- Albertosaurus
-
 Albertosaurus
AlbertosaurusRango temporal: Cretácico superior 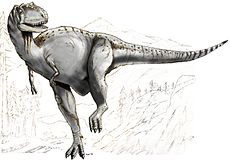
Clasificación científica Reino: Animalia Filo: Chordata Clase: Sauropsida Superorden: Dinosauria Orden: Saurischia Suborden: Theropoda Infraorden: Coelurosauria Familia: Tyrannosauridae Subfamilia: Albertosaurinae Género: Albertosaurus Especie: A. sarcophagus Nombre binomial Albertosaurus sarcophagus
Osborn, 1905Sinonimia A. arctunguis Parks, 1928 Albertosaurus (en griego, "lagarto de Alberta"), nombre común en castellano albertosaurio, es un género de dinosaurio terópodo tiranosáurido que vivió en el oeste de Norteamérica a finales del período Cretácico, hace más de 70 millones de años. La especie tipo A. sarcophagus estaba restringida en el rango de lo que hoy es la provincia canadiense de Alberta, a la cual hace referencia el nombre del género. Existe cierto desacuerdo en la comunidad científica en cuanto al número de especies representadas en el género, que según el caso serían una o dos.
Al ser un tiranosáurido, los albertosaurios fueron depredadores bípedos con una gran cabeza, mandíbulas provistas de docenas de grandes dientes y pequeñas "manos" de dos dedos. Pudo haber sido el mayor depredador en la cadena alimenticia de su ecosistema local. Aunque sea relativamente grande para un terópodo, Albertosaurus era mucho más pequeño que su famoso pariente Tyrannosaurus, probablemente pesando tanto como un rinoceronte negro moderno.
Han sido descubiertos fósiles de más de 20 individuos, permitiendo a los científicos un conocimiento más detallado de la anatomía de Albertosaurus que la que se tiene de otros tiranosáuridos. El hallazgo de diez individuos juntos en un sitio, señala evidencia de comportamiento en grupos y permite estudios de su biología del desarrollo, imposibles con animales menos conocidos.
Contenido
Descripción
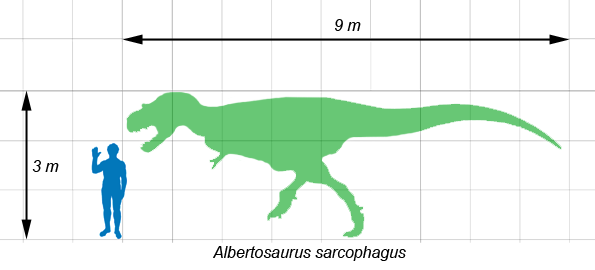
Albertosaurus era de menor tamaño que los gigantescos tiranosáuridos más conocidos como Tarbosaurus y Tyrannosaurus. Los adultos alcanzaban aproximadamente 9 metros de largo,[1] [2] mientras que algunos individuos, los de mayor edad, habrían alcanzado los 10 metros.[3] Varias estimaciones independientes sobre su masa, obtenidas mediante diferentes métodos, sugieren que un Albertosaurus adulto habría pesado entre 1,3[4] y 1,7 toneladas.[5]
 Mandíbulas parciales de un Albertosaurus, expuestas en el Museo Real de Ontario.
Mandíbulas parciales de un Albertosaurus, expuestas en el Museo Real de Ontario.
El enorme cráneo del albertosaurio, sostenido por un cuello corto con forma de S, medía aproximadamente 1 metro de largo en los adultos más grandes.[6] Poseía unas amplias aberturas denominadas fenestras temporales, que reducían el peso de la cabeza, y proporcionaban áreas para la inserción de músculos y órganos sensoriales. Sus largas mandíbulas contenían más de 60 dientes con forma de banana, muchos en comparación a los tiranosáuridos más grandes que poseían menos dientes. A diferencia de la mayoría de terópodos, los tiranosáuridos fueron heterodontes, es decir los dientes tomaron diferentes formas dependiendo de su posición en la boca. Los dientes premaxilares en el extremo de la mandíbula superior eran mucho más pequeños que el resto, compactos y con forma de D en una sección de corte.[2]
Todos los tiranosáuridos, incluyendo a Albertosaurus, compartían una apariencia corporal similar típica para un terópodo. Albertosaurus era bípedo y balanceaba su pesada cabeza y tronco con una larga cola. Sin embargo, los miembros delanteros de los tiranosáuridos eran extremadamente pequeños para su tamaño y tenían solamente dos dedos. Los miembros traseros eran largos y terminaban en un pie de cuatro dedos. El primero de estos dedos era muy pequeño y se ubicaba en la parte posterior y sólo los otros tres se apoyaban en el suelo, con el dedo medio más largo que los demás.[2] Albertosaurus puede haber podido alcanzar velocidades de 11 a 13 metros por segundo (40 a 48 km/h).[7]
Historia
El primer albertosaurio fue encontrado por Joseph Burr Tyrrell durante unas prospecciones geológicas en 1884. El albertosaurio fue nombrado en 1905 por Henry Fairfield Osborn, del Museo Americano de Historia Natural, en una muy breve nota al final de su descripción de Tyrannosaurus rex. El nombre hace honor a Alberta, la provincia canadiense en la cual se encontraron los primeros restos. El nombre genérico también incorpora el término griego σαύρος (sauros, que significa "lagarto"), el sufijo más común en nombres de dinosaurio.[8]
 Un cráneo de albertosáurino descubierto en Nuevo México, durante su preparación. Esta especie aún no descrita puede pertenecer al Albertosaurus.
