- Historia de la evo-devo
-
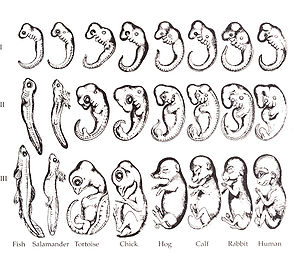 Dibujos de embriones de Ernst Haeckel.
Dibujos de embriones de Ernst Haeckel.
La historia de la biología evolutiva del desarrollo (evo-devo) se remonta a la tradición analógica que, ya en Grecia, estableció un paralelismo entre el desarrollo ontogenético y la organización de los seres vivos. La vinculación entre evolución y desarrollo no tiene lugar hasta la Teoría de la Recapitulación de Ernst Haeckel. Finalmente, la institucionalización de la evo-devo como disciplina autónoma y consolidada tendrá lugar en los años noventa del siglo XX.
Contenido
La tradición analógica
 Aristóteles fue el primero en establecer una analogía entre las formas atravesadas por el embrión durante su ontogenia y las formas animales.
Aristóteles fue el primero en establecer una analogía entre las formas atravesadas por el embrión durante su ontogenia y las formas animales.La idea de recapitulación hunde sus raíces en los presocráticos, para quienes una especie de membrana amniótica primordial habría envuelto al cosmos como a un feto. En Platón encontramos un recapitulacionismo descendente (por degeneración) y en Aristóteles ascendente.[1]
La tradición analógica según la cual existe una relación entre las formas que atraviesa el embrión a lo largo de su desarrollo y las distintas formas animales se remonta a Aristóteles. En la Reproducción de los invertebrados el estagirita concibió la ontogenia como un proceso epigenético que procedía de lo general a lo particular: las primeras etapas se definirían por una forma animal compartida por todo el reino animal que progresivamente se iría concretando para dar lugar, en cada caso, primero a los rasgos genéricos y finalmente a los rasgos propios de cada especie.
La analogía entre ontogenia y Sistema Natural se mantuvo de un modo confuso hasta la obra del preformacionista Charles Bonnet (1720-1793), quien fue el primero en establecer un amplio paralelismo entre los estadios ontogenéticos y la historia de la vida, profundamente ligado a la idea de Scala naturae.
La embriología comparada
El estudio comparativo de la embriología de diversos vertebrados muestra gran similitud con el patrón de desarrollo inicial. Como el embrión se desarrolla, existe individualizantes las características y las similitudes disminuir. Esta similitud se encuentra también en el desarrollo embrionario de todos los animales metazoos. En este caso, sin embargo, cuando varios organismos son diferentes, el período embrionario es menos común entre ellos.
La interpretación filogenética del desarrollo embrionario no habría podido producirse sin el previo desarrollo de la embriología comparada: Martin Heinrich Rathke (1793-1860), N. J. Berrill, D. T. Anderson, Karl Ernst von Baer, Martin Barry.
La obra inaugural del enfoque comparativo de la embriología es, sin duda, la de Kaspar Wolff (1734-1793), quien en su Theoria Generationis (1759) y especialmente en De Formatione Intestinorum (1768) defendió una concepción epigenética del desarrollo y fundó las bases de la futura teoría de las capas germinales.
Christian Pander (1794-1865) fue el primer naturalista en establecer un pararelismo entre el desarrollo ontogenético, la filogenia y el registro paleontológico. Para Pander, tanto el desarrollo individual como la transformación de las especies observada en el registro fósil eran manifestaciones de un mismo fenómeno: la metamorfosis de los seres orgánicos.[2]
La teoría del paralelismo
Inspirados en el marco filosófico de la Naturphilosophie, varios naturalistas pertenecientes a la morfología trascendental alemana establecieron el paralelismo entre el desarrollo ontogenético y el Sistema Natural. Fue el caso de Kielmeyer, Friedrich Tiedemann (1808),[3] Johann Friedrich Meckel (1811) y Carl Gustav Carus (1834):
Es evidente que si procedemos de los estratos más antiguos a los más recientes, puede establacerse una gradación en las formas fósiles desde los animales más imples a los más organizados, de los pólipos a los más complejos, los mamíferos, y que, por lo tanto, el reino animal como un todo tiene sus períodos de desarrollo al igual que el organismo individual. Las especies y los géneros que se han extinguido durante el proceso evolutivo pueden compararse con los órganos que desaparecen durante el desarrollo del animal individual
Tiedemann 1808, p. 73La comparación entre los embriones de los animales superiores y los adultos de los animales inferiores era una idea común a la zoología alemana de la época.
