- Lepospondyli
-
 Lepospondyli
LepospondyliRango temporal: Carbonífero Inferior - Pérmico Inferior 
Rhynchonkos stovalli (A), Phlegethontia longissima (B), Lysorophus tricarinatus (C) & Diplocaulus magnicornis (D)Clasificación científica Reino: Animalia Filo: Chordata Subfilo: Vertebrata Superclase: Tetrapoda Clase: Amphibia (ver el texto) Subclase: Lepospondyli †
Zittel, 1888Clados Ver el texto.
Los lepospóndilos (Lepospondyli) son un grupo extinto de tetrápodos primitivos que vivieron desde comienzos del Carbonífero Inferior (Misisipiense), durante más de 70 Ma, hasta finales del Pérmico Inferior.[1] Con la excepción de un espécimen (Diplocaulus minumus) del grupo Nectridea descubierto en África del Norte (Marruecos), los registros fósiles de lepospóndilos están restringidos a Europa y Norteamérica.[2] Eran animales predominantemente acuáticos y de tamaños, por lo general, reducidos, los cuales exhibían una gran diversidad de formas y adaptaciones, existiendo entre seis y cuatro grupos reconocidos (Acherontiscidae,[3] Adelogyrinidae, Aistopoda, Nectridea, Microsauria y Lysorophia). Lepospondyli fue nombrado en 1888 por Karl Alfred von Zittel, quien acuñó el término para incluir a ciertos tetrápodos paleozoicos que compartían determinadas características en la notocorda y en los dientes.[4]
Contenido
Morfología
Entre las características del grupo destaca la presencia de centros vertebrales (centrum) directamente osificados a partir de las vainas de la notocorda y el que, por lo general, la porción superior de la vértebra, el arco neural, este fusionada con el centro vertebral.[5] Estos centros vertebrales eran alargados y cilíndricos, teniendo usualmente un solo elemento central (al menos en las vértebras presacrales). Al contrario de los temnospóndilos (grupo contemporáneo a Lepospondyli) presentaban tamaños generalmente pequeños (aunque especies del género Diplocaulus podían alcanzar hasta un metro de largo) con cráneos que no superaban los 5 cm y los cuales carecían de colmillos palatales.[6] Carecían, además, de muesca ótica, una pequeña depresión presente en la región posterior de los cráneos de los temnospóndilos y de algunos anfibios modernos (Lissamphibia). Exhibían formas particularmente heterogéneas que iban desde cuerpos elongados con presencia de pedomorfismo (Microbrachis), serpentiformes (Ophiderpeton), compactos y adaptados a una vida terrestre (Pantylus), con cráneos semejantes a un boomerang (Diploceraspis) o con una morfología similar al de las salamandras (Urocordylus).
Diversidad
La especies del grupo Adelogyrinidae, cuyos registros fósiles están restringidos al Carbonífero Inferior (Misisipiense) de Escocia,[7] presentaban una morfología enlongada como adaptación a un medio predominantemente acuático. Poseían extremidades desarrolladas, un cráneo solidificado, elongado y con órbitas localizadas muy adelante. Retenían la cintura escapular pero no otros elementos del esqueleto apendicular.[8] En las especies Adelogyrinus simorhynchus y Palaeomolgophis scoticus existe una larga columna vertebral presacral, pero extremidades bien desarrollas.
De manera similar, las especies del grupo Aistopoda exhibían una morfología altamente especializada, presentando tamaños que variaban entre 5 centímetros y casi un metro de longitud.[4] Poseían una reducción en las extremidades y la cintura pélvica, junto a un cuerpo particularmente elongado y una larga columna vertebral.[6] Rastros de la cintura pectoral pueden identificarse en la especie Lethiscus stocki, la más basal dentro de los aistópodos, lo que sugiere una pérdida secundaria de las extremidades.[9] [10] Se ha especulado respecto a que los miembros de este clado pudieron ocuparon los mismos nichos ecológicos que los de las serpientes, siendo considerados terrestres más que acuáticos.[11] [12] Los registros fósiles datan desde mediados del período Carbonífero Inferior (Viseano) hasta comienzos del período Pérmico, tanto en Europa como en América del Norte.[9]
A diferencia de los miembros de Aistopoda, los grupos pertenecientes a Nectridea (cuyos registros fósiles datan desde finales del período Carbonífero hasta comienzos del período Pérmico en lo que hoy es Norteamérica, Europa y África del Norte) poseían un tronco corto y una cola larga, siendo semejantes a los caudados (salamandras). Las extremidades eran reducidas pero desarrolladas. Los huesos del carpo y del tarso estaban raramente osificados. Las vértebras exhibían un centrum en forma de bobina, con un arco neural y hemal alargados, estando el primero fusionado.[8] Al igual que la mayoría de los lepospóndilos, eran animales pequeños, con tamaños que oscilaban en los 10 y 50 centímetros. Las especies del grupo Keraterpetontidae presentaban largos cuernos tabulares proyectados posteolateralmente, los cuales dan la impresión de un cráneo en forma de boomerang.[13] [9]
Por otra parte, el grupo más heterogéneo y numeroso de lepospóndilos fue Microsauria (con registros que van desde comienzos del período Carbonífero hasta comienzos del Pérmico[14] ), cuyas especies presentaban una muy diversa gama de adaptaciones y características morfológicas como consecuencia de los distintos nichos ecológicos que ocupaban. Se caracterizaban por la presencia de centros vertebrales cilíndricos fusionados con los arcos neurales (aunque algunas especies exhibían intercentros vertebrales presacrales) y la reducción en el número de huesos del cráneo y del complejo occipital-atlas. Como la gran la mayoría de los lepospóndilos, presentaban tamaños reducidos (dándose, además, casos de pedomorfismo).[15]
Por último, las especies del grupo Lysorophia eran acuáticas, presentando cuerpos y cráneos elongados, más de 100 vértebras presacrales, un patrón único en los huesos de la región superior del cráneo y extremidades muy reducidas que son similares en proporción a la de las anfiumas.[4] [16] Ciertas especies estaban adaptadas a una vida en el subsuelo en zonas lodosas dentro de pequeños cuerpos de aguas, recurriendo a la estivación como medio para afrontar sequías periódicas.[17] [18] [19] Por otra parte, el descubrimiento de huellas fósiles asignadas a este grupo sugiere que algunas especies se desplazaban con movimientos ondulatorios al nadar.[20] Los registros fósiles de Lysorophia datan desde finales del período Carbonífero hasta comienzos del período Pérmico.