Un cráneo de albertosáurino descubierto en Nuevo México, durante su preparación. Esta especie aún no descrita puede pertenecer al Albertosaurus.La especie tipo de Albertosaurus es A. sarcophagus, también nombrada por Osborn en 1905. El nombre significa "comedor de carne" y tiene la misma etimología que el contenedor funerario sarcófago, con el cual comparte su nombre: una combinación de las palabras del griego antiguo σάρξ (sarx, que significa "carne") y φάγειν (fagein, que significa "comer").[8] Se conocen más de 20 especímenes de diversas edades.[9]
Primeros descubrimientos
El espécimen tipo es un cráneo parcial, recolectado en 1884 de un afloramiento junto al río Red Deer en Alberta. Este espécimen, junto con un cráneo adicional más pequeño y algo más de material esquelético, fue descubierto por una expedición de la Comisión Geológica de Canadá, dirigida por el famoso geólogo canadiense Joseph B. Tyrell. Este espécimen es ahora guardado en el Museo Canadiense de Naturaleza. Los dos cráneos fueron asignados a la especie Laelaps incrassatus por Edward Drinker Cope en 1892.[10] Sin embargo, en 1877 el nombre Laelaps ya había sido asignado a un género de ácaro, por lo que fue sustituido y rebautizado como Dryptosaurus por el rival de Cope, Othniel Charles Marsh (Cope rechazó aceptar el nuevo nombre). Lawrence Lambe desplazó Laelaps incrassatus al género Dryptosaurus en 1904.[11] Finalmente, puesto que el D. incrassatus se basaba solamente en dientes genéricos de tiranosáurido que no podían indicar fácilmente su asignación a una especie en particular, y que el material del cráneo de Alberta se diferenciaba notablemente del Dryptosaurus, Osborn lo nombró como Albertosaurus sarcophagus en 1905.[8]
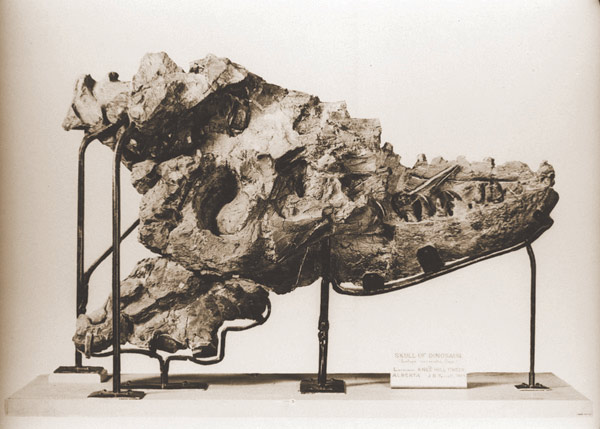
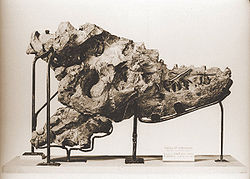 Cráneo parcial del primer Albertosaurio descubierto por Joseph Burr Tyrrell.
