- Origen de las aves
-
 Un modelo de Archaeopteryx lithographica en exhibición en el Museo de Historia Natural de la Universidad de Oxford.
Un modelo de Archaeopteryx lithographica en exhibición en el Museo de Historia Natural de la Universidad de Oxford.
El origen de las aves ha sido un asunto contencioso dentro de la biología evolutiva por muchos años, pero más recientemente ha surgido un consenso científico que sostiene que las aves son un grupo de dinosaurios terópodos que evolucionaron durante la era Mesozoica. Se propuso una relación cercana entre aves y dinosaurios por primera vez en el siglo XIX luego del descubrimiento del ave primitiva Archaeopteryx en Alemania y ha sido casi confirmado desde la década de 1960 por anatomía comparada y el método cladístico de análisis de las relaciones evolutivas. Los descubrimientos que están ocurriendo de fósiles de dinosaurios con plumas en la provincia de Liaoning en la República Popular China ha permitido una compresión nueva sobre el asunto tanto para científicos como para el público general. En el sentido filogenético las aves son dinosaurios.
Las aves comparten con los dinosaurios varias características del esqueleto que son únicas, especialmente con los terópodos derivados de maniraptora como los dromeosáuridos, los que en la mayoría de los análisis muestran ser sus parientes más próximos. Aunque son más difíciles de identificar en los restos fósiles, existen similitudes en los sistemas digestivo y cardiovascular, así como similitudes de comportamiento y la presencia compartida de plumas, que también conectan a las aves con dinosaurios. El impactante descubrimiento de tejidos blandos fosilizados de Tyrannosaurus rex permitió la comparación de anatomía celular y secuenciación de proteína de tejido de colágeno, los que proveyeron información adicional que corrobora la relación de aves y dinosaurios. [cita requerida]
Sólo unos pocos científicos todavía debaten el origen dinosauriano de las aves, sugiriendo que descienden de otros tipos de reptiles arcosaurianos. Pero entre aquellos que apoyan el origen dinosauriano todavía existen controversias en cuanto a la posición filogenética exacta de las primeras aves dentro de los terópodos. El origen del vuelo de las aves es una interrogante aparte pero relacionada para la que se proponen también varias respuestas.
Contenido
Historia de investigaciones
Huxley, Archaeopteryx y las primeras investigaciones
La investigación científica sobre el origen de las aves comenzó poco después de que en 1859 se publicara El origen de las especies de Charles Darwin, el impactante libro que describió su teoría de la evolución por selección natural.[1] En 1860, una pluma fosilizada fue descubierta en la caliza de Solnhofen del Jurásico tardío en Alemania. Christian Erich Hermann von Meyer describió esta pluma como Archaeopteryx lithographica el año siguiente,[2] y Richard Owen decribió un esqueleto casi completo en 1863, reconociendo este como un ave a pesar de muchas características reminiscentes de reptiles, que incluyen extremidades delanteras con garras y una larga cola ósea.[3]
El biólogo Thomas Henry Huxley, conocido como "el buldog de Darwin" por su apoyo feroz a la nueva teoría de evolución, casi inmediatamente se apoderó de Archaeopteryx como un eslabón fósil entre aves y reptiles. Comenzando en 1868, Huxley hizo comparaciones detalladas de Archaeopteryx con varios reptiles prehistóricos y halló que era más similar a dinosaurios como Hypsilophodon y Compsognathus.[4] [5] El descubrimiento en los finales de la década de 1870 del icónico espécimen de Berlín de Archaeopteryx, completo con un juego de dientes reptilianos, aportó pruebas adicionales. Huxley fue el primero que propuso relación evolutiva entre aves y dinosaurios, aunque tenía la oposición del muy influyente Owen, quien permaneció como un creacionista inamovible. Las conclusiones de Huxley fueron aceptadas por muchos biólogos, incluyendo al Barón Franz Nopcsa,[6] mientras otros, notablemente Harry Govier Seeley,[7] argumentaban que las similitudes eran debidas a convergencia evolutiva.
Heilmann y la hipótesis de tecodontos
Un punto de inflexión surgió en los inicios del siglo XX con los escritos de Gerhard Heilmann de Dinamarca. Aunque era artista de oficio, Heilmann tenía interés académico en las aves y desde 1913 a 1916 publicó los resultados de su investigación en varias partes, tratando con la anatomía, la embriología, el comportamiento, la paleontología y la evolución de las aves.[8] Su trabajo, publicado originalmente en danés como, Vor Nuvaerende Viden om Fuglenes Afstamning, fue compilado, traducido al inglés y publicado en 1926 como The Origin of Birds (El origen de las aves).
Como Huxley, Heilamann comparó a Archaeopteryx y otras aves con una lista exhaustiva de reptiles prehistóricos, y también llegó a la conclusión de que los dinosaurios terópodos como Compsognathus eran los más similares. Sin embargo, Heilmann observó que mientras las aves poseen clavículas fusionadas para formar el hueso llamado fércula (hueso de la suerte), en los reptiles dinosaurios no se habían reconocido aún, aunque si se conocían las clavículas en reptiles más primitivos. Como él era un firme adepto de la Ley de Dollo, la que plantea que la evolución no es reversible, Heilmann no pudo aceptar que las clavículas se hubieran perdido en dinosaurios para re-evolucionar nuevamente en aves. Él por lo tanto estaba forzado a descartar a los dinosaurios como ancestros de las aves y atribuyó todas las similitudes a convergencia evolutiva (la tendencia de los organismos a evolucionar formas similares para estilos de vida similares, como por ejemplo los peces y delfines tienen formas similares). Heilamann planteó que los ancestros de las aves se encontrarían en vez entre los tecodontes, grado de reptiles más primitivos.[9] El trabajo extremadamente exhaustivo de Heilmann aseguró a su libro el volverse un clásico en su campo y sus conclusiones sobre el origen de las aves, como en la mayora de otros asuntos, fue aceptado por casi todos los biólogos evolutivos por las siguientes cuatro décadas.[10]
Las clavículas son huesos relativamente delicados y por lo tanto en peligro de ser destruidos o por lo menos dañados hasta no ser reconocibles. Sin embargo, ya se habían encontrado clavículas de dinosaurios terópodos antes de que se publicara la versión inglesa del libro de Heilmann, pero eso pasó desapercibido entonces.[11]
La ausencia de clavículas en dinosaurios se volvió una visión ortodoxa a pesar del descubrimiento de clavículas en el terópodo primitivo Segisaurus en 1936.[12] El siguiente reporte de clavículas en un dinosaurio fue en 1983, y fue en un artículo ruso publicado antes del final de la Guerra Fría.[13]
Al contrario de lo que creía Heilmann, los paleontólogos que ahora aceptan que las clavículas y en la mayoría de los casos las fúrculas son característica estándar no sólo de los terópodos, sino de los dinosaurios saurischianos. Hasta finales de 2007 fúrculas osificadas (es decir de hueso en vez de cartílago) se habían encontrado en casi todos los tipos de terópodos excepto en los más basales, Eoraptor y Herrerasaurus.[14]
El reporte original de una fúrcula en terópodos primitivos Segisaurus (1936) fue confirmado por re-examinación en 2005.[15] Clavículas unidas como fúrculas también fueron encontradas en Massospondylus, un sauropodomorfo del Jurásico temprano.[16]
Ostrom, Deinonychus y el renacimiento de los dinosaurios
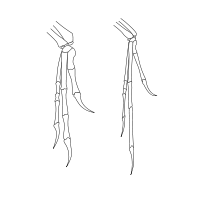 La similitud de las extremidades anteriores de Deinonychus (izquierda) y Archaeopteryx (derecha) llevó a John Ostrom a revivir la conexión entre dinosaurios y aves.
La similitud de las extremidades anteriores de Deinonychus (izquierda) y Archaeopteryx (derecha) llevó a John Ostrom a revivir la conexión entre dinosaurios y aves.La marea comenzó a volverse en contra de la hipótesis 'tecodontos' luego del descubrimiento en 1964 de un nuevo dinosaurio terópodo en Montana. En 1969, este dinosaurio fue descrito y nombrado Deinonychus por John Ostrom de la Universidad de Yale.[17] Al año siguiente, Ostrom redescribió un espécimen de Pterodactylus en el Museo Teyler de Holanda como otro esqueleto de Archaeopteryx.[18] El espécimen consistía principalmente de una sola ala y su descripción le hizo darse cuenta a Ostrom de las similitudes entre las muñecas de Archaeopteryx y Deinonychus.[19]
En 1972, el paleontólogo británico Alick Walker planteó la hipótesis de que las aves surgieron, no de los tecodontes, sino de ancestros de cocodrilo como Sphenosuchus.[20] El trabajo de Ostrom con terópodos y con aves tempranas lo llevó a responder con una serie de publicaciones a mediados de la década de 1970 en las cuales dejó plasmadas las muchas similitudes entre aves y dinosaurios terápodos, resucitando las ideas planteadas por primera vez por Huxley un siglo antes[21] [22] [23] El reconocimiento de Ostrom de la ascendencia dinosauriana de las aves, junto con otras nuevas ideas sobre el metabolismo de los dinosaurios,[24] nivel de actividad y cuidado parental,[25] comenzó lo que se conoce como el renacimiento de los dinosaurios, que desde la década de 1970 continúa aún pasados 30 años.
