and comparative life-history parameters of tyrannosaurid dinosaurs. Nature 430: 772-775.
- ↑ a b «Sue's vital statistics». Sue at the Field Museum. Field Museum of Natural History. Consultado el 15-09-2007.
- ↑ Henderson DM (1 de enero de 1999). «Estimating the masses and centers of mass of extinct animals by 3-D mathematical slicing». Paleobiology 25 (1): pp. 88–106. http://paleobiol.geoscienceworld.org/cgi/content/abstract/25/1/88.
- ↑ Anderson, JF; Hall-Martin AJ Russell DA (1985). «Long bone circumference and weight in mammals, birds and dinosaurs». Journal of Zoology 207 (1): pp. 53–61.
- ↑ a b c Bakker, Robert T. (1986). The Dinosaur Heresies. New York: Kensington Publishing. ISBN 0-688-04287-2. OCLC 13699558.Plantilla:Pn
- ↑ a b c Erickson, G.M., Makovicky, P.J., Currie, P.J., Norell, M.A., Yerby, S.A. & Brochu, C.A. 2004. Gigantism and comparative life-history parameters of tyrannosaurid dinosaurs. Nature 430: 772–775.
- ↑ a b Farlow, JO; Smith MB, Robinson JM (1995). «Body mass, bone "strength indicator", and cursorial potential of Tyrannosaurus rex». Journal of Vertebrate Paleontology 15 (4): pp. 713–725. http://www.vertpaleo.org/publications/jvp/15-713-725.cfm.
- ↑ Seebacher, Frank. (2001). «A new method to calculate allometric length-mass relationships of dinosaurs». Journal of Vertebrate Paleontology 21 (1): pp. 51–60. doi:.
- ↑ Christiansen, Per; & Fariña, Richard A. (2004). «Mass prediction in theropod dinosaurs». Historical Biology 16 (2–4): pp. 85–92. doi:.
- ↑ Hutchinson J.R., Bates K.T., Molnar J., Allen V & Makovicky P.J. (2011), "A Computational Analysis of Limb and Body Dimensions in Tyrannosaurus rex with Implications for Locomotion, Ontogeny, and Growth", PLoS ONE 6(10): e26037. doi:10.1371/journal.pone.0026037
- ↑ dal Sasso, Cristiano; Maganuco, Simone; Buffetaut, Eric; & Méndez, Marcos A. (2005). «New information on the skull of the enigmatic theropod Spinosaurus, with remarks on its sizes and affinities». Journal of Vertebrate Paleontology 25 (4): pp. 888–896. doi:. http://www.bioone.org/perlserv/?request=get-abstract&doi=10.1671%2F0272-4634%282005%29025%5B0888%3ANIOTSO%5D2.0.CO%3B2.
- ↑ Calvo, Jorge O.; Rodolfo Coria (December 1998). «New specimen of Giganotosaurus carolinii (Coria & Salgado, 1995), supports it as the largest theropod ever found» (pdf). Gaia Revista de Geociências 15: pp. 117–122. http://www.mnhn.ul.pt/geologia/gaia/7.pdf.
- ↑ a b c d e Brochu, Christopher A.; Richard A. Ketcham (2003). Osteology of Tyrannosaurus Rex: Insights from a Nearly Complete Skeleton and High-resolution Computed Tomographic Analysis of the Skull. Northbrook, Illinois: Society of Vertebrate Paleontology. OCLC 51651461.
- ↑ Lipkin, Christine; and Carpenter, Kenneth (2008). «Looking again at the forelimb of Tyrannosaurus rex». Tyrannosaurus rex, the Tyrant King (Life of the Past). Bloomington: Indiana University Press. pp. 167–190. ISBN 0-253-35087-5.
- ↑ Montana State University (2006-04-07). «Museum unveils world's largest T-rex skull». Comunicado de prensa. Consultado el 2008-09-13.
- ↑ a b Stevens, Kent A. (June 2006). «Binocular vision in theropod dinosaurs» (PDF). Journal of Vertebrate Paleontology 26 (2): pp. 321–330. doi:. http://www.geocities.com/Athens/Bridge/4602/theropod_binocularvision.pdf.
- ↑ a b Jaffe, Eric (01-07-2006). «Sight for 'Saur Eyes: T. rex vision was among nature's best». Science News 170 (1): pp. 3. doi:. http://www.sciencenews.org/view/generic/id/7500/title/Sight_for_Saur_Eyes_%3Ci%3ET._rex%3Ci%3E_vision_was_among_natures_best.
- ↑ Snively, Eric; Donald M. Henderson, and Doug S. Phillips (2006). «Fused and vaulted nasals of tyrannosaurid dinosaurs: Implications for cranial strength and feeding mechanics» (PDF). Acta Palaeontologica Polonica 51 (3): pp. 435–454. http://www.app.pan.pl/archive/published/app51/app51-435.pdf.
- ↑ Erickson, G.M.; Van Kirk, S.D.; Su, J.; Levenston, M.E.; Caler, W.E.; and Carter, D.R. (1996). «Bite-force estimation for Tyrannosaurus rex from tooth-marked bones». Nature 382: pp. 706–708. doi:.