Según Friedrich Kielmeyer (1765-1844), el mundo estaba regido por un sistema de cinco fuerzas: la sensibilidad, la irritabilidad, la reproducción, la secreción y la propulsión. En su obra, Kielmeyer investiga la relación de estas fuerzas en el interior del organismo y su distribución a lo largo de la Scala naturae. A partir de esta concepción fisiológica de la escala de los seres, Kielmeyer postula la idea de la recapitulación.
En Francia, Étienne Serres estableció también el paralelismo entre estadios de desarrollo temprano de ciertos órganos y los órganos en estado adulto de organismos inferiores en el Sistema Natural. En reconocimiento al trabajo de Serres y Meckel, el historiador de la biología británico Russell bautizó a la ley del paralelismo como la ley de Meckel-Serres.[5]
Las leyes de von Baer
En 1837 Martin Barry (1802-1855)[6] defendió que las homologías estructurales se reconocían mejor sin tener en cuenta su función y que las formas de desarrollo eran similares entre diferentes grupos animales.
En sus trabajos sobre los peces fósiles, Louis Agassiz reflexionó largamente en torno a la relación entre la historia de la Tierra y la anatomía comparada, desarrollando la teoría del paralelismo esbozada por Tiedemann, considerando que los estudios embriológicos aportan más indicaciones sobre los animales fósiles que las reconstituciones a partir de fragmentos. No obstante, influenciado por Cuvier, Agassiz limitó el paralelismo a los grandes tipos animales, que consideró irreductibles entre sí. En su recuperación de la teoría del paralelismo, Agassiz refutó las leyes de von Baer acudiendo a numerosos contra-ejemplos donde mostraba las correspondencias entre las formas orgánicas de animales superiores y las formas adultas de especies inferiores actuales.
El arquetipo embriológico
Los embriólogos no tardaron en aplicar su enfoque a las grandes cuestiones planteadas por la morfología: la homología y la unidad de plan. Von Baer fue el primero en reconocer esta posibilidad en 1828. En 1832 Martin Heinrich Rathke utilizó sus observaciones sobre el desarrollo para mostrar la homología entre lo arcos branquiales y el hyoido en los vertebrados. Johannes Peter Müller (1836) utilizó ampliamente los datos embriológicos, considerando que la embriología debía ser el último juez en las polémicas en anatomía comparada. También Carl Boglislaus Reichert (1838) discutió los dos grandes métodos comparativos que permitirían la definición del tipo: el método anatómico (comparación de adultos) y el método embriológico (comparación de embriogenias). En general, la embriología comparativa se propuso el objetivo de construir el "arquetipo embriológico", frente al arquetipo de los morfólogos. La tesis de la supremacía del criterio embriológico frente al morfológico fue muy criticada por Richard Owen y Carl Gustav Carus.
