- Desarrollo de las extremidades
-
Desarrollo de las extremidades
 Embrión humano de seis semanas
Embrión humano de seis semanas
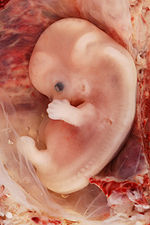 Embrión humano de nueve semanas procedente de un embarazo ectópico.
Embrión humano de nueve semanas procedente de un embarazo ectópico.El desarrollo de las extremidades de tetrápodos (animales con cuatro extremidades) es un área de investigación correspondiente a la biología del desarrollo. La formación del miembro comienza como un esbozo de la extremidad. El factor de crecimiento de fibroblastos (FGF) induce la formación de un organizador llamdo cresta ectodérmica apical (AER, por sus siglas en inglés), que guía el posterior desarrollo y controla la muerte celular. La apoptosis — muerte celular programada — es necesaria para eliminar las membranas interdigitales.
El campo de la extremidad es una región especificada por la expresilón de un gen homeobox (Hox), el cual parece ser controlado por ácido retinoico.[1]
La formación de las extremidades es el resultado de una serie de inducciones epitelio-mesenquimatosas entre las células mesenquimáticas del esbozo de la extremidad y las células ectodérmicas que se sitúan sobre estas. Las células del mesodermo de la placa lateral y el miotomo migran hacia el campo de la extremidad y proliferan para formar el esbozo de la extremidad. La placa lateral produce las porciones cartilaginosas y esqueléticas de la extremidad, en tanto que las células del miotomo producen los componentes musculares. Las células mesodérmicas de la placa lateral secretan un factor de crecimiento de fibroblastos (posiblemente FGF7 y FGF10) para inducir al ectodermo que está situado sobre el a formar una importante estructura llamada cresta ectodérmica apical (AER).[2] La AER recíprocamente secreta FGF8 y FGF4 los cuales mantienen la señalización por FGF10 e inducen la proliferación en el mesodermo. La posición de la expresión de FGF10 está regulada por Wnt8c en el miembro anterior, y por Wnt2b en el posterior. Ambos miembros están especificados por su posición sobre el eje posterior y posiblemente por dos factores de transcripción con T-box: Tbx5 y Tbx4, respectivamente.[3] [4]
La muerte celular programada elimina la membrana interdigital entre los dedos y las articulaciones. La señalización por las proteínas BMP induce la muerte celular y la proteína Noggin bloquea la muerte celular entre los dedos.
Contenido
Condensaciones precartilaginosas
Los elementos esqueléticos de la extremidad están preconfigurados en agregados densos de células mesenquimáticas denominadas condensaciones precartilaginosas. Están mediadas por la matriz extracelular y moléculas de adherencia celular.[5] El cartílago se diferencia a partir de estas condensaciones, dando lugar a los primordios esqueléticos. En la mayor parte de los esqueletos de miembros quiridios de tetrápodos (con la excepción de algunos anfibios), el cartílago esquelético se reemplaza por hueso en las etapas posteriores del desarrollo.
Periodicidades del patron de la extremidad
La extremidad está organizada en tres regiones, de extremo proximal a distal: estilópodo, zeugópodo y autópodo. En las dos regiones más distales el esqueleto contiene varios motivos o patrones periódicos o cuasiperiódicos. El zeugópodo consta de dos elementos paralelos a lo largo del eje anteroposterior y el autópodo consta de entre 3 y 5 elementos (en la mayoría de los casos) a lo largo del mismo eje. Los dedos también tienen una organización cuasiperiódica a lo largo del eje proximo-distal, compuestos de cadenas en tándem de elementos esqueléticos. La generación del plan básico de la extremidad durante el desarro es producto del mesénquima por la intervención de factores que promueven la condensación precartilaginosa y factores que lo inhiben.[6]
El desarrollo del plan básico de la extremidad está acompañado por la generación de diferencias locales entre estos elementos. Por ejemplo, el radio y la ulna del zeugópodo de las extremidades anteriores son diferentes entre si, como lo son los distintos dedos del autópodo. Estas diferencias se pueden tratar esquemáticamente considerando cómo se reflejan en cada uno de los ejes principales de la extremidad.