- ↑ Meers, M.B. (August 2003). «Maximum bite force and prey size of Tyrannosaurus rex and their relationships to the inference of feeding behavior». Historical Biology: A Journal of Paleobiology 16 (1): pp. 1–12. doi:. http://www.ingentaconnect.com/content/tandf/ghbi/2003/00000016/00000001/art00001.
- ↑ Brochu, C.R. (2003). «Osteology of Tyrannosaurus rex: insights from a nearly complete skeleton and high-resolution computed tomographic analysis of the skull». Society of Vertebrate Paleontology Memoirs 7: pp. 1–138. doi:.
- ↑ Smith, J.B. (December 2005). «Heterodonty in Tyrannosaurus rex: implications for the taxonomic and systematic utility of theropod dentitions» (PDF). Journal of Vertebrate Paleontology 25 (4): pp. 865–887. doi:. http://www.geocities.com/Athens/Bridge/4602/trexteeth.pdf.
- ↑ Douglas K, Young S (1998). «The dinosaur detectives». New Scientist. http://www.newscientist.com/channel/life/dinosaurs/mg15821305.300. «One palaeontologist memorably described the huge, curved teeth of T. rex as 'lethal bananas'».
- ↑ a b In the Field, edición de enero-febrero de 2005, Museo Field de Historia Natural.
- ↑ «The First Tyrannosaurus Skeleton, 1905». Linda Hall Library of Science, Engineering and Technology. Consultado el 03-08-2008.
- ↑ «In Celebration of 100 years of Tyrannosaurus Rex: Manospondylus Gigas, Ornithomimus Grandis, and Dynamosaurus Imperiosus, the Earliest Discoveries of Tyrannosaurus Rex in the West». Abstracts with Programs. 2005 Salt Lake City Annual Meeting. http://gsa.confex.com/gsa/2005AM/finalprogram/abstract_96150.htm.
- ↑ Osborn, H. F. (1905). «Tyrannosaurus and other Cretaceous carnivorous dinosaurs». Bulletin of the American Museum of Natural History 21: pp. 259–265.
- ↑ Breithaup, BH; Southwell EH, Matthews NA (2006). «Dynamosaurus Imperiosus and the earliest discoveries of Tyrannosaurus Rex in Wyoming and the West» (pdf). New Mexico Museum of Natural History and Science Bulletin 35: pp. 257–258. http://www.nmnaturalhistory.org/science/bulletins/35/sci_bulletin35_32.pdf.
- ↑ a b c Horner, John R.; Don Lessem (1993). The complete T. rex. New York City: Simon & Schuster. ISBN 0-671-74185-3.Plantilla:Pn
- ↑ a b Osborn, H. F. (1917). «Skeletal adaptations of Ornitholestes, Struthiomimus, Tyrannosaurus». Bulletin of the American Museum of Natural History (New York City: American Museum of Natural History) 35 (43): pp. 733–771. http://hdl.handle.net/2246/1334.
- ↑ Ride, W. D. L. (1999). «Article 23.9 – Reversal of Precedence». International code of zoological nomenclature. London: International Commission on Zoological Nomenclature. ISBN 0-85301-006-4. OCLC 183090345.
- ↑ Taylor, Mike (27-08-2002). «So why hasn't Tyrannosaurus been renamed Manospondylus?». The Dinosaur FAQ. Consultado el 08-10-2008.
- ↑ «So why hasn't Tyrannosaurus been renamed Manospondylus?». Consultado el December 11 de 2005.
- ↑ a b Horner, J.R. & Lessem, D. 1993. The Complete T. rex New York: Simon & Schuster. 238pp.
- ↑ "STAN T. rex" Black Hills Institute for Geological Research, Inc. 2004. Retrieved July 16, 2005.
- ↑ Brochui, C.A. (December 2003). «Lessons From A Tyrannosaur: The Ambassadorial Role Of Paleontology». PALAIOS 18 (6): pp. 475. doi:. http://www.bioone.org/perlserv/?request=get-document&doi=10.1669%2F0883-1351(2003)018%3C0475%3ALFATTA%3E2.0.CO%3B2.
- ↑ [1]
- ↑ "Dig pulls up five T. rex specimens". BBC News. Octubre 10 de 2000.
- ↑ «BBC NEWS : Science/Nature : T. rex fossil has 'soft tissues'». Consultado el December 11 de 2005.
- ↑ «http://www.washingtonpost.com/wp-dyn/content/article/2005/06/03/AR2005060300141.html». Consultado el December 11 de 2005.
- ↑ «Footprint of a Giant». Online guide to the continental Cretaceous-Tertiary boundary in the Raton basin, Colorado and New Mexico. United States Geological Survey. Consultado el 09-10-2008.
- ↑ a b Currie, Philip J.; Jørn H. Hurum and Karol Sabath (2003). «Skull structure and evolution in tyrannosaurid dinosaurs» (PDF). Acta Palaeontologica Polonica 48 (2): pp. 227–234. http://www.app.pan.pl/archive/published/app48/app48-227.pdf.
- ↑ a b c Holtz, Thomas R., Jr. (2004). «Tyrannosauroidea». En David B. Weishampel, Peter Dodson and Halszka Osmólska. The dinosauria. Berkeley: University of California Press. pp. 111–136. ISBN 0-520-24209-2.