Cráneo parcial del primer Albertosaurio descubierto por Joseph Burr Tyrrell.La cama de Huesos de Dry Island
En 1910, el paleontólogo estadounidense Barnum Brown desenterró los restos de un gran grupo de Albertosaurus en otra cantera junto al río Red Deer. A causa del gran número de huesos y del limitado tiempo disponible, el equipo de Brown no alcanzó a recolectar cada espécimen, pero se aseguró de recolectar los huesos de todos los individuos presentes. Entre muchos otros huesos depositados en las colecciones del Museo Americano de Historia Natural en Nueva York, caben destacar siete series de metatarsos derechos, junto con dos huesos aislados del dedo de la pata que no encajan con ninguno de los metatarsos en tamaño. Esto indica la presencia de por lo menos 9 individuos en la cantera. El Museo Real Tyrrell de Paleontología redescubrió el sitio en 1997 y reanudó el trabajo de campo.[12] Posteriormente, esta excavación logró hallar un décimo individuo muy joven en 2002.[4]
Un espécimen originalmente nombrado como A. arctunguis fue también hallado cerca del río Red Deer y se encuentra depositado actualmente en el Museo Real de Ontario en Toronto, Canadá.[13] Seis cráneos y esqueletos más han sido hallados desde entonces en Alberta y depositados en otros museos canadienses.[9]
Todos los fósiles identificables de Albertosaurus sarcophagus se conocen de la Formación del Cañón de Herradura en Alberta. Esta formación data de comienzos de la época Mastrichtiana del Cretácico superior, unos 70 a 73 millones de años atrás. Muchos otros dinosaurios han sido encontrados ahí, incluyendo terópodos más pequeños como Ornithomimus, Chirostenotes y varios dromeosáuridos, y con una amplia variedad de herbívoros como anquilosaurios, ceratopsianos, paquicefalosaurios y hadrosáuridos. Fósiles de Albertosaurus han sido también reportados de los estados estadounidenses de Montana, Nuevo México, y Wyoming, pero estos probablemente no representan al A. sarcophagus y pueden no pertenecer al género Albertosaurus.[2] [9]
Clasificación
Albertosaurus es un miembro de la familia de terópodos Tyrannosauridae. Dentro de esta familia, Albertosaurus sarcophagus es normalmente clasificado con el levemente más antiguo Gorgosaurus libratus (a veces llamado Albertosaurus libratus) en la subfamilia Albertosaurinae.[14] Al Appalachiosaurus se lo ha ubicado como albertosáurino en al menos un estudio de Thomas Holtz en 2004,[2] aunque esto es discutido.[15] En un trabajo inédito más reciente Holtz lo localiza apenas por afuera de Tyrannosauridae,[16] de común acuerdo con otros autores.[17] El cladograma que refleja las relaciones descriptas es el siguiente:
Tyrannosauridae Tyrannosaurinae
Albertosaurinae Albertosaurus
Los albertosáurinos eran más delgados que los robustos tiranosáurinos, como tarbosaurio, tiranosaurio y daspletosaurio, con cráneos proporcionalmente más pequeños y huesos más largos de la pierna (tibia) y pies (metatarsos y falanges).[6] [18]
Gorgosaurus libratus
En 1913, el paleontólogo Charles Hazelius Sternberg descubrió otro esqueleto de tiranosáurido en unos sedimentos algo más antiguos situados en Alberta. Este dinosaurio fue nombrado como Gorgosaurus libratus en 1914 por Lawrence Lambe.[19] Al haber hallado unas pocas diferencias para separar los dos géneros, Dale Russell declaró al Gorgosaurus un sinónimo de Albertosaurus en 1970, creando la nueva combinación Albertosaurus libratus. Este hecho dio lugar a la ampliación del rango temporal (varios millones de años atrás) y del rango geográfico (cientos de km hacia el Sur) de este género.[1]
Un examen más reciente de Albertosaurus y de Gorgosaurus ha puesto en duda la sinonimia propuesta por Russell. En 2003, Phil Currie y sus colegas examinaron los cráneos de las dos especies y llegaron a la conclusión de que ambos géneros deben mantenerse, aunque reconocieron que los dos géneros son taxones hermanos y que la distinción es por consiguiente muy arbitraria. Sin embargo, de acuerdo con Currie, Albertosaurus y Gorgosaurus no son más similares que lo que lo son Daspletosaurus y Tyrannosaurus, casi siempre considerados géneros independientes. Además, varios especímenes sin describir de albertosáurinos han sido hallados en otras partes de Norteamérica, incluyendo Alaska y Nuevo México, así que Currie ha recomendado dejar a estos dos géneros separados hasta obtener más información al respecto.[9] Muchos autores desde entonces han seguido la recomendación de Currie,[2] [4] [20] pero otros no.[15]
Especies inválidas
Han sido nombradas algunas otras especies de Albertosaurus que posteriormente han resultado ser inválidas. William Parks describió un esqueleto parcial hallado en Alberta como Albertosaurus arctunguis en 1928,[13] pero actualmente es considerado un sinónimo de A. sarcophagus.[1] El espécimen de Parks (ROM 807) se mantiene en el Museo Real de Ontario en Toronto. La especie Albertosaurus megagracilis (después renombrada como Dinotyrannus[21] ) se basó en el esqueleto de un pequeño tiranosáurido de la Formación de Hell Creek en Montana (Estados Unidos), pero actualmente se cree que fue un juvenil de Tyrannosaurus.[22] [6]
Al menos seis cráneos y esqueletos más se han descubierto en Alberta y se guardan desde entonces en varios museos canadienses. Fósiles también se han encontrado de los estados americanos de Montana, de Nuevo México, y Wyoming, pero éstos no representan a A. sarcophagus y pueden probablemente incluso no pertenecer a Albertosaurus.[2] [9]
Paleobiología
Como muchos otros tiranosáuridos, Albertosaurus ha sido estudiado exhaustivamente a través de diversos especímenes diferentes, permitiendo estudios detallados sobre su vida y proporcionando evidencias sobre su comportamiento.