Las revelaciones de Ostrom coincidieron con la adopción creciente de la sistemática filogenética (cladística), la que comenzó en la década de 1960 con el trabajo de Willi Hennig.[26] La cladística es un método de ordenamiento de especies basado estrictamente en sus relaciones evolutivas, usando un análisis estadístico de sus características anatómicas. En la década de 1980 la metodología cladística fue aplicada a la filogenia de dinosaurios por primera vez por Jacques Gauthier y otros, mostrando inequívocamente que las aves eran un grupo derivado de dinosaurios terápodos.[27] Los primeros análisis sugirieron que los terópodos dromeosauridos como Deinonychus eran de relación particularmente cercana con aves, un resultado que ha sido corroborado desde entonces muchas veces.[28] [29]
Investigación moderna y dinosaurios con plumas en China
 Fósil de Sinosauropteryx prima.
Fósil de Sinosauropteryx prima.En el principio de la década de 1990 se descubrieron fósiles de aves espectacularmente preservados en varias formaciones geológicas del Cretácico Temprano en la provincia Liaoning del nordeste de China.[30] [31] En 1996, paleontólogos chinos describieron Sinosauropteryx como un nuevo género de ave, el que provenía de la formación Yixian,[32] pero este animal fue rápidamente reconocido como un dinosaurio terópodo con relación cercana a Compsognathus. Sorprendentemente, su cuerpo estaba cubierto de largas estructuras filamentosas. Éstas eran 'protoplumas' dudosas y consideradas como homólogas de las más avanzadas plumas de las aves,[33] aunque algunos científicos discordaron de esta aseveración.[34] Científicos chinos y norteamericanos describieron Caudipteryx y Protarchaeopteryx enseguida después. Basados en caracteres esqueléticos, estos animales eran dinosaurios no avianos, pero sus restos portaban plumas completamente formadas semejantes a las de aves.[35] "Archaeoraptor", descrito sin revisión por pares en un número de 1999 de National Geographic,[36] resultó ser una falsificación contrabandeada,[37] Pero restos legítimos continúan saliendo de Yixian, tanto legal como ilegalmente. Plumas o "protoplumas" se han encontrado en una gran variedad de terópodos en Yixian,[38] [39] y los descubrimientos de dinosaurios extremadamente parecidos a aves,[40] así como aves primitivas parecidas a dinosaurios,[41] han cerrado casi completamente el vacío morfológico entre terópodos y aves.
En vez, una minoría pequeña, incluyendo los ornitólogos Alan Feduccia y Larry Martin, continúa planteando que las aves son descendientes de archosaurios tempranos, como Longisquama o Euparkeria.[42] [43] Estudios embriológicos de biología del desarrollo de aves han planteado preguntas sobre la homología de los dedos de extremidades anteriores en aves y dinosaurios.[44] Sin embargo, debido a la conveniente evidencia aportada por la anatomía comparativa y la filogenética, así como los dramáticos fósiles de dinosaurios con plumas de China, la idea de que las aves son derivadas de dinosaurios, primero defendida por Huxley y luego por Nopcsa y Ostrom, tiene apoyo casi unánime entre los paleontólogos actuales.[10]
Filogenia
Archaeopteryx ha sido históricamente considerada como la primera ave. Aunque nuevos descubrimientos fósiles eliminaron el vacío entre terópodos y Archaeopteryx, así como el vacío entre Archaeopteryx y las aves modernas, taxonomistas filogenéticos, manteniendo la tradición, casi siempre usan a Archaeopteryx como un especificador para definir Aves.[45] [46] Aves ha sido definida, en muy raras ocasiones, como un grupo corona consistente sólo en las aves modernas.[27] La gran mayoría de los paleontólogos consideran a las aves como dinosaurios terópodos pertenecientes al clado Coelurosauria.[10] Dentro de Coelurosauria, muchos análisis cladísticos han encontrado apoyo para un clado llamado Maniraptora, el cual consiste, a su vez, en los clados Therizinosauroidea, Oviraptorosauria, Troodontidae, Dromaeosauridae y Aves.[28] [29] [47] De estos, los miembros de los grupos Dromaeosauridae y Troodontidae suelen ser unidos en el clado Deinonychosauria, el cual es el grupo hermano de Aves (que forman juntos el nodo de clados Eumaniraptora) dentro del clado tronco Paraves.[28] [48]
Otros estudios han propuesto filogenias alternativas en las cuales ciertos grupos de dinosaurios que suelen considerarse no avianos, habrían evolucionado de ancestros avianos. Por ejemplo, un análisis de 2002 concluyó que los oviraptosaurios eran avianos basales.[49] Los alvarezsauridos, que se conocen de Asia y América, han sido clasificados anteriormente de diversas maneras como maniraptoranos basales,[28] [29] [50] [51] paravianos,[47] el taxón hermano de Ornithomimosauria,[52] y también como aves tempranas especializadas.[53] [54] El género Rahonavis, descrito originalmente como un ave temprana,[55] ha sido identificado como dromeosáurido, no aviano, en varios estudios.[48] [56] Se ha sugerido que los dromeosáuridos y los troodóntidos mismos pertenecen a Aves en vez de estar fuera del grupo.[57] [58]
Caracteres que enlazan a aves y dinosaurios
Más de cien caracteres anatómicos diferentes son compartidos por aves y dinosaurios terópodos. Algunos de los más interesantes son discutidos aquí:
Plumas
 El Archaeopteryx de Berlín, 1881.
El Archaeopteryx de Berlín, 1881.Archaeopteryx, el primer buen ejemplo de "dinosaurio con plumas", fue descubierto en 1861. El espécimen inicial fue encontrado en la caliza de Solnhofen en el sur de Alemania, que es una lagerstätte, una formación geológica rara y destacable conocida por sus fósiles soberbiamente detallados. Archaeopteryx es también un fósil transicional, con características claramente intermedias entre las de los reptiles modernos y las aves. Sólo dos años después de la innovadora obra de Darwin El origen de las especies, el descubrimiento de Archaeopteryx azuzó el naciente debate entre los proponentes de la biología evolutiva y los del creacionismo. Esta ave temprana es tan similar a dinosaurios que, sin una clara impresión de plumas en la roca circundante, al menos un espécimen fue confundido con Compsognathus.[59]
Desde la década de 1990, se ha encontrado un número adicional de dinosaurios con plumas, que aportan evidencia incluso más fuerte de la relación cercana entre dinosaurios y aves modernas. La mayoría de estos especímenes fueron desenterrados en la provincia Liaoning, del nordeste de China, la que fue parte de una isla continente durante el periodo Cretácico. Aunque las plumas han sido encontradas sólo en la lagerstätte de la formación Yixian y en unos pocos otros lugares, es posible que dinosaurios no avianos en otras partes del mundo también tuvieran plumas. La falta de evidencias fósiles ampliamente distribuidas para dinosaurios no avianos con plumas puede deberse al hecho de que caracteres delicados como piel y plumas no suelen preservarse por la fosilización y por lo tanto están ausentes del registro fósil.
Un desarrollo reciente en el debate se centra en el descubrimiento de impresiones de "protoplumas" rodeando a muchos fósiles de dinosaurios. Las protoplumas sugieren que los tyrannosauroides pueden haber tenido plumas.[60] Sin embargo, otros plantean que estas protoplumas son simplemente el resultado de la descomposición de fibras de colágeno que yace bajo el tegumento de dinosaurios.[43]
 Impresión fósil de NGMC 91, un espécimen probable de Sinornithosaurus.
Impresión fósil de NGMC 91, un espécimen probable de Sinornithosaurus.Todos los dinosaurios con plumas descubiertos, es decir Beipiaosaurus, Caudipteryx, Dilong, Microraptor, Protarchaeopteryx, Shuvuuia, Sinornithosaurus, Sinosauropteryx y Jinfengopteryx, junto a dinosaurios similares a aves, como Confuciusornis, que son cercanos anatómicamente a las aves modernas, han sido encontrados en la misma área y formación en el norte de China. La familia Dromaeosauridae, en particular, parece haber sido densamente emplumada y al menos un dromaeosaurido, Cryptovolans, puede haber sido capaz de volar.