- ↑ a b c Paul, Gregory S. (1988). Predatory dinosaurs of the world: a complete illustrated guide. New York: Simon and Schuster. ISBN 0-671-61946-2. OCLC 18350868.Plantilla:Pn
- ↑ a b Holtz, Thomas R. (1994). «The Phylogenetic Position of the Tyrannosauridae: Implications for Theropod Systematics». Journal of Palaeontology 68 (5): pp. 1100–1117. doi:. http://www.jstor.org/pss/1306180.
- ↑ Maleev, E. A. (1955). «[Gigantic carnivorous dinosaurs of Mongolia]» (en ruso). Doklady Akademii Nauk SSSR 104 (4): pp. 634–637.
- ↑ Rozhdestvensky, AK (1965). «Growth changes in Asian dinosaurs and some problems of their taxonomy». Paleontological Journal 3: pp. 95–109.
- ↑ Carpenter, Kenneth (1992). «Tyrannosaurids (Dinosauria) of Asia and North America». En Niall J. Mateer and Pei-ji Chen. Aspects of nonmarine Cretaceous geology. Beijing: China Ocean Press. ISBN 9787502714635. OCLC 28260578.
- ↑ Carr, Thomas D.; Thomas E. Williamson and David R. Schwimmer (March 2005). «A New Genus and Species of Tyrannosauroid from the Late Cretaceous (Middle Campanian) Demopolis Formation of Alabama». Journal of Vertebrate Paleontology 25 (1): pp. 119–143. doi:.
- ↑ Hurum, Jørn H.; Karol Sabath (2003). «Giant theropod dinosaurs from Asia and North America: Skulls of Tarbosaurus bataar and Tyrannosaurus rex compared» (PDF). Acta Palaeontologica Polonica 48 (2): pp. 161–190. http://www.app.pan.pl/archive/published/app48/app48-161.pdf.
- ↑ Olshevsky, George (1995). «The origin and evolution of the tyrannosaurids». Kyoryugaku Saizensen [Dino Frontline] 9–10: pp. 92–119.
- ↑ Carr, T.D.; T.E. Williamson (2004). «Diversity of late Maastrichtian Tyrannosauridae (Dinosauria: Theropoda) from western North America». Zoological Journal of the Linnean Society 142: pp. 479–523. doi:.
- ↑ Gilmore, C.W. (1946). «A new carnivorous dinosaur from the Lance Formation of Montana». Smithsonian Miscellaneous Collections 106: pp. 1–19.
- ↑ Bakker, R.T.; M. Williams and P.J. Currie (1988). «Nanotyrannus, a new genus of pygmy tyrannosaur, from the latest Cretaceous of Montana». Hunteria 1 (5): pp. 1–30.
- ↑ Carr, TD (1999). «Craniofacial ontogeny in Tyrannosauridae (Dinosauria, Theropoda)». Journal of Vertebrate Paleontology 19: pp. 497–520.
- ↑ Currie, Philip J. (2003). «Cranial anatomy of tyrannosaurid dinosaurs from the Late Cretaceous of Alberta, Canada» (PDF). Acta Palaeontologica Polonica 42 (2): pp. 191–226. http://www.app.pan.pl/archive/published/app48/app48-191.pdf.
- ↑ «L A F A U N A».
- ↑ Scientific American, 290, no. 2, febrero de 2004 pp. 23-24
- ↑ Leidy, J (1865). «Memoir on the extinct reptiles of the Cretaceous formations of the United States». Smithsonian Contributions to Knowledge 14: pp. 1–135.
- ↑ «Tyrannosaurus». American Museum of Natural History. Consultado el 16-10-2008.
- ↑ a b Newman, BH (1970). «Stance and gait in the flesh-eating Tyrannosaurus». Biological Journal of the Linnean Society 2: pp. 119–123. doi:.
- ↑ «The Age of Reptiles Mural». Yale University (2008). Consultado el 16-10-2008.
- ↑ Osborn, H. F. (1905). «Tyrannosaurus and other Cretaceous carnivorous dinosaurs». Bulletin of the AMNH (New York City: American Museum of Natural History) 21 (14): pp. 259–265. http://hdl.handle.net/2246/1464.
- ↑ Lambe, L. M. (1914). «On a new genus and species of carnivorous dinosaur from the Belly River Formation of Alberta, with a description of the skull of Stephanosaurus marginatus from the same horizon». Ottawa Naturalist 27: pp. 129–135.
- ↑ Osborn, Henry Fairfield; Barnum Brown (1906). «Tyrannosaurus, Upper Cretaceous carnivorous dinosaur». Bulletin of the AMNH (New York City: American Museum of Natural History) 22 (16): pp. 281–296. http://hdl.handle.net/2246/1473.
- ↑ Carpenter, Kenneth; Matt Smith (2001). «Forelimb Osteology and Biomechanics of Tyrannosaurus rex». En Darren Tanke and Kenneth Carpenter. Mesozoic vertebrate life. Bloomington: Indiana University Press. pp. 90–116. ISBN 0-253-33907-3.
- ↑ a b Horner JR, Padian K (September 2004). «Age and growth dynamics of Tyrannosaurus rex». Proceedings. Biological sciences / the Royal Society 271 (1551): pp. 1875–80. doi:. PMID 15347508. PMC 1691809. http://journals.royalsociety.org/openurl.asp?genre=article&issn=0962-8452&volumen=271&número=1551&spage=1875.