Patrones de crecimiento
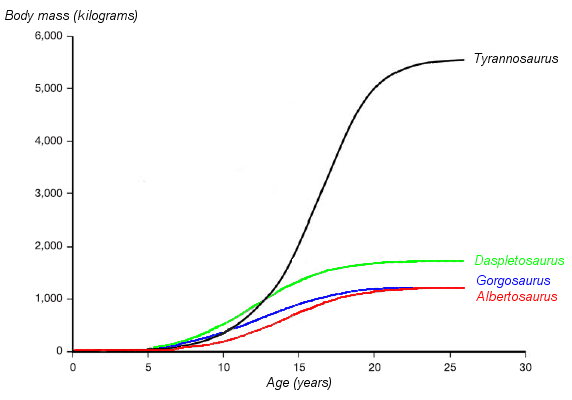
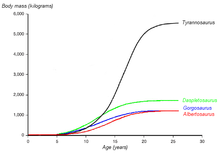 Un gráfico mostrando las curvas hipotéticas de crecimiento (masa corporal versus edad) de cuatro tiranosáuridos. La del Albertosaurus está dibujada en rojo. Basado en Erickson et al. 2004.
Un gráfico mostrando las curvas hipotéticas de crecimiento (masa corporal versus edad) de cuatro tiranosáuridos. La del Albertosaurus está dibujada en rojo. Basado en Erickson et al. 2004.Casi la totalidad de las edades de Albertosaurus están representadas en el registro fósil. Por medio de estudios histológicos de los huesos se puede determinar casi siempre la edad de un individuo en el momento de su muerte, permitiéndonos así estimar los ritmos de crecimiento y compararlos con los de otras especies. Recientemente ha sido hallado un fósil de Albertosaurus de dos años de edad, el más joven hallado hasta el momento, en la cama de huesos de Dry Island que ha sido re-abierta, que debió pesar cerca de 50 kilogramos y medir 2 metros de largo. Otro espécimen de la misma cantera es el más viejo, teniendo alrededor de 28 años de edad y unos 10 metros de largo. Un espécimen de 24 años de edad de la colección del Museo Tyrrell de Paleontología habría pesado aproximadamente 1,14 toneladas. Sin embargo, otro individuo de 1,28 toneladas, que se encuentra en la colección del Museo Americano de Historia Natural tenía 22 años de edad cuando murió. Cuando se estudian especímenes de edad y tamaño intermedios, representándose su curva de crecimiento, obtenemos una gráfica en forma de S, donde la mayor tasa de crecimiento se produce en un período de 4 años, que finaliza alrededor de los 16 años, como se observa en otros tiranosáuridos. La tasa de crecimiento durante esta fase se estima en un aumento de 122 kg por año. Otros tiranosáuridos de tamaño similar muestran un ritmo de crecimiento parecido, pero el ritmo es mucho más bajo que el de Tyrannosaurus, el cual crecía casi ocho veces más rápido (601 kg por año) durante esta fase de crecimiento.[3] Albertosaurus parece alcanzar la madurez esquelética, a los 16 años de edad, la misma está marcada por el fin de la fase de rápido crecimiento, que al parecer también era la edad donde alcanzaba su madurez sexual, sin embargo el crecimiento continuaba a un ritmo lento a lo largo de la vida de los animales.[4] [3] El alcanzar la madurez sexual cuando el crecimiento todavía es activo parece ser una característica compartida entre dinosaurios pequeños[23] y grandes[24] así como en mamíferos grandes tales como seres humanos y elefantes.[24] Este patrón de maduración sexual relativamente temprana se diferencia llamativamente del patrón en los pájaros, que retrasan su madurez sexual hasta después de que hayan acabado el crecimiento.[20] [24]
Biología del desarrollo
La mayoría de los albertosaurios conocidos tenían alrededor de catorce años o más al momento de su muerte. Los animales juveniles rara vez son encontrados fosilizados, principalmente por un caso de muestreo sesgado, donde es menos probable que los huesos más pequeños de los animales más jóvenes sean preservado por la fosilización que los huesos más grandes de adultos, y que los huesos pequeños son más difíciles de ser notados por los recolectores en el campo.[25] Aún así los albertosaurios jóvenes son relativamente grandes para ser animales juveniles, pero sus restos son raros en el registro fósil comparado con los adultos. Se ha sugerido que este fenómeno es una consecuencia de la biología del desarrollo y que los fósiles de albertosaurios jóvenes sean raros porque simplemente no murieron tan a menudo como los adultos.[3]