Esqueleto
Debido a que las plumas se asocian a menudo con las aves, los dinosaurios con plumas son a menudo anunciados como el eslabón perdido entre aves y dinosaurios. Sin embargo, las múltiples características esqueléticas también compartidas por ambos grupos representan la conexión más importante para los paleontólogos. Además, es cada vez más claro que la relación entre aves y dinosaurios, y la evolución del vuelo, son asuntos más complejos de lo que antes se pensaba. Por ejemplo, mientras que antes se creía que las aves evolucionaron en una progresión lineal, algunos científicos, más notablemente Gregory S. Paul, concluyen que dinosaurios como los dromeosaurios pueden haber evolucionado a partir de las aves, perdiendo su capacidad de vuelo mientras mantenían sus plumas en una manera similar a lo que ocurre en los modernos avestruz y otros ratites.
Las comparaciones de esqueletos de aves y dinosaurios, así como análisis cladístico, fortalecen el caso del enlace, particularmente para una rama de terópodos llamada maniraptores. Las similitudes en el esqueleto incluyen cuello, pubis, muñeca (carpo semi-lunado), brazo y cintura pectoral, omóplato, fúrcula y quilla del esternón.
Pulmones
Los grandes dinosaurios carnívoros tenían un sistema complejo de sacos aéreos similar a los encontrados en aves modernas, según una investigación dirigida por Patrick O'Connor de la Universidad de Ohio. Los pulmones de dinosaurios terópodos (carnívoros que caminaban en dos patas y tenían pies similares a aves) probablemente bombeaban el aire a las cavidades de los sacos en sus esqueletos, como en el caso de aves. "Lo que formalmente era considerado único de aves estaba presente en alguna forma en los ancestros de aves", dijo O'Connor.[61] [62]
Corazón y postura al dormir
Escaneos modernos de tomografía computarizada (TAC) de la cavidad pectoral de un dinosaurio (realizada en 2000) encontró los aparentes restos de corazón complejo de cuatro cámaras, muy parecido a los que presentan los mamíferos y aves actuales.[63] La idea es controvertida dentro de la comunidad científica, siendo atacado como mala ciencia anatómica[64] o simplemente como pensamiento elucubrativo.[65] Un fósil de troodonto recientemente descubierto demuestra que los dinosaurios dormían como ciertas aves modernas, con sus cabezas recogidas bajo los brazos.[66] Este comportamiento, el cual debe haber ayudado a mantener la cabeza tibia, es también característico de aves modernas.
Biología reproductiva
Un descubrimiento de caracteres de un esqueleto de Tyrannosaurus rex aportó aún más evidencia de caracteres comunes de aves y dinosaurios y permitió, por primera vez, a los paleontólogos establecer el sexo de un dinosaurio. Cuando ponen huevos, las aves hembras desarrollan un tipo especial de tejido dentro de los huesos de sus extremidades. Este hueso medular, rico en calcio, forma una capa que recubre el interior de las cavidades medulares por dentro de la capa ósea dura exterior, y es usada para producir la cáscara de huevos. La presencia de tejidos óseos derivados de endotelio recubriendo el interior de las cavidades medulares de porciones de la extremidad trasera de un espécimen de Tyrannosaurus rex sugiere que esta especie usaba estrategias reproductivas similares a aves, y revela que el espécimen es femenino.[67] Investigaciones adicionales han encontrado hueso medular en el terópodo Allosaurus y en el ornitópodo Tenontosaurus. Debido a que la línea de dinosaurios que incluye a B Allosaurus y Tyrannosaurus divergió de la línea que dio lugar a Tenontosaurus muy temprano en la evolución de los dinosaurios, esto sugiere que los dinosaurios en general producían tejido medular.[68]
Incubación y cuidado de jóvenes
 Un espécimen de Citipati osmolskae que incubaba, se conserva en el American Museum of Natural History en Nueva York.
Un espécimen de Citipati osmolskae que incubaba, se conserva en el American Museum of Natural History en Nueva York.Varios especímenes Citipati se han encontrado apoyados sobre los huevos en su nido en posición aparentemente de incubación.[69]
Numerosas especies de dinosaurios, por ejemplo Maiasaura, se han encontrado en manadas que mezclan individuos muy jóvenes y adultos, sugiriendo interacciones ricas entre ellos.
Un embrión de dinosaurio fue encontrado sin dientes, lo que sugiere que se requería algún cuidado parental para alimentar al joven dinosaurio, posiblemente que el dinosaurio adulto regurgitara el alimento en la boca del joven dinosaurio (ver altricial) Este comportamiento se ve en numerosas especies de aves; los padres regurgitan el alimento en la boca del polluelo.
Molleja
Otra pieza de evidencia de que las aves y los dinosaurios están relacionados estrechamente es el uso de piedras en la molleja. Estas piedras son tragadas por los animales para ayudar a la digestión y el desmenuzado del alimento y las fibras duras una vez que entran al estómago. Cuando se hallan en asociación con fósiles, las piedras de molleja son llamadas gastrolitos.[70] Estas piedras se hallan también en algunos peces (mugiles, alosas, y el gilarú, un tipo de trucha) y en cocodrilos.
Evidencia molecular y tejidos blandos
 Fósil de un individuo juvenil de Scipionyx samniticus. El fósil preserva trazas claras de tejidos blandos.
Fósil de un individuo juvenil de Scipionyx samniticus. El fósil preserva trazas claras de tejidos blandos.Uno de los mejores ejemplos de impresiones de tejido blando en un fósil de dinosaurio fue descubierta en Petraroia, Italia. El descubrimiento fue reportado en 1998, y se describió el espécimen de un coelurosaurio pequeño muy joven, Scipionyx samniticus. El fósil incluye porciones del intestino, colon, hígado, músculos, y tráquea de un dinosaurio inmaduro.[71]
En marzo de 2005, la doctora Mary Higby Schweitzer y su equipo anunciaron el decubrimiento de material flexible con apariencia real de tejido blando dentro de un hueso de 68 millones de años de la pierna de un Tyrannosaurus rex de la formación Hell Creek en Montana. Luego del descubrimiento, el tejido fue rehidratado y se obtuvieron siete tipos de colágeno. Comparando sus datos con los de colágenos de aves vivientes (especialmente de pollo), se reveló que los antiguos terápodos y las aves están estrechamente relacionados.[72]
La extracción exitosa de ADN antiguo de fósiles de dinosaurios ha sido reportada en dos ocasiones separadas, pero en bajo inspección posterior y revisión por pares, ninguno de estos reportes pudo ser confirmado.[73] Sin embargo, un péptido visual funcional de un dinosaurio teórico ha sido inferido usando métodos de reconstrucción filogenética analítica en secuencias de genes de especie modernas relacionadas tales como reptiles y aves.[74] Adicionalmente, varias proteínas han sido supuestamente detectadas fósiles de dinosaurios,[75] incluyendo hemoglobina.[76]
Debates
Origen del vuelo de aves
 El notable Microraptor de cuatro alas, un "primo" de las aves.
El notable Microraptor de cuatro alas, un "primo" de las aves.Debates sobre el origen del vuelo de las aves son casi tan viejos como la idea de que las aves evolucionaron de los dinosaurios, inmediatamente después de 1862. Dos teorías dominaron la mayor parte de la discusión desde entonces: la teoría de corredoras ("desde el suelo para arriba") propone que las aves evolucionaron de predadores pequeños y rápidos que corrían por el suelo; y la teoría de arborícolas ("desde los árboles para abajo") propone que el vuelo propulsado evolucionó de vuelo planeado no propulsado por animales arborícolas (trepadores). Una teoría más reciente, "corredoras en inclinación con asistencia de alas" (WAIR), es una variante de la teoría de corredoras y propone que las alas desarrollaron su funciones aerodinámicas como resultado de la necesidad de correr rápidamente hacia arriba de pendientes muy empinadas, por ejemplo para escapar de depredadores.
Teoría de corredoras ("desde el suelo para arriba")
 Reconstrucción de Rahonavis, un dinosaurio con plumas terrestre que algunos piensan que estaba bien equipado para volar.
Reconstrucción de Rahonavis, un dinosaurio con plumas terrestre que algunos piensan que estaba bien equipado para volar.La teoría de las corredoras para el origen del vuelo fue propuesta por primero por Samuel Wendell Williston, y luego elaborada por Baron Nopcsa. Esta hipótesis propone que algunos animales de carrera rápida con largas colas usaban sus brazos para mantener el equilibrio mientras corrían. Las versiones modernas de esta teoría difieren en muchos detalles desde la versión Williston-Nopcsa, mayormente como resultado de los descubrimientos posteriores a la época de Nopcsa.