La embriología evolutiva
 Karl Gegenbaur
Karl GegenbaurLa introducción de la dimensión evolucionista en anatomía comparada no supuso una gran ruptura para el trabajo cotidiano y los conceptos de los anatomistas, que continuaron siendo los mismos aunque provistos de una fundamentación realista. Así, las homologías se explicaron por el origen evolutivo común y el arquetipo se convirtió en ancestro común. Lo mismo sucedió con la ley del paralelismo, que pasó a reinterpretarse históricamente como en la teoría de la recapitulación[7] Gerd Müller distingue dos etapas en la embriología evolutiva: una primera etapa descriptiva, centrada en el concepto de homología, y una segunda etapa en la que se trataron de dilucidar los mecanismos responsables de tales semejanzas[8]
Programa descriptivo
Tanto a finales (de Beer, 1971) como en los comienzos (Balfour, 1873) del primer siglo de embriología evolutiva, las investigaciones en torno a la homología fueron centrales para la comprensión de las relaciones entre embriología y evolución.[9]
- Fritz Müller (1821-1897): En Für Darwin (1864)[10] Müller apoyó la teoría de la evolución de Darwin con evidencias procedentes de la biología del desarrollo. Con respecto a la causa evolutiva, Müller defendió la actuación de la selección natural en las larvas y no sólo en los adultos. En 1880, Müller propuso un triple paralelismo entre ontogénesis, filogénesis y regeneración.[11]
- Alexander Kowalevsky (1840-1901) fue uno de los primeros embriólogos en establecer homologías entre diferentes filos. Las técnicas histológicas le permitieron localizarlas en los estadios más tempranos del desarrollo del embrión, como había predicho Karl Ernst von Baer, y su trabajo contribuyó de manera esencial a la aceptación de la evolución.
- Carl Gegenbaur (1826-1903): Su obra más conocida es Grundriss der vergieichenden Anatomk ("Elementos de Anatomía Comparada"), donde subraya la importancia de la embriología para el estudio de la evolución, basándose en la anatomía comparada de los órganos homólogos.
- Francis Balfour: partiendo de la homología embriológica entre las capas germinales, su primera gran contribución, por encargo de Michael Foster, consistió en demostrar la homología entre el nodo de Hesen y el blastoporo.
- William Brooks
- Edwin Stephen Goodrich (1868-1946)
Programa mecanicista y teórico
La teoría de la recapitulación
La idea según la cual el desarrollo repite la evolución estaba implícita en El origen de las especies, aunque su primera exposición detallada se debe a Fritz Müller.[12] Según Müller, el individuo repite durante su desarrollo los cambios mofológicos atravesados por sus ancestros de acuerdo con las leyes de la herencia y la adaptación.
- Ernst Haeckel y su Teoría de la Recapitulación
- Ilya Ilyich Mechnikov fue uno de los primeros en interpretar las leyes de von Baer desde una perspectiva filogenética, considerando que la evolución consiste en la modificación de los embriones y no de los adultos.[13]
- Sir Ray Lankester (1847-1929)
- William Bateson (1861-1926): Bateson estudió con especial atención las transformaciones relacionadas con el número de partes de un organismo y con los reemplazos de órganos. En este contexto, en 1894 acuñó el término homeosis para definir la adquisición por parte de un órgano de características propias de otro órgano. Se trataba, en su opinión, de un concepto más apropiado para entender las homologías, frente a los métodos anatómicos y embriológicos tradicionales. Las diferencias en el número y el tipo de segmentos representaban, para Bateson, patrones evolutivos discontinuos.
- Hans Spemann (1869-1941)
- Walter Garstang (1868–1949): Garstang fue especialmente crítico con la interpretación ortodoxa de la ley biogenética abanderada por Ernst Haeckel. Así, defendió que la evolución podía ocurrir independientemente en larvas y adultos y que la información filogenética se encontraría en las primeras. No obstante, admitió también que muchos de estos rasgos podían deberse a adaptaciones secundarias a la vida larvaria (cenogénesis). Garstang es célebre por su interpretación causal de la ley biogenética. Frente a Haeckel, Garstang defendió que la ontogenia no recapitula la filogenia sino que la crea.
- Gavin de Beer es universalmente reconocido como uno de los grandes precursores de la moderna evo-devo. Su teoría de la heterocronía y su reformulación del concepto de homología se mantienen vigentes.