- ↑ a b Schweitzer MH, Wittmeyer JL, Horner JR (June 2005). «Gender-specific reproductive tissue in ratites and Tyrannosaurus rex». Science (New York, N.Y.) 308 (5727): pp. 1456–60. doi:. PMID 15933198. http://www.sciencemag.org/cgi/pmidlookup?view=long&pmid=15933198.
- ↑ Lee, Andrew H.; and Werning, Sarah (2008). «Sexual maturity in growing dinosaurs does not fit reptilian growth models». Proceedings of the National Academy of Sciences 105 (2): pp. 582–587. doi:. PMID 18195356.
- ↑ a b Erickson GM, Currie PJ, Inouye BD, Winn AA (July 2006). «Tyrannosaur life tables: an example of nonavian dinosaur population biology». Science (New York, N.Y.) 313 (5784): pp. 213–7. doi:. PMID 16840697.
- ↑ Carpenter, Kenneth (1992). «Variation in Tyrannosaurus rex». En Kenneth Carpenter and Philip J. Currie. Dinosaur Systematics: Approaches and Perspectives. Cambridge: Cambridge University Press. pp. 141–145. ISBN 0-521-43810-1.
- ↑ Larson, P.L. 1994. Tyrannosaurus sex. In: Rosenberg, G.D. & Wolberg, D.L. Dino Fest. The Paleontological Society Special Publications. 7: 139–155.
- ↑ Erickson GM, Kristopher Lappin A, Larson P (2005). «Androgynous rex – the utility of chevrons for determining the sex of crocodilians and non-avian dinosaurs». Zoology (Jena, Germany) 108 (4): pp. 277–86. doi:. PMID 16351976. http://linkinghub.elsevier.com/retrieve/pii/S0944-2006(05)00047-4.
- ↑ Schweitzer MH, Elsey RM, Dacke CG, Horner JR, Lamm ET (April 2007). «Do egg-laying crocodilian (Alligator mississippiensis) archosaurs form medullary bone?». Bone 40 (4): pp. 1152–8. doi:. PMID 17223615. http://linkinghub.elsevier.com/retrieve/pii/S8756-3282(06)00748-4.
- ↑ a b Fields, Helen (May 2006). «Dinosaur Shocker». Smithsonian Magazine. http://www.smithsonianmag.com/science-nature/10021606.html.
- ↑ Schweitzer, Mary H.; Jennifer L. Wittmeyer, John R. Horner and Jan K. Toporski (March 2005). «Soft-tissue vessels and cellular preservation in Tyrannosaurus rex». Science 307 (5717): pp. 1952–5. doi:. PMID 15790853. http://www.sciencemag.org/cgi/content/abstract/307/5717/1952.
- ↑ Rincon, Paul. «Protein links T. rex to chickens», BBC News, 12-04-2007. Consultado el 02-10-2008.
- ↑ Vergano, Dan. «Yesterday's T. Rex is today's chicken», USA Today, 13-04-2007. Consultado el 08-10-2008.
- ↑ Schmid, Randolph E., Associated Press. «Scientists study evidence modern birds came from dinosaurs», Newsvine, 24-04-2008. Consultado el 08-10-2008.
- ↑ Kaye, Thomas G.; Gary Gaugler and Zbigniew Sawlowicz (July 2008). «Dinosaurian Soft Tissues Interpreted as Bacterial Biofilms». ONE 3 (7): pp. e2808. doi:. PMID 18665236.
- ↑ Newswise (2008-07-24). «New Research Challenges Notion That Dinosaur Soft Tissues Still Survive». Comunicado de prensa. Consultado el 2008-10-08.
- ↑ Discover (2008-07-30). «Researchers Debate: Is It Preserved Dinosaur Tissue, or Bacterial Slime?». Comunicado de prensa. Consultado el 2008-09-04.
- ↑ Paul, Gregory S. (2008). «The extreme lifestyles and habits of the gigantic tyrannosaurid superpredators of the Late Cretaceous of North America and Asia». Tyrannosaurus rex, the Tyrant King (Life of the Past). Bloomington: Indiana University Press. p. 316. ISBN 0-253-35087-5.
- ↑ Xu, Xing; Mark A. Norell, Xuewen Kuang, Xiaolin Wang, Qi Zhao and Chengkai Jia (07-10-2004). «Basal tyrannosauroids from China and evidence for protofeathers in tyrannosauroids». Nature 431 (7009): pp. 680–684. doi:. PMID 15470426.
- ↑ Bakker, Robert T. (1968). «The superiority of dinosaurs» (PDF). Discovery 3 (2): pp. 11–12. Archivado del original el September 9, 2006. http://web.archive.org/web/20060909131058/http://bio.fsu.edu/~amarquez/Evolutionary%20Morphology%20fall%202004/Bakker/Bakker%201968%20-%20Superiority%20of%20DInos.pdf.
- ↑ Bakker, Robert T. (1972). «Anatomical and ecological evidence of endothermy in dinosaurs» (PDF). Nature 238: pp. 81–85. doi:. Archivado del original el September 9, 2006. http://web.archive.org/web/20060909173036/http://bio.fsu.edu/~amarquez/Evolutionary%20Morphology%20fall%202004/Bakker/14-%20Bakker%201972%20-%20dino%20endothermy.pdf.