Nopsca teorizó que incrementando el área de la superficie de los alargados brazos podía haber ayudado a predadores corredores pequeños a mantener su equilibrio, y que las escamas de las extremidades anteriores se hicieron alargadas, evolucionando en plumas. Nopsca también propuso que existían tres etapas en la evolución del vuelo. Primero, se realizaba vuelo pasivo, en el cual las alas en desarrollo servían a modo de paracaídas. Segundo, fue posible el vuelo activo, en el cual el animal lograba el vuelo por el batir de sus alas. Él usó a Archaeopteryx como ejemplo de esta segunda etapa. Finalmente, las aves ganaron la habilidad para planear.[77]
Ahora se piensa que las plumas no evolucionaron de escamas, dado que las plumas están hechas de proteínas diferentes.[78] Más seriamente, la teoría de Nopsca asume que las plumas evolucionaron como parte de la evolución del vuelo, y descubrimientos recientes prueban que esa suposición es falsa.
Las plumas son muy comunes en dinosaurios coelurosaurianos (incluyendo el tiranosauroideos temprano Dilong).[79] Las aves modernas se clasifican como coelurosaurios por casi todos los palaeontólogos,[80] aunque no por pocos ornitólogos.[81] [82] La versión moderna de la hipótesis "desde el suelo hacia arriba" argumenta que los ancestros de las aves eran pequeños dinosaurios predadores, corredores terrestres, "con plumas" (algo parecidos a correcaminos en su estilo de caza[83] ) que usaban sus extremidades delanteras para el equilibrio mientras perseguían a su presa, y que la patas delanteras y las plumas luego evolucionaron de forma que proveían el planeo y luego propulsaron el vuelo. Las funciones más ampliamente sugeridas de las plumas incluyen el aislamiento térmico y la exhibición competitiva, como en las aves modernas.[84] [85]
Muchos de los fósiles de Archaeopteryx vienen de sedimentos marnos y se ha sugerido que las alas pueden haber ayudado a las ave a correr sobre el agua a la manera del basilisco común o lagarto de Jesucristo (Basiliscus basiliscus).[86]
La mayoría de los ataques recientes a la hipótesis "desde el suelo hacia arriba" intentan refutar la suposición de la versión moderna de que las aves son dinosaurios coelurosaurianos modificados. Los ataques más fuertes se basan en análisis embriológicos que concluyen que las alas de las aves están formadas por los dedos 2, 3 y 4 (correspondientes a los dedos índice, mayor y anular de los humanos; el primero de estos tres en las aves es el álula que se usa para evitar la pérdida de sustentación en vuelo a baja velocidad, por ejemplo en el aterrizaje); pero las "manos" de coelurosaurios están formadas por los dedos 1, 2 y 3 (equivalentes a pulgar índice y mayor en los humanos).[87] Sin embargo estos análisis embriológicos fueron de inmediato desafiados en el terreno embriológico planteándose que la "mano" a menudo se desarrolla de modo diferente en los clados que han perdido algunos dedos en el curso de la evolución, y que las "manos" de las aves en realidad sí se desarrollan de los dedos 1, 2 y 3.[88] [89] [90] Este debate es complejo y aún no resuelto – vea "Homología digital" más abajo.
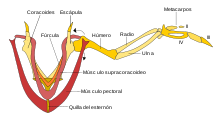 El supracoracoides trabaja usando un sistema como de poleas para levantar el ala, mientras que los pectorales proveen el poderoso batido hacia abajo.
El supracoracoides trabaja usando un sistema como de poleas para levantar el ala, mientras que los pectorales proveen el poderoso batido hacia abajo.Corredoras en inclinación con asistencia de alas
La hipótesis de las "corredoras en inclinación con asistencia de alas" (WAIR) fue incitada por la observación de pollos jóvenes de perdiz chucar (Alectoris chukar), y propone que las alas desarrollaron su función aerodinámica como resultado de la necesidad de correr rápidamente hacia arriba en pendientes muy inclinadas como las de troncos de árboles, por ejemplo para escapar de predadores.[91] Esto la hace un tipo especializado dentro de la teoría de corredoras ("del suelo hacia arriba"). Observe que en este escenario las aves necesitan "empuje hacia abajo" para incrementar el agarre de las patas.[92] [93] Pero las primeras aves, incluyendo a Archaeopteryx, carecían del mecanismo del hombro por el cual las alas de las aves modernas producen rápidos y poderosos batidas hacia arriba, dado que la fuerza hacia abajo de la cual la teoría WAIR depende se genera por batidas hacia arriba, parece que las aves primitivas eran incapaces de realizar la carrera en inclinación con asistencia de alas.[94]
Teoría de arborícolas ("desde los árboles para abajo")
La hipótesis arborícola plantea que los ancestros de aves vivían en los árboles, saltando de rama en rama. Este estilo de vida habría favorecido la evolución de alargados metatarsos (huesos de los pies) y un hallux (dedo correspondiente al gordo) dirigido hacia atrás dando posibilidad de asirse a las ramas. Las extremidades delanteras y traseras se abrían adaptado para propósitos separados, las delanteras para trepar y las traseras para saltar. Esta propone que las delanteras usadas para trepar, se quedaron largas, en vez de reducirse, como es común en la evolución de animales corredores.[77] La superficie de sus "alas" se incrementó progresivamente hasta desarrollar una buena habilidad de planeo. Después del planeo, habrían comenzado a batir las alas para incrementar la eficiencia del vuelo.
Hay poca evidencia de dinosaurios trepadores — sólo Epidendrosaurus/Scansoriopteryx y quizás Microraptor — comparados con la consistencia que se encuentra en los numerosos terópodos terrestres corredores de patas largas, pero estos dinosaurios trepadores han sido encontrados como muy cercanos a las primeras aves en los recientes análisis filogenéticos [cita requerida]. El hecho de que los sedimentos de bosques se preservan muy raramente podría explicar la escasez de sus fósiles.
Algunas investigaciones recientes socavan la hipótesis de "árboles abajo" sugiriendo que las primeras aves y sus ancestros inmediatos no treparon árboles. La aves modernas que se alimentan en árboles tienen garras mucho más curvadas que aquellas que se alimentan en el suelo; las garras de las aves mesozoicas y de los dinosaurios terópodos no avianos estrechamente relacionados son como aquellas de aves modernas que se alimentan en el suelo.[95]
La significación disminuida de Archaeopteryx
Archaeopteryx fue la primera ave y por mucho tiempo fue el único que se conocía de los animales con plumas del Mesozoico (dinosaurios con plumas, si uno acepta el punto de vista de la mayoría de que las aves son dinosaurios modificados). Como resultado la discusión de la evolución de las aves y del vuelo de las aves se centró en Archaeopteryx al menos hasta mediados de la década de 1990.
Ha habido debate acerca de si Archaeopteryx podía realmente volar. Pareciera que Archaeopteryx tenía estructuras craneales y sensores de equilibrio en el oído interno que las aves usan para controlar su vuelo.[96] Archaeopteryx también tenía arreglo de las plumas de vuelo como el de las aves modernas y plumas de vuelo similarmente asimétricas en sus alas y cola. Pero Archaeopteryx carecía del mecanismo del hombro por el cual las alas de aves modernas producen el rápido y fuerte batir hacia arriba (vea el diagrama arriba de la polea supracoracoidea); Esto puede significar que esta y otras aves primitivas eran incapaces de volar batiendo alas y sólo podían planear.[94]
Pero el descubrimiento desde la década de 1990 de muchos dinosaurios con plumas significa que Archaeopteryx ya no es más la figura clave central en la evolución del vuelo de las aves. Otros pequeños coelurosaurios con plumas del Cretácico y Jurásico tardío muestran plumas que pueden ser precursoras del vuelo de aves, por ejemplo: Rahonavis, un corredor terrestre que tenía una garra en forma de hoz elevada en el segundo dedo similar a la de Velociraptor, algunos paleontólogos piensan que estaba mejor adaptado para el vuelo que Archaeopteryx;[97] Epidendrosaurus, un dinosaurio arborícola que puede dar algún apoyo a la teoría "de los árboles hacia abajo";[98] Microraptor, un dinosaurio arborícola que puede haber sido capaz de un vuelo poderoso pero, si así era, más como un biplano dado que tenía plumas bien desarrolladas en las piernas.[99]
 El oviraptosaurio Caudipteryx zoui: ¿un ave no voladora?
El oviraptosaurio Caudipteryx zoui: ¿un ave no voladora?Dinosaurios secundariamente no voladores
Una teoría, defendida por Gregory Paul en sus libros Dinosaurios predadores del mundo (1988) y Dinosaurios de los aires (2002), sugiere que algunos grupos de dinosaurios carnívoros, especialmente deinonychosaurios pero quizás otros como oviraptorosaurios, therizinosaurios, alvarezsauridos y ornitomimosaurios, son realmente descendientes de formas que podían volar. Esta teoría plantea que criaturas parecidas a Archaeopteryx están menos relacionadas a las aves vivientes que estos supuestos dinosaurios secundariamente no voladores.