Evolución y linajes celulares
A finales del siglo XIX varios biólogos trataron de encontrar el vínculo entre evolución y desarrollo en el análisis de los linajes celulares. En 1898 Wilson y Lillie ofrecieron dos conferencias en el Laboratorio de Biología Marina de Woods Hole, Massachusetts, que ilustraron el modo en el que la embriología estaba siendo utilizada en apoyo de la teoría evolutiva.:[13]
- Edmund Beecher Wilson: en 1898 Wilson utilizó las semejanzas entre embriones para describir relaciones filogenéticas. Observando la segmentación espiral en moluscos, platelmintos y anélidos concluyó que los mismos órganos procedían del mismo grupo de células, concluyendo que todos estos organismos debían compartir un ancestro común.
- Frank Rattray Lillie realizó su investigación sobre el desarrollo de los embriones moluscos y la modificación de los linajes celulares. Su obra se concentró en las diferencias de la segmentación. Según Lillie, la biología evolutiva debía estudiar los cambios en el desarrollo embrionario que permitían la supervivencia en determinados ambientes, en lugar de centrarse en las homologías ancestrales que vinculaban a los animales en grandes líneas de ascendencia común.
La separación entre evolución y desarrollo
La embriología experimental
A partir de 1870, un número creciente de embriólogos, insatisfecho con la morfología evolucionista (Huxley, Lankester y Balfour), adopta la búsqueda de las causas directas, materiales, de la forma animal.[14] La embriología experimental se desarrolló en Alemania a finales del siglo XIX con el nombre de Entwicklungsmechanik ("mecánica del desarrollo"). Los principales representantes de esta corriente fueron Wilhelm His, Laurent Chabry, Oscar Hertwig, Wilhelm Roux, Hans Driesch y Theodor Boveri. Más adelante, la escuela americana liderada por Edmund Beecher Wilson y Thomas Hunt Morgan.
La Síntesis moderna
La exclusión de la embriología de la Síntesis moderna.[15]
La obra más representativa es The Theory of the Gene, de Thomas Hunt Morgan, donde establece la distinción entre genética y desarrollo que apartará a la embriología de los estudios filogenéticos: la genética se ocupa de la transmisión de los caracteres y la embriología de la expresión de tales caracteres, por lo que esta última no tiene ninguna relevancia para la biología evolutiva.
Reacciones embriológicas a la Síntesis Moderna
Integradores de la biología del desarrollo en la Teoría Sintética
Alemania: Valentin Haecker
La escuela rusa: Sergei Tchetverikov (1880-1959), Boris L'vovitch Astaurov (1904-1974), Elizaveta I. Balkaschina (1899-1981)
- Boris Ephrussi centró gran parte de su investigación en la herencia citoplasmática.
- Ivan I. Schmalhausen (Factors in Evolution, 1949)
Reino Unido
- Conrad Hal Waddington: canalización (genética), asimilación genética, paisaje epigenético
- Charlotte Auerbach (1899-1994): estudio comparativo entre ejemplares mutantes y salvajes de drosophila con el fin de estudiar las relaciones entre genética y desarrollo.
La reacción de la Síntesis
La Síntesis moderna incorporó muchas de estas aportaciones, pero reinterpretándolas desde una perspectiva poblacional: La selección estabilizadora de Schmalhausen fue aplicada, no a las ontogenias sino a las poblaciones, Dobzhansky redefinió en términos poblacionales el concepto de norma de reacción y, junto con Mayr, interpretó la asimilación genética de Waddington como selección ordinaria con efectos de umbral.[16]
Génesis y consolidación de la evo-devo moderna
Tras el largo eclipse sufrido por la teoría de la recapitulación con la Síntesis evolutiva moderna, en los años 70 y 80 comenzaron a acumularse las dificultades para dar cuenta de ciertas características de la evolución no integradas en la visión neodarwinista: la variación constreñida (Alberch 1982; Maynard Smith et al. 1985), la rapidez de los cambios morfológicos (Eldredge y Gould 1972), los caracteres no adaptativos (Gould and Lewontin 1979) y el origen de organizaciones de alto nivel como la homología o los planes corporales (Riedl 1978). Ante esta situación, en la década de los ochenta se multiplicaron los programas de investigación que trataban de redescubrir los vínculos entre evolución y desarrollo: se organizaron varios encuentros científicos, aparecieron diversos libros sobre la materia y creció la investigación empírica en el campo de la embriología experimental, hasta entonces separada de los problemas evolutivos (Katz et. al 1981, Alberch and Gale 1983, 1985; Raff et al. 1984; Müller 1986, 1989).[8]
Entre los congresos destacan La conferencia Dahlem 1981,[17] editada por Bonner en 1982, la de Sussex en 1982, el simposio sobre desarrollo y evolución celebrado por la British Society of Developmental Biologists y editado por Goodwin, Holder y Wylie en 1983, los congresos en Plzen (1984), Columbia (1985), Woods Hole (1985) y Dijon (1986).