Patrones axiales y temas relacionados
Muchos trabajos de investigación sobre el desarrollo de los patrones esqueléticos están influídos por el concepto de información posicional propuesto por Lewis Wolpert en 1971.[7] En sintonía con esta idea se han realizado esfuerzos para identificar moléculas señalizadoras difusibles (morfógenos) que atraviesan los ejes ortogonales de las extremidades en desarrollo y determinan localizaciones e identidades de elementos esqueléticos de un modo dependiente de concentración. No obstante, las células mesenquimatosas del miembro, cuando se disocian y cultivan o se reintroducen en "peladuras" ectodérmicas pueden recapitular aspectos esenciales de formación de patrones, morfogénesis o diferenciación celular.[8] De ese modo, los morfógenos asociados al eje pueden servir realmente a los elementos cartilaginosos de refinamiento secundario formados por un mecanismo "central" relativamente independiente.[6]
Patrones axiales y temas relacionados
Muchos trabajos de investigación sobre el desarrollo de los patrones esqueléticos están influídos por el concepto de información posicional propuesto por Lewis Wolpert en 1971.[7] En sintonía con esta idea se han realizado esfuerzos para identificar moléculas señalizadoras difusibles (morfógenos) que atraviesan los ejes ortogonales de las extremidades en desarrollo y determinan localizaciones e identidades de elementos esqueléticos de un modo dependiente de concentración. No obstante, las células mesenquimatosas del miembro, cuando se disocian y cultivan o se reintroducen en "peladuras" ectodérmicas pueden recapitular aspectos esenciales de formación de patrones, morfogénesis o diferenciación celular.[8] De ese modo, los morfógenos asociados al eje pueden servir realmente a los elementos cartilaginosos de refinamiento secundario formados por un mecanismo "central" relativamente independiente.[6]
Formación del patrón proximal-distal
Los genes Hox contribuyen a la especificación del estilópodo, zeugópodo y autópodo. Las mutaciones de los genes Hox conducen a pérdidas o anormalidades sobre el eje proximal-distal.[9] Existen dos modelos distintos que explican la formación de patrones en este eje:
Modelo de zona de progreso
La AER crea una zona de proliferación celular que se va situando en la extremidad en dirección proximal-distal. La cronología con la que las células abandonan el AER determina su valor posicional. Las estructuras proximales se forman más pronto que las distales.
El modelo de zona de progreso (modelo ZP) fue propuesto hace 30 años pero las últimas evidencias están en conflicto con este modelo:
Evidencia experimental:- Si se extirpa la AER en un periodo tardío del desarrollo se produce menos disrupción de las estructuras distales que si efectúa en en el desarrollo temprano.
- Si se implanta un ápice nuevo del esbozo en el extremo de un esbozo maduro se produce la eliminación y duplicación de estructuras.
Modelo de expansión de progenitores (preespecificación)
Segun este modelo, las células quedan especificadas para cada segmento en el esbozo de extremidad temprano y esta población de células se expande fuera de sus límites a medida que el esbozo crece. Este modelo es consistente con las siguientes observaciones: La división celular se contempla a lo largo de todo el esbozo de la extremidad. La muerte celular tiene lugar en una zona de 200 μm zsubyacente a la AER cuando se eliminaw; la muerte celular ejecuta algún tipo de establecimiento de patrones. Mediante bolitas que liberan FGF se puede rescatar el desarrollo de una extremidad si se extirpa el AER, evitando esta muerte celular.