Aunque en la mayoría de los actuales análisis cladísticos, Archaeopteryx es más cercano a aves que deinonychosaurios o oviraptorosaurios, tales animales como Microraptor o Sinornithosaurus aparentemente quedan cercanos a la base del clado de deinonicosaurianos y parecen tener más adaptaciones para el vuelo que los deinonychosaurios tardíos. Archaeopteryx es todavía suficientemente basal en sus características como para sugerir que los descendientes del Cretácico temprano y medio de las primeras aves podrían teóricamente haber revertido a un modo de vida más dinosauriano. Hesperornis, cuyos ancestros se volvieron no voladores secundariamente alrededor de límite Jurásico/Cretácico, sugiere que el pico aviano fue menos probable de perderse nuevamente que lo que podía perderse la habilidad de volar, pero la posibilidad de que los dientes re-evolucionaran es mayor de lo que a primera vista parece. El hecho de que una gallina moderna naciera con dientes muestra que ese gen particular fue capaz de permanecer silente por casi 100 millones de años y reaparecer en una gallina mutante.[100]
Cladograma simplificado de Mayr et al. (2005)Coelurosauria Therizinosauroideas
Archaeopteryx y Rahonavis
(aves)Deinonychosauria Confuciusornis
(ave)Microraptor
(dromaeosaurio)Dromeosaurios
Grupos que suelen considerarse como aves están en negritas.[58]
Con la inserción de los nuevos datos aportados por el redescrito décimo fósil de Archaeopteryx dentro de una matriz de datos mayor, Mayr y sus colaboradores (2005) mostraron que Archaeopteryx era un clado hermano de un clado constituido por: dos grupos que tradicionalmente son considerados terópodos no avianos, estos son Deinonychosauria y Troodontidae, así como las aves más derivadas, representadas en el análisis por Confuciusornis. Como en la hipótesis de Paul, en este escenario Deinonychosauria y Troodontidae son parte de Aves, el linaje de las aves propiamente dicho, y secundariamente evolucionados como no voladores. Éste es sin embargo un asunto de definición taxonómica y filogenética.[58]Confuciusornis se considera actualmente más cercano a las aves modernas que Archaeopteryx; es improbable que hayan divergido de los ancestros de las aves modernas mucho después de lo que lo hizo Archaeopteryx en cualquier caso,[cita requerida] y su cráneo es muy diferente de ambos. El aparato de vuelo es mucho más avanzado que en Archaeopteryx, pero en una suerte de condición "intermedia" (mientras que el artículo de Mayr et al. quizás indica que no fue intermedio en posición filogenética, dado que resulta un clado hermano de Microraptor). La única cosa que resulta clara del estudio de Mayr y colaboradores es que de las dos aves primitivas comparadas — y ninguna de las dos es necesariamente muy cercana de los ancestros de las aves modernas — Confuciusornis resulta más cercana a un grupo distintivo de terópodos, tradicionalmente vistos como no avianos, de lo que resulta Archaeopteryx.
El artículo despertó un vigoroso debate,[cita requerida] en el cual los autores dejan claro que ellos consideran sus datos aún equívocos en cuanto a qué es lo que ocurrió primero, si el vuelo de las aves o la mayor diversificación de terópodos. ni las aves más modernas que Confuciusornis, ni muchos terópodos interesantes fueron incluidos en el estudio, así que su punto principal del mismo es apoyar el caso de que el vuelo similar al de las aves estaba presente no sólo en los ancestros de las aves modernas. Si es que el vuelo se desarrollo independientemente varias veces como sugiere Barsbold o sólo una vez, con la mayoría, si no todos, los terópodos terrestres que deben ser no voladores secundarios, no se resuelve. Aunque la evaluación estadística de la matriz de datos tentativamente sugiere la última alternativa, la confiabilidad es insuficiente para sacar una conclusión al respecto.
En 2007 se encontraron primordios de plumas en la ulna (uno los dos huesos del "antebrazo") de un Velociraptor, y los investigadores concluyeron que este terópodo tenía plumas de apariencia moderna, compuestas por un raquis (eje) y láminas formadas por barbas. Ellos pensaron que "podría" indicar que Velociraptor tuviera ancestros voladores - pero apuntaron que las plumas podrían haber tenido otras funciones en vez de el vuelo, por ejemplo la exhibición o protección del nido.[101]
Homología de dedos
Existe un debate entre los embriólogos y los paleontólogos sobre si las manos de los dinosaurios terópodos y las aves son esencialmente diferentes, basados en el conteo de las falanges (dedos) en la mano y sus orígenes homólogos. Ésta es un área de investigación importante y fieramente debatida porque sus resultados pueden desafiar el consenso de que las aves son descendientes de los dinosaurios.
Los embriólogos (y algunos paleontólogos que se oponen a la conexión entre aves y dinosaurios) han numerado por mucho tiempo los dedos de las aves como II-III-IV en base a múltiples estudios del desarrollo en el huevo.[102] [103] [104] [105] [106] Esto se basa en el hecho de que en la mayoría de los amniotas, el primer dedo en formarse en una mano de 5 dedos es el cuarto dedo, el que desarrolla un eje primario. Por lo tanto, los embriólogos identifican el eje primario en las aves como el dedo IV, y los dedos sobrevivientes como dedos II-III-IV. Fósiles de manos de terópodos avanzados tienen los dedos I-II-III. Si esto fuera cierto, entonces el desarrollo de los dedos II-III-IV en las aves es incompatible con una ancestralidad en dinosaurios terópodos. Sin embargo, sin ninguna base ontogénica (del desarrollo) para plantear definitivamente cuál dedo es cuál en la mano de los terópodos (porque ningún terópodo puede observarse creciendo y desarrollándose en el presente, excepto las aves), el etiquetado de los dedos de la mano de los terópodos no es conclusivo.
Los paleontólogos han identificado los dedos de las aves como I-II-III. Ellos argumentan que los dedos de las aves se numeran igual que se hace en los de los dinosaurios terópodos, por conservación de la fórmula de las falanges. El conteo de falanges en archeosaurios es 2-3-4-5-3; muchos linajes de archosaurios tienen reducido el número de dedos, pero tienen la misma fórmula de falanges en los dedos que permanecen. En otras palabras, los paleontólogos aseveran que los archosaurios de linajes diferentes tienden a perder los mismos dedos cuando ocurre la pérdida de dedos, desde afuera hacia adentro. Los tres dedos de los dromeosaurios, y Archaeopteryx tienen la misma fórmula de falanges de I-II-III correspondientes a los dedos I-II-II de archosaurios basales. Por lo tanto, los dedos perdidos deberían ser los IV y V. Si esto es real, entonces las aves modernas también poseerían los dedos I-II-III.[106] También, un equipo de investigadores ha propuesto un desplazamiento del marco de conteo de los dedos de los terópodos que dieron lugar a las aves (por lo tanto haciendo al dedo I el II, al II el III, y al III el IV).[107] Sin embargo, tales corrimientos del marco son raros en amniotas y tendrían que ocurrir sólo en las extremidades anteriores sin ocurrir en las posteriores (algo que no se conoce que haya ocurrido en ningún animal) en el linaje de aves-terópodos para poder ser consistente con el origen de las aves en terópodos.[108]
Véase también
- Icnología de aves
- Aves extintas
- Dinosaurios con plumas
- Aves no voladoras
- Origen del vuelo de aves
- Paradoja temporal (paleontología)
Notas
- ↑ Darwin, Charles R. (1859). On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life. London: John Murray. p. 502pp. http://darwin-online.org.uk/content/frameset?itemID=F373&viewtype=side&pageseq=16.
- ↑ von Meyer, C.E. Hermann. (1861). «Archaeopteryx lithographica (Vogel-Feder) und Pterodactylus von Solnhofen» (en alemán). Neues Jahrbuch für Mineralogie, Geologie und Paläontologie 1861: pp. 678–679.
- ↑ Owen, Richard. (1863). «On the Archeopteryx [sp] of von Meyer, with a description of the fossil remains of a long-tailed species, from the lithographic stone of Solenhofen [sp]». Philosophical Transactions of the Royal Society of London 153: pp. 33–47. doi:.
- ↑ Huxley, Thomas H. (1868). «On the animals which are most nearly intermediate between birds and reptiles». Annals of the Magazine of Natural History 4 (2): pp. 66–75.
- ↑ Huxley, Thomas H. (1870). «Further evidence of the affinity between the dinosaurian reptiles and birds». Quarterly Journal of the Geological Society of London 26: pp. 12–31.
- ↑ Nopcsa, Franz. (1907). «Ideas on the origin of flight». Proceedings of the Zoological Society of London: pp. 223–238.
- ↑ Seeley, Harry G. (1901). Dragons of the Air: An Account of Extinct Flying Reptiles. London: Methuen & Co.. p. 239pp.
- ↑ Nieuwland, Ilja J.J. (2004). «Gerhard Heilmann and the artist’s eye in science, 1912-1927». PalArch's Journal of Vertebrate Palaeontology 3 (2). http://www.palarch.nl/Palaeontology/archive.htm. [published online]
- ↑ Heilmann, Gerhard (1926). The Origin of Birds. London: Witherby. p. 208pp.