Entre finales de los 70 y la década de los 80 se publicaron varios libros fundamentales: Ontogeny and Phylogeny (Gould,1977);[18] Order in Living Organisms (Riedl, 1978); Embryos, Genes and Evolution : the Developmental Genetic Basis of Evolutionary change (Raff y Kaufman, 1983),[19] Mechanisms of Morphological Evolution de Wallace Arthur en 1984.[20]
La publicación de Ontogenia y Filogenia, de Stephen Jay Gould, fue fundamental. Por primera vez se postulaban mecanismos causales (heterocronía y alometría) que explicarían tanto las homologías como los cambios morfológicos relativamente rápidos. La alometría había sido propuesta por de Beer en 1940 y la alometría por Huxley en 1932, y Gould las utiliza para demostrar cómo los cambios en el desarrollo pueden crear rápidamente las novedades macroevolutivas requeridas por la teoría del equilibrio puntuado.[21]
La institucionalización de la evo-devo
A partir de finales de los noventa la evo-devo se convirtió en una disciplina institucionalizada. Aparecieron dos revistas dedicadas a este campo: Evolution & Development y una sección independiente en el Journal of Experimental Zoology llamada Molecular and Developmental Evolution (ahora Molecular and Developmental Biology). La revista Entwicklungsmechanik fundada por Roux pasó a llamarse Genes, Development, and Evolution[22]
La National Science Foundation estableció un panel especial dedicado a la “evolución de los mecanismos de desarrollo”. El 8 de enero del 2000 la Society for Integrative and Comparative Biology (SICB) inauguró una división destinada a la biología evolutiva del desarrollo, presidida por Rudolf A. Raff. En ella se establecieron como los dos grandes programas de investigación de la evo-devo la explicación de las innovaciones evolutivas y la evolución de los planes corporales.
- John Tyler Bonner: estudio del origen de la multicelularidad en los Myxomycota (1955), edición de las actas de la Conferencia de Dahlem (1982)
- S. J. Gould volvió a poner de actualidad la cuestión de la Recapitulación en su obra Ontogenia y Filogenia (1977), donde presentó un modelo conceptual (el "modelo del reloj") en el que recuperaba modificada la teoría de la heterocronía de Gavin de Beer.
- David B. Wake
- Pere Alberch
- George Oster
- Rupert Riedl
- Viktor Hamburger fue el único embriólogo invitado a la serie de conferencias que darían lugar al clásico libro de Mayr y Provine The Evolutionary Synthesis (1980). Después de Waddington, Hamburger fue el primer biólogo en denunciar la ausencia de la embriología de la Síntesis evolutiva moderna, culpando en parte a los propios embriólogos de esta deficiencia, dadas las reticencias de estos últimos a aceptar la diferencia entre desarrollo y herencia.