- ↑ Barrick, Reese E.; William J. Showers (July 1994). «Thermophysiology of Tyrannosaurus rex: Evidence from Oxygen Isotopes». Science (New York City) 265 (5169): pp. 222–224. doi:. PMID 17750663. http://www.sciencemag.org/cgi/content/abstract/265/5169/222.
- ↑ Trueman, Clive; Carolyn Chenery, David A. Eberth and Baruch Spiro (2003). «Diagenetic effects on the oxygen isotope composition of bones of dinosaurs and other vertebrates recovered from terrestrial and marine sediments». Journal of the Geological Society 160 (6): pp. 895. doi:.
- ↑ Barrick, Reese E.; William J. Showers (October 1999). «Thermophysiology and biology of Giganotosaurus: comparison with Tyrannosaurus». Palaeontologia Electronica 2 (2). http://palaeo-electronica.org/1999_2/gigan/issue2_99.htm.
- ↑ Barrick, Reese E.; Michael K. Stoskopf and William J. Showers (1999). «Oxygen isotopes in dinosaur bones». En James O. Farlow and M. K. Brett-Surman. The Complete Dinosaur. Bloomington: Indiana University Press. pp. 474–490. ISBN 0-253-21313-4.
- ↑ Paladino, Frank V.; James R. Spotila and Peter Dodson (1999). «A blueprint for giants: modeling the physiology of large dinosaurs». En James O. Farlow and M. K. Brett-Surman. The Complete Dinosaur. Bloomington: Indiana University Press. pp. 491–504. ISBN 0-253-21313-4.
- ↑ Chinsamy, Anusuya; Willem J. Hillenius (2004). «Physiology of nonavian dinosaurs». En David B. Weishampel, Peter Dodson and Halszka Osmólska. The dinosauria. Berkeley: University of California Press. pp. 643–659. ISBN 0-520-24209-2.
- ↑ Lockley, MG; Hunt AP (1994). «A track of the giant theropod dinosaur Tyrannosaurus from close to the Cretaceous/Tertiary boundary, northern New Mexico». Ichnos 3: pp. 213–218. doi:.
- ↑ «T.rex footprint discovered». Natural History Museum (2007). Consultado el 09-12-2008.
- ↑ Christiansen, P. (1998). «Strength indicator values of theropod long bones, with comments on limb proportions and cursorial potential» (pdf). Gaia 15: pp. 241–255. ISSN 0871-5424. http://www.mnhn.ul.pt/geologia/gaia/19.pdf.
- ↑ «Giraffe». WildlifeSafari.info. Consultado el 29-04-2006.
- ↑ «The History of Woodland Park Zoo – Chapter 4». Consultado el 29-04-2006.
- ↑ Alexander, R.M. (7 de agosto de 2006). «Dinosaur biomechanics». Proc Biol Sci. (The Royal Society) 273 (1596): pp. 1849–1855. doi:. PMID 16822743.
- ↑ a b Hanna, Rebecca R. (2002). «Multiple injury and infection in a sub-adult theropod dinosaur (Allosaurus fragilis) with comparisons to allosaur pathology in the Cleveland-Lloyd dinosaur quarry collection». Journal of Vertebrate Paleontology 22 (1): pp. 76–90. doi:. catalogs the injuries of the Allosaurus known as "Big Al" – at least one was attributed to a fall.
- ↑ Hutchinson, J.R. (2004). «Biomechanical Modeling and Sensitivity Analysis of Bipedal Running Ability. II. Extinct Taxa» (PDF). Journal of Morphology 262 (1): pp. 441–461. doi:. PMID 15352202. http://www.rvc.ac.uk/AboutUs/Staff/jhutchinson/documents/JRH13.pdf.
- ↑ a b c d e Hutchinson JR, Garcia M (February 2002). «Tyrannosaurus was not a fast runner». Nature 415 (6875): pp. 1018–21. doi:. PMID 11875567.
- ↑ Sellers, W.I., and Manning, P.L. (July 2007). «Estimating dinosaur maximum running speeds using evolutionary robotics». Proc. R. Soc. B (The Royal Society) 274 (1626): pp. 2711. doi:. PMID 17711833. PMC 2279215. http://dinonews.net/rubriq/articles.php5?action=open&ref=2007_sellers_speed.
- ↑ Seward, L (21-08-2007). «T. rex 'would outrun footballer'». BBCNews. Consultado el 16-10-2008.
- ↑ Callison, G.; H. M. Quimby (1984). «Tiny dinosaurs: Are they fully grown?». Journal of Vertebrate Paleontology 3: pp. 200–209. doi:.
- ↑ a b Manning P (2008). «T. rex speed trap». En Carpenter, Kenneth; Larson, Peter E.. Tyrannosaurus rex, the Tyrant King (Life of the Past). Bloomington: Indiana University Press. pp. 205–228. ISBN 0-253-35087-5.
- ↑ Paul, G.S., and Christiansen, P. (September 2000). «Forelimb posture in neoceratopsian dinosaurs: implications for gait and locomotion». Paleobiology 26 (3): pp. 450. doi:. http://www.bioone.org/perlserv/?request=get-document&doi=10.1666%2F0094-8373(2000)026%3C0450%3AFPINDI%3E2.0.CO%3B2.