- ↑ a b c Padian, Kevin. (2004). «Basal Avialae». En Weishampel, David B.; Dodson, Peter; & Osmólska, Halszka (eds.). The Dinosauria (Second Edition edición). Berkeley: University of California Press. pp. 210–231. ISBN 0-520-24209-2.
- ↑ Por ejemplo en 1923, 3 años antes de la compilación inglesa de libro de Heilmann, Roy Chapman Andrews encontró un buen fósil de Oviraptor en Mongolia, pero Henry Fairfield Osborn, quien analizó el fósil en 1924, confundió la fúrcula con una interclavícula; descrita en Paul, G.S. (2002). Dinosaurs of the Air: The Evolution and Loss of Flight in Dinosaurs and Birds. JHU Press. ISBN 0801867630. http://books.google.com/books?id=OUwXzD3iihAC&pg=PA9&lpg=PA9&dq=oviraptor+furcula&source=web&ots=H-AeT6WeoR&sig=K8QkvBqB7RDf5aq0eZh8oKpEMto#PPA9,M1. <!—La primera identificación correcta de clavículas/fúrculas de dinosaurios fue en 1983, también en un Oviraptor: Barsbold, R. (1983). «[Carnivorous dinosaurs from the Cretaceous of Mongolia». Trudy Soumestnaya Sovetsko-Mongol'skaya Paleontogicheskaya Ekspeditsiya 19: pp. 1–117. (¡en ruso!) }} -->
- ↑ Camp, Charles L. (1936). «A new type of small theropod dinosaur from the Navajo Sandstone of Arizona». Bulletin of the University of California Department of Geological Sciences 24: pp. 39–65.
- ↑ In an Oviraptor: Barsbold, R. (1983). «[Carnivorous dinosaurs from the Cretaceous of Mongolia». Trudy Soumestnaya Sovetsko-Mongol'skaya Paleontogicheskaya Ekspeditsiya 19: pp. 1–117. (¡en ruso!) Vea el resumen e imagenes en «A wish for Coelophysis».
- ↑ Lipkin, C., Sereno, P.C., and Horner, J.R. (November 2007). «THE FURCULA IN SUCHOMIMUS TENERENSIS AND TYRANNOSAURUS REX (DINOSAURIA: THEROPODA: TETANURAE)». Journal of Paleontology 81 (6): pp. 1523–1527. doi:. http://jpaleontol.geoscienceworld.org/cgi/content/extract/81/6/1523. – Texto completo actualmente en: «The Furcula in Suchomimus Tenerensis and Tyrannosaurus rex». Lista una gran número de terópodos en los cuales la fúrcula ha sido encontrada, así como se descrinben las de Suchomimus Tenerensis y Tyrannosaurus rex.
- ↑ Carrano, M,R., Hutchinson, J.R., and Sampson, S.D. (December 2005). «New information on Segisaurus halli, a small theropod dinosaur from the Early Jurassic of Arizona». Journal of Vertebrate Paleontology 25 (4): pp. 835–849. doi:. http://www.rvc.ac.uk/AboutUs/Staff/jhutchinson/documents/JH18.pdf.
- ↑ Yates, Adam M.; and Vasconcelos, Cecilio C. (2005). «Furcula-like clavicles in the prosauropod dinosaur Massospondylus». Journal of Vertebrate Paleontology 25 (2): pp. 466–468. doi:.
- ↑ Ostrom, John H. (1969). «Osteology of Deinonychus antirrhopus, an unusual theropod from the Lower Cretaceous of Montana». Bulletin of the Peabody Museum of Natural History 30: pp. 1–165.
- ↑ Ostrom, John H. (1970). «Archaeopteryx: Notice of a "new" specimen». Science 170 (3957): pp. 537–538. doi:. PMID 17799709.
- ↑ Chambers, Paul (2002). Bones of Contention: The Archaeopteryx Scandals. London: John Murray Ltd. pp. 183–184.
- ↑ Walker, Alick D. (1972). «New light on the origin of birds and crocodiles». Nature 237 (5353): pp. 257–263. doi:.
- ↑ Ostrom, John H. (1973). «The ancestry of birds». Nature 242 (5393): pp. 136. doi:.
- ↑ Ostrom, John H. (1975). «The origin of birds». Annual Review of Earth and Planetary Sciences 3: pp. 55–77. doi:.
- ↑ Ostrom, John H. (1976). «Archaeopteryx and the origin of birds». Biological Journal of the Linnean Society 8 (2): pp. 91–182. doi:.
- ↑ Bakker, Robert T. (1972). «Anatomical and ecological evidence of endothermy in dinosaurs». Nature 238 (5359): pp. 81–85. doi:.
- ↑ Horner, John R.; & Makela, Robert (1979). «Nest of juveniles provides evidence of family structure among dinosaurs». Nature 282 (5736): pp. 296–298. doi:.
- ↑ Hennig, E.H. Willi (1966). Phylogenetic Systematics. translated by Davis, D. Dwight; & Zangerl, Rainer.. Urbana: University of Illinois Press.
- ↑ a b Gauthier, Jacques. (1986). «Saurischian monophyly and the origin of birds». En Padian, Kevin. (ed.). The Origin of Birds and the Evolution of Flight. Memoirs of the California Academy of Sciences 8. pp. 1–55.
- ↑ a b c d Senter, Phil (2007). «A new look at the phylogeny of Coelurosauria (Dinosauria: Theropoda)». Journal of Systematic Palaeontology 5 (4): pp. 429–463. doi:.
- ↑ a b c Turner, Alan H.; Hwang, Sunny; & Norell, Mark A. (2007). «A small derived theropod from Öösh, Early Cretaceous, Baykhangor, Mongolia». American Museum Novitates 3557 (3557): pp. 1–27. doi:. http://hdl.handle.net/2246/5845.
- ↑ Sereno, Paul C.; & Rao Chenggang (1992). «Early evolution of avian flight and perching: new evidence from the Lower Cretaceous of China». Science 255 (5046): pp. 845–848. doi:. PMID 17756432.
- ↑ Hou Lian-Hai; Zhou Zhonghe; Martin, Larry D.; & Feduccia, Alan (1995). «A beaked bird from the Jurassic of China». Nature 377 (6550): pp. 616–618. doi:.
- ↑ Ji Qiang; & Ji Shu-an (1996). «On the discovery of the earliest bird fossil in China and the origin of birds». Chinese Geology 233: pp. 30–33.
- ↑ Chen Pei-ji; Dong Zhiming; & Zhen Shuo-nan. (1998). «An exceptionally preserved theropod dinosaur from the Yixian Formation of China». Nature 391 (6663): pp. 147–152. doi:.
- ↑ Lingham-Soliar, Theagarten; Feduccia, Alan; & Wang Xiaolin. (2007). «A new Chinese specimen indicates that ‘protofeathers’ in the Early Cretaceous theropod dinosaur Sinosauropteryx are degraded collagen fibres». Proceedings of the Royal Society B: Biological Sciences 274 (1620): pp. 1823–1829. doi:.
- ↑ Ji Qiang; Currie, Philip J.; Norell, Mark A.; & Ji Shu-an. (1998). «Two feathered dinosaurs from northeastern China». Nature 393 (6687): pp. 753–761. doi:.
- ↑ Sloan, Christopher P. (1999). «Feathers for T. rex?». National Geographic 196 (5): pp. 98–107.
- ↑ Monastersky, Richard (2000). «All mixed up over birds and dinosaurs». Science News 157 (3): pp. 38. doi:. http://www.sciencenews.org/view/generic/id/94/title/All_mixed_up_over_birds_and_dinosaurs.
- ↑ Xu Xing; Tang Zhi-lu; & Wang Xiaolin. (1999). «A therizinosaurid dinosaur with integumentary structures from China». Nature 399 (6734): pp. 350–354. doi:.
- ↑ Xu Xing; Norell, Mark A.; Kuang Xuewen; Wang Xiaolin; Zhao Qi; & Jia Chengkai. (2004). «Basal tyrannosauroids from China and evidence for protofeathers in tyrannosauroids». Nature 431 (7009): pp. 680–684. doi:.
- ↑ Xu Xing; Zhou Zhonghe; Wang Xiaolin; Kuang Xuewen; Zhang Fucheng; & Du Xiangke (2003). «Four-winged dinosaurs from China». Nature 421 (6921): pp. 335–340. doi:.
- ↑ Zhou Zhonghe; & Zhang Fucheng (2002). «A long-tailed, seed-eating bird from the Early Cretaceous of China». Nature 418 (6896): pp. 405–409. doi:.
- ↑ Martin, Larry D. (2006). «A basal archosaurian origin for birds». Acta Zoologica Sinica 50 (6): pp. 977–990.