Evidencia experimental:
- Las celulas marcadas en distintas posiciones de un esbozo de extremidad temprano quedan restringidas a un segmento concreto de este miembro.[10]
- Las extremidades que carecen de la expresion d FGF4 & yFGF8 mostraron todas las estructuras del miembro y no sólo las partes proximales.[11]
No obstante, más recientemente los investigadores que eran partidarios en principio de ambos modelos estan de acuerde en que ninguno de los dos explica adecuadamente los datos experimentales disponibles.[12]
Fijación del patrón antero-posterior
La zona de actividad polarizante (ZPA) del esbozo del miembro tiene una actividad organizadora de patrones gracias a un gradiente morfogénico de Sonic hedgehog (Shh). Shh es suficiente y necesario para crar la ZPA y especificar el patrón antero-posterior en la parte distal del miembro (Shh no es necesario para la polaridad del estilópodo). Shh se activa en la zona posterior mediante la expresión temprana de genes Hoxd genes, la expresión de Hoxb8, y la expresión de dHAND. Shh se mantiene en la zona posterior gracias a un bucle de retroalimentación entre la ZPA la AER. Shh induce que la AER produzca FGF4 y FGF8, lo cual mantiene la expresión de Shh.
Los dedos 3,4 y 5 son especificados por un gradiente temporal de Shh. El dedo 2 está especificado por una forma difusible de amplio rango de Shh y el dedo 1 no requiere Shh. Éste último escinde el complejo represor de la transcripción Ci/Gli3 para convertir el factor de transcripción Gli3 en un activador que ejerce su función sobre los genes HoxD a lo largo del eje antero-posterior. La pérdida del represor Gli3 conduce a la formación de dedos genéricos (sin patron) en cantidades extranumerarias.[13]
Patrón Dorsoventral
La formación del patrón dorsoventral se realiza a partir de señales Wnt7a del ectodermo, no del mesodermo. Wnt7a es necesario y suficiente para dorsalizar la extremidad. Wnt7a también ejerce su infuencia sobre el eje anteroposterior, y la pérdida de Wnt7a provoca que el lado dorsal de los miembros se transforma en ventral, y causando la pérdida de los dedos posteriores. Si se reemplaza la señalización de Wnt7a se rescata este defecto. Wnt7a también se necesita para mantener la expresión de Shh.
Wnt-7a también hace que se exprese Lmx-1, un gen Lim Hox gen (y por lo mismo un factor de transcripción). Lmx-1 está implicado en la dorsalización del miembro, lo que se pone de manifiesto knoqueando el gen Lmx-1 en ratón.[14] Los ratones carentes de Lmx-1 producen piel ventral en ambos lados de sus patas. Hay otros factores que controlan el patrón dorsoventral: Engrailed-1 reprime el efecto dorsalizador de Wnt-7a en el lado ventral de la extremidad.[15]
Patrón Dorsoventral
La formación del patrón dorsoventral se realiza a partir de señales Wnt7a del ectodermo, no del mesodermo. Wnt7a es necesario y suficiente para dorsalizar la extremidad. Wnt7a también ejerce su infuencia sobre el eje anteroposterior, y la pérdida de Wnt7a provoca que el lado dorsal de los miembros se transforma en ventral, y causando la pérdida de los dedos posteriores. Si se reemplaza la señalización de Wnt7a se rescata este defecto. Wnt7a también se necesita para mantener la expresión de Shh.
Wnt-7a también hace que se exprese Lmx-1, un gen Lim Hox gen (y por lo mismo un factor de transcripción). Lmx-1 está implicado en la dorsalización del miembro, lo que se pone de manifiesto knoqueando el gen Lmx-1 en ratón.[14] Los ratones carentes de Lmx-1 producen piel ventral en ambos lados de sus patas. Hay otros factores que controlan el patrón dorsoventral: Engrailed-1 reprime el efecto dorsalizador de Wnt-7a en el lado ventral de la extremidad.[15]
Referencias
- ↑ Stratford T, Horton C, Maden M. (1996). «Retinoic acid is required for the initiation of outgrowth in the chick limb bud.» Curr Biol. Vol. 6. n.º 9. pp. 1124–33. DOI 10.1016/S0960-9822(02)70679-9. PMID 8805369.
- ↑ Yonei-Tamura S, Endo T, Yajima H, Ohuchi H, Ide H, Tamura K. (1999). «FGF7 and FGF10 directly induce the apical ectodermal ridge in chick embryos.» Dev Biol.. Vol. 211. n.º 1. pp. 133–43. DOI 10.1006/dbio.1999.9290. PMID 10373311.