- ↑ Erickson GM, Van Kirk SD (1996). "Bite-Force Estimation for Tyrannosaurus rex from Tooth-Marked Bones". Nature 382: 706–708.
- ↑ Fósil monstruo marino destrona al Tiranosaurio. El Universo. 17 de marzo de 2009.
- ↑ http://www.abc.es/20090316/nacional-sociedad/mayor-depredador-historia-vivia-200903162236.html El mayor depredador de la historia vivía en el mar y pesaba 45 toneladas. ABC. 17 de marzo de 2009.
- ↑ Lambe, L. B. (1917). «The Cretaceous theropodous dinosaur Gorgosaurus». Memoirs of the Geological Survey of Canada 100: pp. 1–84.
- ↑ Farlow, J. O. and Holtz, T. R. (2002). «The Fossil Record of Predation» (pdf) págs. 251–266.
- ↑ a b c Horner, J.R. (1994). «Steak knives, beady eyes, and tiny little arms (a portrait of Tyrannosaurus as a scavenger)». The Paleontological Society Special Publication 7: pp. 157–164.
- ↑ Amos, J. (31-07-2003). «T. rex goes on trial». BBC.
- ↑ The king of killers or fearsome freeloader? It is one of the big questions currently in palaeontology.
- ↑ «T. Rex brain study reveals a refined "nose"». Calgary Herald (28-10-2008). Consultado el 29-10-2008.
- ↑ Paul, G.S. (1988). Predatory Dinosaurs of the World. Simon and Schuster. ISBN 0671619462. OCLC 18350868.
- ↑ Ruxton GD, Houston DC (April 2003). «Could Tyrannosaurus rex have been a scavenger rather than a predator? An energetics approach». Proceedings. Biological sciences / the Royal Society 270 (1516): pp. 731–3. doi:. PMID 12713747. PMC 1691292. http://journals.royalsociety.org/openurl.asp?genre=article&issn=0962-8452&volumen=270&número=1516&spage=731.
- ↑ Chin, K., Erickson, G.M. et al. (18-06-1998). «A king-sized theropod coprolite». Nature 393: pp. 680. doi:. http://www.nature.com/nature/journal/v393/n6686/abs/393680a0.html. Summary at Monastersky, R. (20-06-1998). «Getting the scoop from the poop of T. rex». Science News 153 (25): pp. 391. doi:. http://www.sciencenews.org/pages/sn_arc98/6_20_98/fob2.htm.
- ↑ a b Walters, Martin (1995). Bloomsbury Illustrated Dictionary of Prehistoric Life (Bloomsbury Illustrated Dictionaries). Godfrey Cave Associates Ltd. ISBN 1-85471-648-4.
- ↑ Carpenter, K. (1998). «Evidence of predatory behavior by theropod dinosaurs». Gaia 15: pp. 135–144. http://vertpaleo.org/publications/jvp/15-576-591.cfm.
- ↑ Happ, John; and Carpenter, Kenneth (2008). «An analysis of predator-prey behavior in a head-to-head encounter between Tyrannosaurus rex and Triceratops». Tyrannosaurus rex, the Tyrant King (Life of the Past). Bloomington: Indiana University Press. pp. 355–368. ISBN 0-253-35087-5.
- ↑ Tanke, Darren H.; and Currie, Philip J. (1998). «Head-biting behavior in theropod dinosaurs: paleopathological evidence» (pdf). Gaia (15): pp. 167–184. ISSN 0871-5424. http://www.mnhn.ul.pt/geologia/gaia/12.pdf.
- ↑ Farlow, J. O. and T. R. Holtz, Jr. 2002. The fossil record of predation in dinosaurs. pp. 251–266, in M. Kowalewski and P. H. Kelley (eds.), The Fossil Record of Predation. The Paleontological Society Papers 8.
- ↑ a b Longrich N R., Horner J.R., Erickson G.M. & Currie P.J. (2010), "Cannibalism in Tyrannosaurus rex", Public Library of Science.
- ↑ 1999. The teeth of the Tyrannosaurus. Scientific American 281: 40-41.
- ↑ The Complete T. Rex: How Stunning New Discoveries are Changing our Understanding of the World's Most Famous Dinosaur copyright 1993 by John R. Horner: pp 214-215
- ↑ The Complete T. Rex: How Stunning New Discoveries are Changing our Understanding of the World's Most Famous Dinosaur copyright 1993 by John R. Horner: pp 214-215
Bibliografía
- Riehecky, Janet, "Tiranosaurio", Plymouth, MN : Child’s World, c1994.
- Farlow, J. O., Smith, M. B., Robinson, J. M. (1995). Body mass, bone "strength indication," and cursorial potential of Tyrannosaurus rex (Masa corporal, hueso la "indicación de fuerza", y potencial corredor del Tyrannosaurus rex). Journal of Vertebrate Paleontology, 15: 713-725.
- Holtz, T. R. (1994). The phylogenetic position of the Tyrannosauridae: implications for theropod systematics (la posición filogenética de Tyrannosauridae: implicaciones para la sistemática terópoda). Journal of Paleontology 68(5): 1100-1117.