- Wallace Arthur
- Stuart Newman
- Gerd Müller
- Günter Wagner
- Brian Goodwin (1983)
- Rudolf A. Raff & Thomas Kaufman: Embryos, Genes, and Evolution (1983)
- Brian K. Hall
- Scott Gilbert
La genética evolutiva del desarrollo
Para algunos autores, el origen de la evo-devo como disciplina consolidada tiene que ver con la transformación que la biología del desarrollo experimentó como consecuencia de los avances en biología molecular. A partir de los años cincuenta la genética del desarrollo conoce un desarrollo espectacular cuyos frutos más impactantes para la teoría evolutiva se resumen en el descubrimiento de los homeobox: el descubrimiento de inesperadas similitudes en la regulación genética de especies filogenéticamente lejanas y del reducido número de reguladores genéticos implicados en la fundación embriológica de todos los planes corporales.[23]
El descubrimiento de las homologías moleculares
Los primeros descubrimientos vinieron del estudio de la homeosis. Como vimos arriba, en los años cuarenta tanto Waddington como Goldschmidt habían identificado mutaciones en la Drosophila por las que un segmento se transformaba en otro tipo de segmento: una antena en una pata o un halterio en un ala. Las llamadas "mutaciones homeóticas" podían ser la clave para comprender las relaciones entre genética, desarrollo y evolución.
En 1978 y 1985 Edward B. Lewis propuso una hipótesis que relacionaba tales mutaciones con la evolución. Partiendo de la teoría de Ohno (1970) de la evolución por duplicación genética, Lewins propuso que el segundo segmento torácico (que posee tanto alas como patas) sería el punto de partida evolutivo para los insectos. El gen responsable de este segmento habría sufrido sucesivas duplicaciones, y conforme estos nuevos genes fuesen activos, se habrían formado una nueva serie de estructuras responsables de la diferenciación de los segmentos. En el último segmento, todos estos genes estarían activos y las mutaciones darían lugar a fenotipos atávicos, como sucede cuando los halterios se convierten en alas.
Desde entonces, las nuevas técnicas moleculares permitieron aislar y secuenciar estos genes. Los resultados obtenidos por los grupos liderados por E. B. Lewis y D. S. Hogness (California), W. Gehring (Basel) y Thomas Kaufman (Indiana) permitieron identificar una región altamente estable que se denominó el homeobox. Los genes responsables de las mutaciones homeóticas se revelaban, así, homólogos. Su localización a lo largo de la práctica totalidad del reino animal demostró su papel en el control de la diferenciación del eje antero-posterior (o de diversos ejes corporales) más que la identidad característica de cada segmento.
A lo largo de los años 80, y gracias a las nuevas técnicas de hibridación molecular, se fue demostrando que los mismos genes homeóticos existían en los vertebrados[24] La siguiente hipótesis consitió en suponer que, puesto que los genes homeóticos eran los responsables del establecimiento del eje antero-posterior en las moscas, los mismos genes podrían crear el eje antero-posterior en los humanos. Esta hipótesis se fue confirmando progresivamente. En primer lugar, se demostró que los genes homeóticos de ciertos vertebrados y de la drosophila estaban agrupados en el mismo orden cromosómico; más tarde, que el patrón de expresión genético en el establecimiento del eje antero-posterior era también el mismo;[25] y, por último, que la región enhancer de un gen homeótico humano (deformed) podía funcionar en la Drosophila activando la expresión de los genes en la misma posición relativa que en el embrión humano.[26] Asimismo, se demostró que la ausencia de estos genes provocaba cambios atávicos, como la formación de mandíbulas y vértebras del cuello reptilianas en ratones.[27]
Todos estos descubrimientos han puesto en cuestión la clásica distinción entre homología y analogía. Es el caso del ojo y el corazón de insectos y vertebrados, cuya formación depende de la expresión de genes homólogos (Pax-6 y Csx/tinman, respectivamente).[28] En general, todos estos resultados moleculares han demostrado la unidad de los mecanismos de desarrollo.
La idea de unidad del mundo animal culminó con la introducción de las nociones de zootipo y estadio filotípico.[29]
- Genetistas del desarrollo: Edward B. Lewis, Christiane Nüsslein-Volhard, Eric Wieschaus, Antonio García-Bellido, Matthew P. Scott, Walter J. Gehring
Referencias
- ↑ Schmitt (2004), p.44
- ↑ Schmitt, S. (2005) "From eggs to fossils: epigenesis and transformation of species in Pander’s biology", Int. J. Dev. Biol. 49: 1-8 [1]
- ↑ En su libro de texto de 1816 declaraba: ""Every animal, before reaching its full development, passes through the stage of organisation of one or more classes lower in the scale, or, every animal begins its metamorphosis with the simplest organisation " (p. 57)
- ↑ Die Zeugung (1805), Lehrbuch der Naturphilosophie (1809-11)
- ↑ Russell, E. S. (1916). Form and Function. A Contribution to the History of Animal Morphology. Londres.