Una hipótesis postula que luego de una eclosión masiva los pequeños albertosarios murieron en grandes números, pero no se ha preservado en el registro fósil debido a su pequeño tamaño y fragilidad. Después de apenas dos años, los jóvenes eran más grandes que cualquier otro depredador en la región aparte del albertosaurio adulto, y más veloces que la mayoría de sus presas. Esto dio lugar a una disminución dramática de su tasa de mortalidad y sigue habiendo una rareza correspondiente en los fósiles. Las tasas de mortalidad se duplican al llegar a los doce años, quizás como resultado de las demandas fisiológicas de la fase de crecimiento rápido, y después se vuelve a duplicar otra vez con el inicio de la madurez sexual entre las edades de catorce y dieciséis años. Esta tasa de mortalidad alta continúa a través de edad adulta, quizás debido a las altas demandas fisiológicas, tensión y lesiones recibidas durante la competición intraespecífica para los compañeros y recursos, y eventual, los efectos cada vez mayores de la senescencia.
La tasa de mortalidad más alta en adultos puede explicar su preservación más común. Los animales muy grandes eran raros porque pocos individuos sobrevivieron bastante tiempo para lograr tales tamaños. Las altas tasas de mortalidad infantil, seguidas por mortalidad reducida entre jóvenes y un aumento repentino en mortalidad después de madurez sexual, con muy pocos animales alcanzando tamaño máximo, son un patrón observado en muchos mamíferos grandes modernos, incluyendo elefantes, búfalo africano, y rinoceronte. El mismo patrón también se considera en otros tiranosáuridos. Como conclusión y comparando con los animales modernos y lo que se sabe de otros tiranosáuridos parece apoyar a esta hipótesis anteriormente detallada de biología del desarrollo, pero el sesgo en el registro fósil pueden todavía desempeñar un papel importante, especialmente a partir de que más de dos tercios de todo los especímenes de albertosaurios se conocen a partir de un lugar.[3] [20]
Comportamiento en manada
 Modelos de albertosaurio, Museo Tyrrell.
Modelos de albertosaurio, Museo Tyrrell.La capa de huesos del Albertosaurus descubierta por Barnum Brown y su equipo contiene los restos de al menos 10 individuos. Se observa que este grupo estaba compuesto por dos o tres individuos totalmente crecidos y de unos 21 años de edad; un adulto más joven de 17 años; cuatro sub-adultos en plena fase de crecimiento rápido entre los 12 y los 16 años; y un juvenil de 10 años, el cual no alcanzó la fase de crecimiento. También se halló un individuo muy joven, de unos dos años de edad.[4]
La ausencia de restos de herbívoros en las inmediaciones y el estado similar de preservación entre los muchos individuos de la cantera de capa de huesos de Albertosaurus llevó a Currie a la conclusión de que la localidad no fue una "trampa de depredadores" como Rancho La Brea en California, y que todos los animales hallados murieron al mismo tiempo, proporcionando evidencias de un comportamiento en manada.[12] Otros científicos son escépticos al respecto, señalando que los animales pudieron haberse agrupado por una inundación u otras causas.[26]
Existen numerosas evidencias de un comportamiento gregario entre dinosaurios herbívoros, incluyendo ceratopsianos y hadrosaurios.[27] Por el contrario, solo en raras ocasiones se han hallado varios dinosaurios depredadores en un mismo sitio. Pequeños terópodos como Coelophysis bauri, Deinonychus antirrhopus y Megapnosaurus rhodesiensis han sido encontrados en grupos, cuando hay depredadores más grandes como Allosaurus fragilis o Mapusaurus roseae.[28] [29] [30] También existen ciertas evidencias de comportamiento gregario en otros tiranosáuridos. Restos fragmentarios de individuos más pequeños fueron encontrados junto a "Sue", Tyrannosaurus rex montado en el Museo Field de Historia Natural en Chicago. Una capa de huesos en la Formación Two Medicine de Montana contenía al menos tres especímenes de una especie no nombrada de Daspletosaurus, preservados junto a varios hadrosaurios.[31] Estos hallazgos pueden corroborar la evidencia de un comportamiento social en Albertosaurus, aunque algunos o todos los lugares ya mencionados pueden representar agregaciones temporales o no naturales. Otros han especulado que en vez de grupos sociales, por lo menos algunos de estos hallazgos representan caso de frenesí alimentario similar a los del dragones de Cómodo sobre reses muertas, donde los carnívoros compiten agresivamente, pudiendo morir alguno en las luchas sobre el cual el resto practica el canibalismo.[25]