- Horner, John R. and Lessem, Don. The Complete T. Rex. How Stunning New Discoveries Are Changing Our Understanding of the World's Most Famous Dinosaur. (El T. rex completo. Cómo los nuevos estupendos descubrimientos están cambiando nuestra comprensión del dinosaurio más famoso del mundo). Simon and Schuster, 1993. ISBN 0-671-74185-3
- Hutchinson, J. R., García, M. (2002). Tyrannosaurus was not a fast runner (el Tyrannosaurus no era un corredor rápido). Nature 415:1018-1021.
- Osborn, H. F. (1905). Tyrannosaurus and other Cretaceous carnivorous dinosaurs (el Tyrannosaurus y otros dinosaurios carnívoros cretácicos). Bulletin of the American Museum of Natural History 35:733-771.
- Osborn, H. F. (1917). Skeletal adaptations of Ornitholestes, Struthiomimus, Tyrannosaurus (las adaptaciones del esqueleto de Ornitholestes, Struthiomimus, el Tyrannosaurus). Bulletin of the American Museum of Natural History 35: 733-71.
- Schweitzer, M. H., Wittmeyer, J. L., Horner, J. R., Toporski, J. K. (2005). Soft-Tissue Vessels and Cellular Preservation in Tyrannosaurus rex (tejidos suaves de vasos y preservación celular en el Tyrannosaurus rex). Science 5717: 1952-1955.