- ↑ Barry, M. 1837. On the unity of structure in the animal kingdom. Edinburgh New Phil. J. 22: 116-141, 345-364.
- ↑ Schmitt, Stéphane (2006). Aux origines de la biologie moderne. L'anatomie comparée d'Aristote à la théorie de l'évolution. Paris: Éditions Belin, p.381..
- ↑ a b Müller, G.B. Evolutionary developmental biology. Handbook of evolution, vol.2.
- ↑ Hall, B.K. (2000) Balfour, Garstang and de Beer: The First Century of Evolutionary Embryology. Amer. Zool., 40: 718–728
- ↑ La traducción al inglés Facts and arguments for Darwin (1869) se encuentra disponible online en el Proyecto Gütenberg
- ↑ Müller, Fritz (1880-1881) "Haeckel's biogenetisches Grundgesetz bei der Neubilding verlorener Glieder", Kosmos, vol. VIII.
- ↑ Russell (1916)
- ↑ a b Gilbert, S. F. 2003. Edmund Beecher Wilson and Frank R. Lillie and the relationship between evolution and development, Developmental Biology, Seventh edition, Sinauer
- ↑ A pesar de la crisis sufrida por la morfología evolucionista, esta corriente se prolonga en el siglo XX en trabajos como los de Goodrich y de Beer
- ↑ Hamburger, 1980; Adams, 1980; Gilbert, 1988
- ↑ Amundson, R. (2005). The Changing Role of the Embryo in Evolutionary Thought: Roots of Evo-Devo. Cambridge: Cambridge University Press. 0521806992., p.196.
- ↑ La conferencia Dahlem ("Dahlem Konferenzen" sobre "Evolución y Desarrollo" tuvo lugar en Berlín entre el 10 y el 15 de mayo de 1981
- ↑ Gould, S.J. (1977). Ontogeny and Phylogeny. Cambridge MA: Harvard Univ. Press.
- ↑ Raff y Kaufman, T.C. (1983) Embryos, Genes, and Evolution: the developmental-genetic basis of evolutionary change. Bloomington: Indiana University Press.
- ↑ Wallace, A. (1984) Mechanisms of Morphological Evolution: a Combined Genetic, Developmental and Ecological Approach. New York: Wiley
- ↑ Gilbert, Scott F., Opitz, John M., Rudolf A. Raff (1996) Resynthesizing Evolutionary and Developmental Biology. Developmental biology 173, 357–372
- ↑ Manfred D. Laubichler and Jane Maienschein (2007) Introduction, Laubichler and Maienschein (ed.) From Embryology to Evo-Devo. A History of Developmental Evolution, MIT Press.
- ↑ Para el relato de los descubrimientos moleculares clave en biología del desarrollo seguimos el artículo de Gilbert et al. (1996). Todas las referencias se han obtenido de dicho artículo.
- ↑ El primer gen controlador del desarrollo (gen Hox) en vertebrados fue descubierto por Carrasco et al. (1984) en las ranas.
- ↑ McGinnis and Krumlauf, 1992; Krumlauf, 1993; Bachiller et al., 1994
- ↑ McGinnis et al., 1990; Malicki et al., 1992
- ↑ Chisaka and Capecchi, 1991; Rijli et al., 1993
- ↑ véase el artículo homología para una explicación detallada de ambos casos
- ↑ J. M. W. Slack, P. W. H. Holland & C. F. Graham (1993) "The zootype and the phylotypic stage". Nature 361, 490 - 492.
- Gilbert (2003) The morphogenesis of evolutionary developmental biology. Int. J. Dev. Biol. 47: 467-477





Wikimedia foundation. 2010.