Currie también realizó algunas especulaciones acerca de los hábitos de caza en grupo de Albertosaurus. Las proporciones de las extremidades inferiores de los individuos más pequeños son comparables a las de los ornitomímidos, los cuales estuvieron probablemente entre los dinosaurios más veloces. El albertosaurio más joven era probablemente tan rápido como su presa, entre las que se podrían incluir ceratopsianos y hadrosaurios. Currie sugirió que los miembros más jóvenes de la manada podrían ser los responsables de conducir la presa hacia los adultos, que eran más grandes y fuertes, pero también más lentos.[12] Los jóvenes debieron tener una forma de vida distinta a la de los adultos, ocupando el espacio como depredador entre los enormes adultos y los terópodos contemporáneos más pequeños, el mayor de ellos tenía una masa dos veces menor que Albertosaurus.[2] Una situación similar se observa en los dragones de Komodo modernos, con los jóvenes comenzando su vida como pequeños insectívoros para luego convertirse en los depredadores dominantes en sus islas.[32] Sin embargo, como la preservación del comportamiento en el registro fósil es extremadamente rara, esta idea no puede ser probada fácilmente.
Paleoecología
 La Formación Cañón Horseshoe queda expuesta en esta localidad tipo del Cañón de la Herradura de Alberta.
La Formación Cañón Horseshoe queda expuesta en esta localidad tipo del Cañón de la Herradura de Alberta.Todos los fósiles identificables como A. sarcophagus son conocidos de Formación Cañón Horseshoe en Alberta. Esta formación geológica datada entre el Mastrichtiano temprano durante el periodo Cretácico superior, hace aproximadamente 73 a 70 millones de años. Inmediatamente debajo de esta formación está la pizarra de Bearpaw, a una formación de origen marino que representa una sección del vía marítima interior occidental. La vía marítima retrocedió cuando la temperatura promedio del año retrocedio a finales del Cretácico haciéndose más frío y los niveles del mar disminuyeron a final del cretáceo, exponiendo la tierra que había sido previamente subacuática. No era un proceso parejo, y la vía marítima ingresaría periódicamente para cubrir las partes de la región a través del Cañón Horseshoe antes de finalmente de retroceder en conjunto en los años después.
Debido a los niveles cambiantes del mar, muchos diversos ambientes se representan en la Formación Cañón Herradura, incluyendo ambientes de mar adentro y costeros, como lagunas, estuarios y llanuras de mareas así como numerosas vetas de carbón que representan una antigua turba pantanosa. Como la mayor parte de los vertebrados fósiles de la formación, los restos de Albertosaurus se encuentran en los depósitos colocados en deltas y terrenos inundables de los grandes ríos durante la segunda mitad de la formación. [33]
La fauna de Formación Cañón Horseshoe es bien conocida, contiene vertebrados fósiles, incluyendo comúnmente dinosaurios entre estos. Tiburones, rayas, esturiones, Amiiformes, Lepisosteiformes y el símil Lepisosteiformes Aspidorhynchus formando parte de la fauna ictícola. Los mamíferos incluyen a multituberculados y el marsupial Didelphodon. El plesiosaurio marino Leurospondylus se ha encontrado en rocas de la Formación Cañón Horseshoe, con los ambientes de agua dulce donde abundaban tortugas, Champsosaurus y crocodilos como Leidyosuchus y Stangerochampsa. Los dinosaurios dominaban la fauna, especialmente hadrosáuridos, cuáles componen la mitad de todos los dinosaurios conocidos, incluyendo los géneros Edmontosaurus, Saurolophus y Hypacrosaurus. Ceratopsianos u ornitomímidos eran también comunes, junto componiendo otra tercera parte de la fauna. Junto con los mucho más raros anquilosáuridos y paquicefalosáuridos, todos estos animales habrían sido presa para una variedad diversa de terópodos carnívoros, incluyendo troodóntidos, dromeosáuridos, y cenagnátidos. Los albertosaurios adultos son los principales predadores en este ambiente, con los juveniles posiblemente ocupando nichos intermedios.[33]
Véase también
- Anexo: Géneros válidos de dinosaurios
 Portal:Dinosaurios. Contenido relacionado con Dinosaurios.