- ↑ a b Feduccia, Alan; Lingham-Soliar, Theagarten; & Hincliffe, J. Richard. (2005). «Do feathered dinosaurs exist? Testing the hypothesis on neontological and paleontological evidence». Journal of Morphology 266 (2): pp. 125–166. doi:.
- ↑ Burke, Ann C.; & Feduccia, Alan. (1997). «Developmental patterns and the identification of homologies in the avian hand». Science 278 (5338): pp. 666–668. doi:.
- ↑ Sereno, Paul C. (1997). «The origin and evolution of dinosaurs». Annual Review of Earth and Planetary Sciences 25: pp. 435–489. doi:.
- ↑ Chiappe, Luis M. (1997). «Aves». En Currie, Philip J.; & Padian, Kevin. (eds.).. Encyclopedia of Dinosaurs. San Diego: Academic Press. pp. 45–50. ISBN 0-12-226810-5.
- ↑ a b Holtz, Thomas R.; & Osmólska, Halszka. (2004). «Saurischia». En Weishampel, David B.; Dodson, Peter; & Osmólska, Halszka (eds.). The Dinosauria (Second Edition edición). Berkeley: University of California Press. pp. 21–24. ISBN 0-520-24209-2.
- ↑ a b Turner, Alan H.; Pol, Diego; Clarke, Julia A.; Erickson, Gregory M.; & Norell, Mark A. (2007). «A basal dromaeosaurid and size evolution preceding avian flight». Science 317 (5843): pp. 1378–1381. doi:. PMID 17823350.
- ↑ Osmólska, Halszka; Maryańska, Teresa; & Wolsan, Mieczysław. (2002). «Avialan status for Oviraptorosauria». Acta Palaeontologica Polonica 47 (1): pp. 97–116. http://app.pan.pl/article/item/app47-097.html.
- ↑ Martinelli, Agustín G.; & Vera, Ezequiel I. (2007). «Achillesaurus manazzonei, a new alvarezsaurid theropod (Dinosauria) from the Late Cretaceous Bajo de la Carpa Formation, Río Negro Province, Argentina». Zootaxa 1582: pp. 1–17. http://www.mapress.com/zootaxa/2007f/z01582p017f.pdf.
- ↑ Novas, Fernando E.; & Pol, Diego. (2002). «Alvarezsaurid relationships reconsidered». En Chiappe, Luis M.; & Witmer, Lawrence M. (eds.). Mesozoic Birds: Above the Heads of Dinosaurs. Berkeley: University of California Press. pp. 121–125. ISBN 0-520-20094-2.
- ↑ Sereno, Paul C. (1999). «The evolution of dinosaurs». Science 284 (5423): pp. 2137–2147. doi:. PMID 10381873.
- ↑ Perle, Altangerel; Norell, Mark A.; Chiappe, Luis M.; & Clark, James M. (1993). «Flightless bird from the Cretaceous of Mongolia». Science 362 (6421): pp. 623–626. doi:.
- ↑ Chiappe, Luis M.; Norell, Mark A.; & Clark, James M. (2002). «The Cretaceous, short-armed Alvarezsauridae: Mononykus and its kin». En Chiappe, Luis M.; & Witmer, Lawrence M. (eds.). Mesozoic Birds: Above the Heads of Dinosaurs. Berkeley: University of California Press. pp. 87–119. ISBN 0-520-20094-2.
- ↑ Forster, Catherine A.; Sampson, Scott D.; Chiappe, Luis M.; & Krause, David W. (1998). «The theropod ancestry of birds: new evidence from the Late Cretaceous of Madagascar». Science 279 (5358): pp. 1915–1919. doi:. PMID 9506938.
- ↑ Makovicky, Peter J.; Apesteguía, Sebastián; & Agnolín, Federico L. (2005). «The earliest dromaeosaurid theropod from South America». Nature 437 (7061): pp. 1007–1011. doi:.
- ↑ Paul, Gregory S. (2002). Dinosaurs of the Air: The Evolution and Loss of Flight in Dinosaurs and Birds. Baltimore: Johns Hopkins University Press. p. 472pp. ISBN 978-0801867637.
- ↑ a b c Mayr, Gerald; Pohl, Burkhard; & Peters, D. Stefan (2005). «A well-preserved Archaeopteryx specimen with theropod features.». Science 310 (5753): pp. 1483–1486. doi:. PMID 16322455.
- ↑ Wellnhofer, P. (1988). Ein neuer Exemplar von Archaeopteryx. Archaeopteryx 6:1–30.
- ↑ Xu, et al. "Basal tyrannosauroids from China and evidence for protofeathers in tyrannosauroids." Nature. 2004 October 7; 431(7009):680-4. PMID: 15470426
- ↑ O'Connor, P.M. and Claessens, L.P.A.M. (2005). Basic avian pulmonary design and flow-through ventilation in non-avian theropod dinosaurs. Nature 436:253.
- ↑ Paul C. Sereno, Ricardo N. Martinez, Jeffrey A. Wilson, David J. Varricchio, Oscar A. Alcober, Hans C. E. Larsson (2008). «Evidence for Avian Intrathoracic Air Sacs in a New Predatory Dinosaur from Argentina». PLoS ONE 3 (9). doi:. http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0003303.
- ↑ Fisher, P. E., Russell, D. A., Stoskopf, M. K., Barrick, R. E., Hammer, M. & Kuzmitz, A. A. (2000). Cardiovascular evidence for an intermediate or higher metabolic rate in an ornithischian dinosaur. Science 288, 503–505.
- ↑ Hillenius, W. J. & Ruben, J. A. (2004). The evolution of endothermy in terrestrial vertebrates: Who? when? why? Physiological and Biochemical Zoology 77, 1019–1042.
- ↑ Dinosaur with a Heart of Stone. T. Rowe, E. F. McBride, P. C. Sereno, D. A. Russell, P. E. Fisher, R. E. Barrick, and M. K. Stoskopf (2001) Science 291, 783
- ↑ Xu, X. and Norell, M.A. (2004). A new troodontid dinosaur from China with avian-like sleeping posture. Nature 431:838-841.See commentary on the article.
- ↑ Schweitzer, M.H.; Wittmeyer, J.L.; and Horner, J.R. (2005). «Gender-specific reproductive tissue in ratites and Tyrannosaurus rex». Science 308: pp. 1456–1460. doi:. PMID 15933198.
- ↑ Lee, Andrew H.; and Werning, Sarah (2008). «Sexual maturity in growing dinosaurs does not fit reptilian growth models». Proceedings of the National Academy of Sciences 105 (2): pp. 582–587. doi:. PMID 18195356.
- ↑ Norell, M. A., Clark, J. M., Dashzeveg, D., Barsbold, T., Chiappe, L. M., Davidson, A. R., McKenna, M. C. and Novacek, M. J. (November 1994). «A theropod dinosaur embryo and the affinities of the Flaming Cliffs Dinosaur eggs» (abstract page). Science 266 (5186): pp. 779–782. doi:. PMID 17730398. http://www.sciencemag.org/cgi/content/abstract/266/5186/779.
- ↑ Wings O (2007). «A review of gastrolith function with implications for fossil vertebrates and a revised classification». Palaeontologica Polonica 52 (1): pp. 1–16. http://www.app.pan.pl/article/item/app52-001.html.
- ↑ Dal Sasso, C. and Signore, M. (1998). Exceptional soft-tissue preservation in a theropod dinosaur from Italy. Nature 292:383–387. See commentary on the article
- ↑ Schweitzer, M.H., Wittmeyer, J.L. and Horner, J.R. (2005). Soft-Tissue Vessels and Cellular Preservation in Tyrannosaurus rex. Science 307:1952–1955. Also covers the Reproduction Biology paragraph in the Feathered dinosaurs and the bird connection section. See commentary on the article
- ↑ Wang, H., Yan, Z. and Jin, D. (1997). Reanalysis of published DNA sequence amplified from Cretaceous dinosaur egg fossil. Molecular Biology and Evolution. 14:589–591. See commentary on the article.
- ↑ Chang, B.S.W., Jönsson, K., Kazmi, M.A., Donoghue, M.J. and Sakmar, T.P. (2002). Recreating a Functional Ancestral Archosaur Visual Pigment. Molecular Biology and Evolution 19:1483–1489. See commentary on the article.
- ↑ Embery, et al. "Identification of proteinaceous material in the bone of the dinosaur Iguanodon." Connect Tissue Res. 2003; 44 Suppl 1:41-6. PMID: 12952172
- ↑ Schweitzer, et al. "Heme compounds in dinosaur trabecular bone." Proc Natl Acad Sci U S A. 1997 Jun 10; 94(12):6291–6. PMID: 9177210
- ↑ a b Terres, John K. (1980). The Audubon Society Encyclopedia of North American Birds. New York, NY: Knopf. pp. 398–401. ISBN 0394466519.
- ↑ Poling, J. (1996). «Feathers, scutes and the origin of birds». dinosauria.com.