- ↑ Ohuchi H, Takeuchi J, Yoshioka H, Ishimaru Y, Ogura K, Takahashi N, Ogura T, Noji S. (1998). «Correlation of wing-leg identity in ectopic FGF-induced chimeric limbs with the differential expression of chick Tbx5 and Tbx4.» Development.. Vol. 125. n.º 1. pp. 51–60. PMID 9389663.
- ↑ Rodriguez-Esteban C, Tsukui T, Yonei S, Magallon J, Tamura K, Izpisua Belmonte JC. (1999). «The T-box genes Tbx4 and Tbx5 regulate limb outgrowth and identity.» Nature.. Vol. 398. n.º 6730. pp. 814–8. DOI 10.1038/19769. PMID 10235264.
- ↑ Hall BK, Miyake T (2000). «All for one and one for all: condensations and the initiation of skeletal development.» BioEssays. Vol. 22. n.º 2. pp. 138–47. DOI <138::AID-BIES5>3.0.CO;2-4 10.1002/(SICI)1521-1878(200002)22:2<138::AID-BIES5>3.0.CO;2-4. PMID 10655033.
- ↑ a b c Newman SA, Bhat R (2007). «Activator-inhibitor dynamics of vertebrate limb pattern formation.» Birth Defects Res C Embryo Today. Vol. 81. n.º 4. pp. 305–19. DOI 10.1002/bdrc.20112. PMID 18228262.
- ↑ a b Wolpert L (1971). «Positional information and pattern formation.» Curr Top Dev Biol.. Vol. 6. n.º 6. pp. 183–224. DOI 10.1016/S0070-2153(08)60641-9. PMID 4950136.
- ↑ a b Ros MA, Lyons GE, Mackem S, Fallon JF. (2007). «Recombinant limbs as a model to study homeobox gene regulation during limb development.» Dev Biol.. Vol. 166. n.º 1. pp. 59–72. DOI 10.1006/dbio.1994.1296. PMID 7958460.
- ↑ Wellik D, Capecchi M (2003). «Hox10 and Hox11 genes are required to globally pattern the mammalian skeleton.» Science. Vol. 301. n.º 5631. pp. 363–7. DOI 10.1126/science.1085672. PMID 12869760.
- ↑ Dudley A, Ros M, Tabin C (2002). «A re-examination of proximodistal patterning during vertebrate limb development.» Nature. Vol. 418. n.º 6897. pp. 539–44. DOI 10.1038/nature00945. PMID 12152081.
- ↑ Sun X, Mariani F, Martin G (2002). «Functions of FGF signalling from the apical ectodermal ridge in limb development.» Nature. Vol. 418. n.º 6897. pp. 501–8. DOI 10.1038/nature00902. PMID 12152071.
- ↑ Tabin C, Wolpert L (2007). «Rethinking the proximodistal axis of the vertebrate limb in the molecular era.» Genes Dev.. Vol. 21. n.º 12. pp. 1433–42. DOI 10.1101/gad.1547407. PMID 17575045.
- ↑ Chiang C, Litingtung Y, Harris M, Simandl B, Li Y, Beachy P, Fallon J (2001). «Manifestation of the limb prepattern: limb development in the absence of sonic hedgehog function.» Dev Biol. Vol. 236. n.º 2. pp. 421–35. DOI 10.1006/dbio.2001.0346. PMID 11476582.
- ↑ a b Riddle RD, Ensini M, Nelson C, Tsuchida T, Jessell TM, Tabin C (1995). «'Induction of the LIM homeobox gene Lmx1 by WNT7a establishes dorsoventral pattern in the vertebrate limb» Cell. Vol. 17;83(4). n.º 2. pp. 631–40. DOI 10.1016/0092-8674(95)90103-5. PMID 7585966.
- ↑ a b http://genetics.med.harvard.edu/~tabin/Pdfs/Johnson.pdf
Categorías: Biología del desarrollo | Embriología
Wikimedia foundation. 2010.