Portal:Dinosaurios. Contenido relacionado con Dinosaurios.
Referencias
- ↑ a b c Russell, D.A. 1970. Tyrannosaurs from the Late Cretaceous of western Canada. National Museum of Natural Sciences Publications in Paleontology 1: 1-34.
- ↑ a b c d e f g h Holtz, T.R. 2004. Tyrannosauroidea. In: Weishampel, D.B., Dodson, P. & Osmolska, H. (Eds.). The Dinosauria (2nd Edition). Berkeley: University of California Press. Pp. 111-136.
- ↑ a b c d e Erickson, Gregory M.; Currie, Philip. J.; Inouye, Brian D.; & Wynn, Alice A. (2006). «Tyrannosaur life tables: an example of nonavian dinosaur population biology». Science 313: pp. 213–217. doi:. PMID 16840697.
- ↑ a b c d e Erickson, G.M., Makovicky, P.J., Currie, P.J., Norell, M.A., Yerby, S.A., & Brochu, C.A. 2004. Gigantism and comparative life-history parameters of tyrannosaurid dinosaurs. Nature 430: 772-775.
- ↑ Christiansen, P. & Fariña, R.A. Mass prediction in theropod dinosaurs. Historical Biology 16: 85-92.
- ↑ a b c Currie, P.J. 2003a. Allometric growth in tyrannosaurids (Dinosauria: Theropoda) from the Upper Cretaceous of North America and Asia. Canadian Journal of Earth Sciences 40: 651-665.
- ↑ "Albertosaurus." In: Dodson, Peter & Britt, Brooks & Carpenter, Kenneth & Forster, Catherine A. & Gillette, David D. & Norell, Mark A. & Olshevsky, George & Parrish, J. Michael & Weishampel, David B. The Age of Dinosaurs. Publications International, LTD. p. 106-107. ISBN 0-7853-0443-6.
- ↑ a b c Osborn, H.F. 1905. Tyrannosaurus and other Cretaceous carnivorous dinosaurs. Bulletin of the American Museum of Natural History. 21: 259-265.
- ↑ a b c d e Currie, P.J. 2003b. Cranial anatomy of tyrannosaurids from the Late Cretaceous of Alberta. Acta Palaeontologica Polonica 48(2): 191-226.
- ↑ Cope, E.D. On the skull of the dinosaurian Laelaps incrassatus Cope. American Philosophical Society, Proceedings 30: 240-245.
- ↑ Lambe, L.M. 1904. On Dryptosaurus incrassatus (Cope) from the Edmonton Series of the Northwest Territory. Contributions to Canadian Palaeontology 3: 1-27.
- ↑ a b c Currie, P.J. 1998. Possible evidence of gregarious behavior in tyrannosaurids. Gaia 15: 271-277. [not printed until 2000]
- ↑ a b Parks, W.A. 1928. Albertosaurus arctunguis, a new species of therapodous dinosaur from the Edmonton Formation of Alberta. University of Toronto Studies, Geological Series 25: 1-42.
- ↑ Currie, P.J., Hurum, J.H. & Sabath, K. 2003. Skull structure and evolution in tyrannosaurid phylogeny. Acta Palaeontologica Polonica 48(2): 227-234.
- ↑ a b Carr, T.D., Williamson, T.E. & Schwimmer, D.R. 2005) A new genus and species of tyrannosaurid from the Late Cretaceous (middle Campanian) Demopolis Formation of Alabama. Journal of Vertebrate Paleontology 25(1): 119-143.
- ↑ Holtz, Thomas R. (20-09-2005). «RE: Burpee Conference (LONG)». Consultado el 18-06-2007.
- ↑ Carr, Thomas D.; Williamson, Thomas E.; & Schwimmer, David R. (2005). «A new genus and species of tyrannosauroid from the Late Cretaceous (middle Campanian) Demopolis Formation of Alabama». Journal of Vertebrate Paleontology 25 (1): pp. 119–143. doi:.
- ↑ Currie, Philip J.; Hurum, Jørn H; & Sabath, Karol. (2003). "Skull structure and evolution in tyrannosaurid phylogeny". Acta Palaeontologica Polonica 48 (2): 227–234.
- ↑ Lambe, L.M. 1914. On a new genus and species of carnivorous dinosaur from the Belly River Formation of Alberta, with a description of the skull of Stephanosaurus marginatus from the same horizon. Ottawa Naturalist 28: 13-20.