- ↑ Prum, R., and Brush, A.H. (2002). «The evolutionary origin and diversification of feathers» (PDF). The Quarterly Review of Biology 77: pp. 261–295. doi:. http://www.mcorriss.com/Prum_&_Brush_2002.pdf.
- ↑ Mayr, G., B. Pohl & D.S. Peters (2005). "A well-preserved Archaeopteryx specimen with theropod features". Science, 310(5753): 1483-1486.
- ↑ Feduccia, A. (1999). The Origin and Evolution of Birds. Yale University Press. ISBN 9780300078619. http://yalepress.yale.edu/book.asp?isbn=9780300078619. See also Feduccia, A. (February 1995). «Explosive Evolution in Tertiary Birds and Mammals». Science 267 (5198): pp. 637–638. doi:. PMID 17745839. http://taxonomy.zoology.gla.ac.uk/~rdmp1c/teaching/L3/tutorials/feduccia/feduccia.html.
- ↑ Feduccia, A. (1993).
- ↑ Se han encontrado huellas cretácicas de un ave con estilo de vida similar - Lockley, M.G., Li, R., Harris, J.D., Matsukawa, M., and Liu, M. (August, 2007). «Earliest zygodactyl bird feet: evidence from Early Cretaceous roadrunner-like tracks». Naturwissenschaften 94 (8): pp. 657. doi:. http://www.springerlink.com/content/hl850l4128573g33/?p=36f762e67c7a46e493a25f6a7ada455d&pi=0.
- ↑ Burgers, P. and L. M. Chiappe (1999). «The wing of Archaeopteryx as a primary thrust generator». Nature (399): pp. 60–62. http://www.stephenjaygould.org/ctrl/news/file013.html.
- ↑ Cowen, R.. History of Life. Blackwell Science.
- ↑ Videler, J.J. 2005: Avian Flight. Oxford University. Press, Oxford.
- ↑ Burke, A.C., and Feduccia, A. (1997). «Developmental patterns and the identification of homologies in the avian hand» (abstract page). Science 278 (666 fecha=1997): pp. 666. doi:. http://www.sciencemag.org/cgi/content/abstract/278/5338/666?ijkey=dczDGiBvoF7W6. Summarized at «Embryo Studies Show Dinosaurs Could Not Have Given Rise To Modern Birds». ScienceDaily (October 1997).
- ↑ Chatterjee, S. (April 1998). «Counting the Fingers of Birds and Dinosaurs». Science 280 (5362): pp. 355. doi:. http://www.sciencemag.org/cgi/content/full/280/5362/355a.
- ↑ Vargas, A.O., Fallon, J.F. (October 2004). «Birds have dinosaur wings: The molecular evidence» (abstract page). Journal of Experimental Zoology Part B: Molecular and Developmental Evolution 304B (1): pp. 86–90. doi:. http://www3.interscience.wiley.com/cgi-bin/abstract/109741948/ABSTRACT?CRETRY=1&SRETRY=0.
- ↑ Pennisi, E. (January 2005). «Bird Wings Really Are Like Dinosaurs' Hands» (PDF). Science 307: pp. 194–195. doi:. PMID 15653478. http://www.ncsce.org/PDF_files/shift/Pennisi.pdf.
- ↑ There is a video clip of a very young chick doing this at «Wing assisted incline running and evolution of flight».
- ↑ Dial, K.P. (2003). «Wing-Assisted Incline Running and the Evolution of Flight» (abstract page). Science 299 (5605): pp. 402–404. doi:. PMID 12532020. http://www.sciencemag.org/cgi/content/abstract/299/5605/402. Summarized in Morelle, Rebecca (24 January 2008). «Secrets of bird flight revealed» (Web). Scientists believe they could be a step closer to solving the mystery of how the first birds took to the air.. BBC News. Consultado el 25-01-2008.
- ↑ Bundle, M.W and Dial, K.P. (2003). «Mechanics of wing-assisted incline running (WAIR)» (PDF). The Journal of Experimental Biology 206: pp. 4553–4564. doi:. PMID 14610039. http://dbs.umt.edu/flightlab/pdf/bundle%20and%20dial%20JEB%202003.pdf.
- ↑ a b Senter, P. (2006). «Scapular orientation in theropods and basal birds, and the origin of flapping flight». Acta Palaeontologica Polonica 51 (2): pp. 305–313. http://www.app.pan.pl/article/item/app51-305.html.
- ↑ Glen, C.L., and Bennett, M.B. (November 2007) (abstract page). Foraging modes of Mesozoic birds and non-avian theropods. 17. http://www.current-biology.com/content/article/abstract?uid=PIIS0960982207019859.
- ↑ Alonso, P.D., Milner, A.C., Ketcham, R.A., Cokson, M.J and Rowe, T.B. (August 2004). «The avian nature of the brain and inner ear of Archaeopteryx». Nature 430 (7000): pp. 666–669. doi:. http://www.nature.com/nature/journal/v430/n7000/full/nature02706.html.
- ↑ Chiappe, L.M.. Glorified Dinosaurs: The Origin and Early Evolution of Birds. Sydney: UNSW Press.
- ↑ Zhang, F., Zhou, Z., Xu, X. & Wang, X. (2002). «A juvenile coelurosaurian theropod from China indicates arboreal habits». Naturwissenschaften 89 (9): pp. 394–398. doi:.
- ↑ Chatterjee, S., and Templin, R.J. (2007). "Biplane wing planform and flight performance of the feathered dinosaur Microraptor gui." Proceedings of the National Academy of Sciences, 104(5): 1576-1580. [1]
- ↑ Harris, Matthew P.; Sean M. Hasso, Mark W.J. Ferguson and John F. Fallon (February 2006). «The Development of Archosaurian First-Generation Teeth in a Chicken Mutant». Current Biology (Elsevier) 16 (4): pp. 371–377. doi:.
- ↑ Turner, A.H.; Makovicky, P.J.; Norell, M.A. (2007). «Feather quill knobs in the dinosaur Velociraptor». Science 317 (5845): pp. 1721. doi:. PMID 17885130. http://www.sciencemag.org/cgi/content/full/317/5845/1721.
- ↑ Scientists Say No Evidence Exists That Therapod Dinosaurs Evolved Into Birds
- ↑ Scientist Says Ostrich Study Confirms Bird "Hands" Unlike Those Of Dinosaurs
- ↑ Embryo Studies Show Dinosaurs Could Not Have Given Rise To Modern Birds
- ↑ 2 Scientists Say New Data Disprove Dinosaur-Bird Theory - New York Times
- ↑ a b Chatterjee, Sankar (17 April 1998). «Counting the Fingers of Birds and Dinosaurs». Science. doi:10.1126/science.280.5362.355a. Consultado el June 21 de 2007.
- ↑ Wagner, G. P. and Gautthier, J. A. 1999. 1,2,3 = 2,3,4: A solution to the problem of the homology of the digits in the avian hand. Proc. Natl. Acad. Sci. USA 96: 5111-5116
- ↑ Developmental Biology 8e Online. Chapter 16: Did Birds Evolve From the Dinosaurs?
Referencias
- Barsbold, Rinchen (1983): O ptich'ikh chertakh v stroyenii khishchnykh dinozavrov. ["Avian" features in the morphology of predatory dinosaurs]. Transactions of the Joint Soviet Mongolian Paleontological Expedition 24: 96-103. [Original article in Russian.] Translated by W. Robert Welsh, copy provided by Kenneth Carpenter and converted by Matthew Carrano. PDF fulltext
- Bostwick, Kimberly S. (2003): Bird origins and evolution: data accumulates, scientists integrate, and yet the "debate" still rages. Cladistics 19: 369–371. doi 10.1016/S0748-3007(03)00069-0 PDF fulltext
- Dingus, Lowell & Rowe, Timothy (1997): The Mistaken Extinction: Dinosaur Evolution and the Origin of Birds. W. H. Freeman and Company, New York. ISBN 0-7167-2944-X
- Dinosauria On-Line (1995): Archaeopteryx's Relationship With Modern Birds. Retrieved 2006-SEP-30.
- Dinosauria On-Line (1996): Dinosaurian Synapomorphies Found In Archaeopteryx. Retrieved 2006-SEP-30.
- Heilmann, G. (1926): The Origin of Birds. Witherby, London. ISBN 0-486-22784-7 (1972 Dover reprint)
- Mayr, Gerald; Pohl, B. & Peters, D. S. (2005): A Well-Preserved Archaeopteryx Specimen with Theropod Features. Science 310(5753): 1483-1486. doi 10.1126/science.1120331
- Olson, Storrs L. (1985): The fossil record of birds. In: Farner, D.S.; King, J.R. & Parkes, Kenneth C. (eds.): Avian Biology 8: 79-238. Academic Press, New York.











Wikimedia foundation. 2010.