Véase también
Enlaces externos
 Portal:Dinosaurios. Contenido relacionado con Dinosaurios.
Portal:Dinosaurios. Contenido relacionado con Dinosaurios. Wikimedia Commons alberga contenido multimedia sobre TyrannosaurusCommons.
Wikimedia Commons alberga contenido multimedia sobre TyrannosaurusCommons. Wikiespecies tiene un artículo sobre Tyrannosaurus. Wikispecies
Wikiespecies tiene un artículo sobre Tyrannosaurus. Wikispecies
Información general
- Sobre el T. rex en Duiops.net
- Otros datos interesantes sobre el T. rex
- Abundante información del T. rex en Enchanted Learning
- Excelente diseño e interesantes características del T. rex en Yahooligans!
BBC - Noticias
Especímenes famosos
Otros
Categorías:- Fósiles
- Tyrannosauroidea
- Dinosaurios del Cretácico
- Dinosaurios norteamericanos
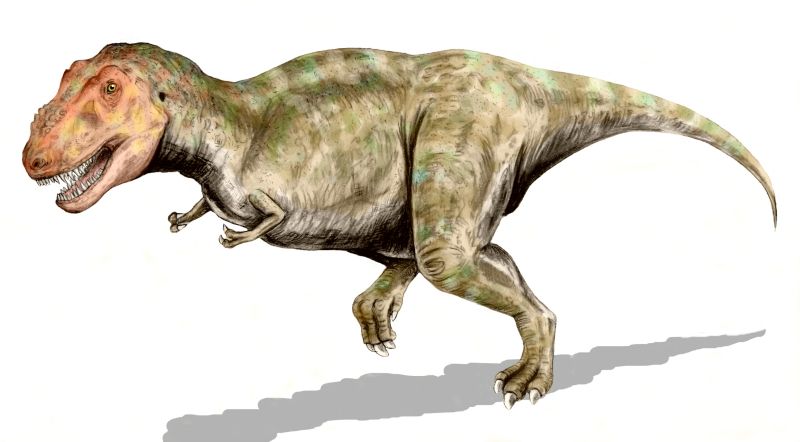
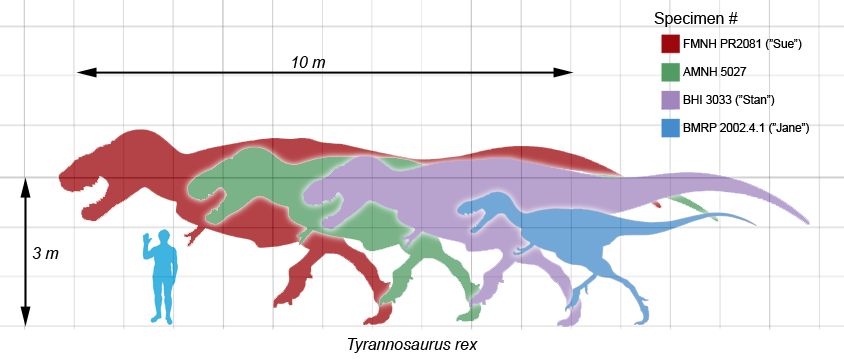


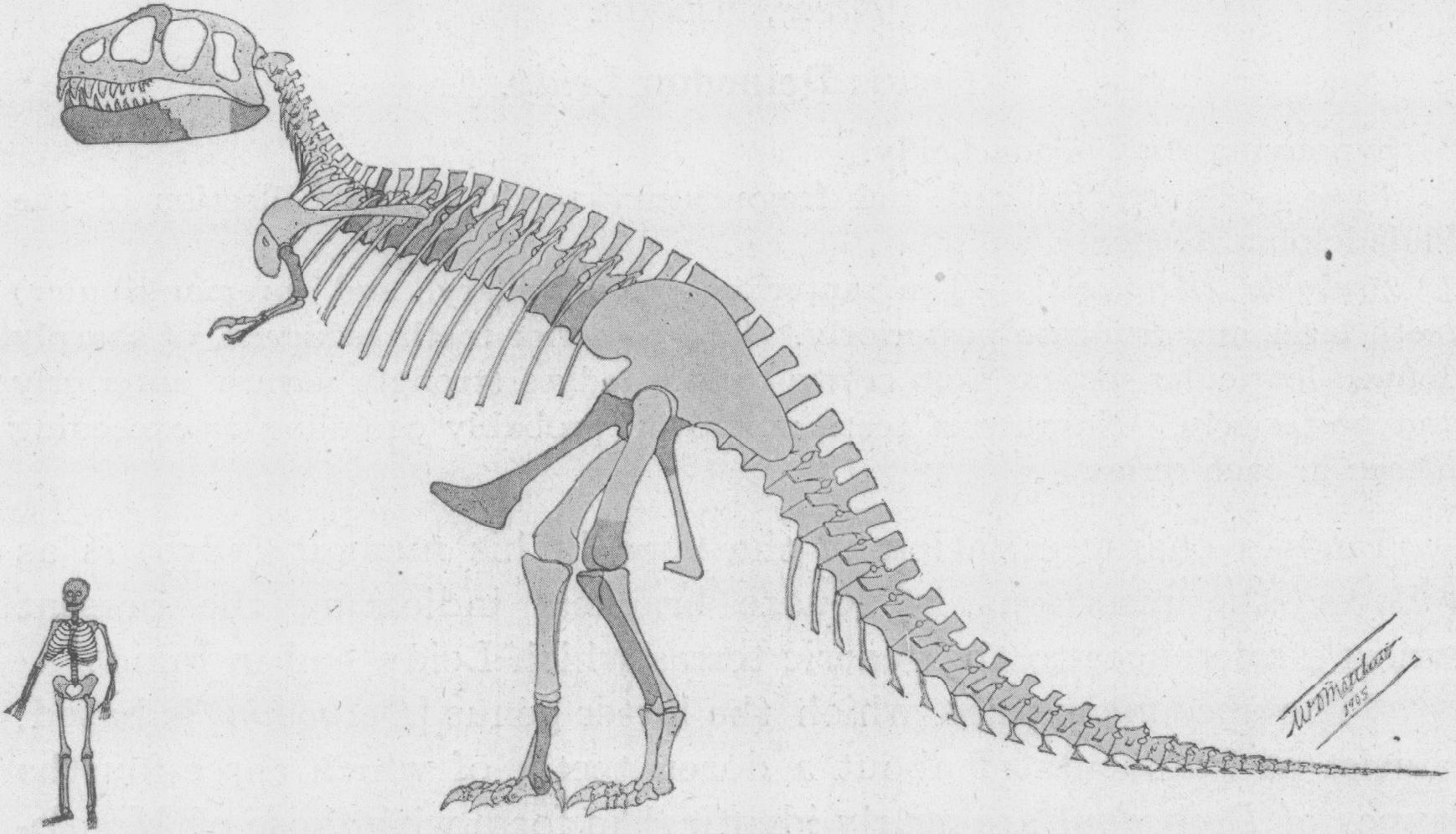




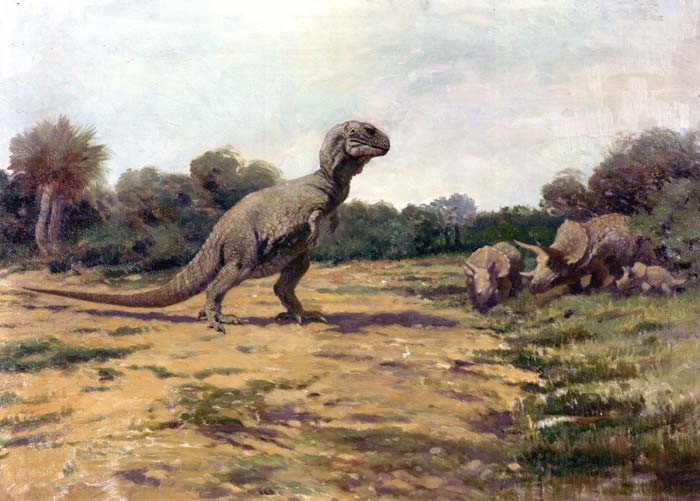

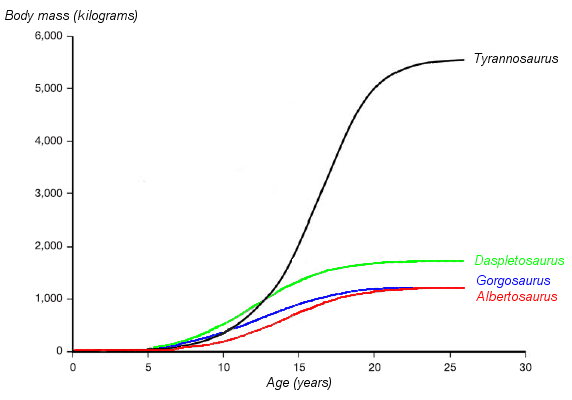








{kind=link}
Wikimedia foundation. 2010.