- ↑ a b c Ricklefs, Robert E. (2007). «Tyrannosaur ageing». Biology Letters 3 (2): pp. 214–217. doi:.
- ↑ Olshevsky, G. 1995. [The origin and evolution of the tyrannosaurids.] Kyoryugaku Saizensen [Dino Frontline] 9: 92–119. (en japonés)
- ↑ Paul, G.S. Predatory Dinosaurs of the World. New York: Simon & Schuster. 464pp.
- ↑ Erickson, Gregory M.; Curry Rogers, Kristi; Varricchio, David J.; Norell, Mark.; & Xu, Xing (2007). «Growth patterns in brooding dinosaurs reveals the timing of sexual maturity in non-avian dinosaurs and génesis of the avian condition». Biology Letters 3: pp. 558–561. doi:.
- ↑ a b c Lee, Andrew H.; Werning, Sarah (2008). «Sexual maturity in growing dinosaurs does not fit reptilian growth models». PNAS 105: pp. 582–587. doi:. PMID 18195356.
- ↑ a b Roach, Brian T.; & Brinkman, Daniel T. (2007). «A reevaluation of cooperative pack hunting and gregariousness in Deinonychus antirrhopus and other nonavian theropod dinosaurs». Bulletin of the Peabody Museum of Natural History 48 (1): pp. 103–138. doi:. http://www.peabody.yale.edu/scipubs/abstracts/abs_b48-1b.html.
- ↑ Eberth, D.A. & McCrea, R.T. 2001. Were large theropods gregarious? Journal of Vertebrate Paleontology 21(3): 46A. [abstract publicado]
- ↑ Horner, J.R. 1997. Behavior. En: Currie, P.J. & Padian, K.P. (Eds.). Encyclopedia of Dinosaurs Berkeley: University of California Press, pp. 45-50
- ↑ Maxwell, W.D. & Ostrom, J.H. 1995. Taphonomy and paleobiological implications of Tenontosaurus-Deinonychus associations. Journal of Vertebrate Paleontology. 15(4): 707-712.
- ↑ Raath, M.A. Morphological variation in small theropods and its meaning in systematics: evidence from Syntarsus rhodesiensis. En: Carpenter, K. & Currie, P.J. Dinosaur Systematics: Approaches and Perspectives. Cambridge: Cambridge University Press. Pp. 91-105.
- ↑ Coria, R.A. & Currie, P.J. 2006. A new carcharodontosaurid (Dinosauria, Theropoda) from the Upper Cretaceous of Argentina. Geodiversitas. 28(1): 71-118.
- ↑ Currie, P.J., Trexler, D., Koppelhus, E.B., Wicks, K., & Murphy, N. 2005. An unusual multi-individual tyrannosaurid bonebed in the Two Medicine Formation (Late Cretaceous, Campanian) of Montana (USA). En: Carpenter, K. (Ed.). The Carnivorous Dinosaurs. Bloomington: Indiana University Press, pp. 313-324.
- ↑ Auffenberg, Walter (2000). The Behavioral Ecology of the Komodo Monitor. Gainesville: University Press of Florida. pp. 460pp.. ISBN 978-0813006215.
- ↑ a b Eberth, David A. (1997). «Edmonton Group». En Currie, Philip J. & Padian, Kevin (eds.).. Encyclopedia of Dinosaurs. San Diego: Académic Press. pp. 199-204. ISBN 0-12-226810-5.
Enlaces externos
 Wikimedia Commons alberga contenido multimedia sobre AlbertosaurusCommons.
Wikimedia Commons alberga contenido multimedia sobre AlbertosaurusCommons. Wikiespecies tiene un artículo sobre Albertosaurus. Wikispecies
Wikiespecies tiene un artículo sobre Albertosaurus. Wikispecies- Sobre el Albertosaurus en Duiops.net
- Una breve historia de Joseph B. Tyrrell y su descubrimiento del cráneo holotipo del Albertosaurus sarcophagus (en inglés)
- Operation Dino-Lift, detallando el descubrimiento, excavación, y preparación de un posible Albertosaurus de tierras de Bureau of Land Management en Nuevo México (en inglés)
- Albertosaurus en Dinodata (en inglés, se requiere registrarse en forma gratuita)
- Albertosaurus dentro de Tyrannosauroidea en The Thescelsosaurus (en inglés)
- Ficha e imagen de Albertosaurus en Dinosaurier-web (en alemán)
- Significado y pronunciación del nombre (en inglés)
- Albertosaurus en The Theropod Database (en inglés)
Categorías:- Fósiles
- Tyrannosauroidea
- Dinosaurios del Cretácico
- Dinosaurios norteamericanos




Wikimedia foundation. 2010.