Evolución y sistemática
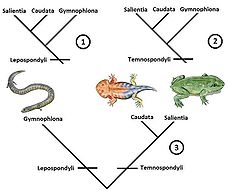 Hipótesis sobre las relaciones filogenéticas de los anfibios modernos:
Hipótesis sobre las relaciones filogenéticas de los anfibios modernos:
1. Origen monofilético a partir de Lepospondyli
2. Origen monofilético a partir de Temnospondyli
3. Origen polifilético.Los especialistas no han llegado a un consenso respecto a si los lepospóndilos son o no anfibios, debido a que no existe una certeza sobre las relaciones filogenéticas de los anfibios modernos (Lissamphibia).[21] Las discusiones actuales están centradas en tres hipótesis, de las cuales una de ellas postula un origen monofilético de Lissamphibia a partir de los lepospóndilos, en cuyo caso se incluiría a Lepospondyli dentro de Amphibia.[22] [23] [24] [25] Por otro lado, se ha propuesto, a su vez, un origen monofilético de Lissamphibia a partir de los temnospóndilos[26] [27] [28] o bien un origen polifilético (en este caso difilético), con los caudados y anuros derivando de los temnospóndilos, mientras que los gimnofiones formarían parte de los lepospóndilos.[29] [30] [31]
Esta última hipótesis se vio reforzada por el descubrimiento de la especie Eocaecilia micropodia, una cecilia del Jurásico Inferior que fue asociada con la especie Rhynchonkos stovalli (Pérmico Inferior) del grupo Microsauria.[8] Sin embargo, esta visión de una relación entre E. micropodia y Microsauria ha sido rechazada por ciertos investigadores,[32] quienes argumentan que las apomorfías atribuidas a esa relación son homoplásticas, reflejando adaptaciones convergentes a un estilo de vida fosorial.[33] Por otra parte, un origen monofilético de los anfibios modernos con respeto a los linajes actuales de amniotas está fuertemente avalado por los análisis moleculares.[34] [35] [36] Por lo tanto, si los lepospóndilos son un grupo más cercano a los amniotas, la hipótesis del origen polifilético puede ser indirectamente rechazada ya que requeriría de una relación más cercana de las cecilias con los amniotas que con el clado Batrachia.
Las relaciones con respecto a los otros tetrápodos del Paleozoico no han sido completamente esclarecidas, aunque los últimos estudios han concluido una relación más cercana con los amniotas (que están dentro de un clado compuesto por el grupo Diadectomorpha y Solenodonsaurus) que con los temnospóndilos.[37] [38] [39]
Por otra parte, las relaciones filogenéticas entre los distintos grupos de lepospóndilos tampoco han sido aclaradas, siendo aún materia de debate debido a que tanto los detalles en la anatomía del cráneo, la forma general del cuerpo, y presumiblemente del estilo de vida, varían considerablemente. A su vez, los miembros más antiguos de cada uno de estos grupos son altamente especializados y muestran, a simple vista, poca evidencia de haber evolucionado de un ancestro en común. A pesar de las muchas autopomorfías presentes, los análisis filogenéticos recientes respaldan un ancestro en común entre estos grupos.[40] [41] [29]
Ruta & Coates (2007) establecen a la especie Westlothiana lizziae como el grupo hermano de Lepospondyli, mientras que tanto Acherontiscus caledoniae como las especies de Adelogyrinidae, conformarían un clado que incluye al grupo Colosteidae.[37] Dentro del mismo estudio se propone un carácter parafilético de Microsauria con respecto a los otros lepospóndilos, a Nectridea como parafilético respecto a Aistopoda y a Lysorophia como el grupo hermano de este último clado.
A diferencia de Ruta & Coates (2007), Anderson (2001) y Vallin & Laurin (2004) no excluyen a Adelogyrinidae de Lepospondyli, pero rescatan también un carácter parafilético de Microsauria. Los resultados de Anderson (2001) coinciden con la parafilia de Microsauria repecto a los otros lepospóndilos y la del grupo Nectridea, el cual incluye, además de Aistopoda, a Lysorophia.[29] Por su parte, Vallin & Laurin (2004) dividen a Lepospondyli en dos clados. Uno conformado por Adelogrynidae y Aistopoda, mientras que el segundo clado presenta a Nectridea como el linaje más basal y a Lysorophia y los anfibios modernos como grupos hermanos e insertos dentro de Microsauria.[42]
Lepospondyli "Lepospondyli" Leposposndyli Cladogramas basados en Anderson (2001) (A),[29] Vallin & Laurin (2004)[42] (B) y Ruta & Coates (2007) (C).[37]
Referencias
- ↑ Anderson, J. S. & Kissel, R. A. (2002) Lepospondyl diversity in the Early Permian and Late Pennsylvanian. Journal of Vertebrate Paleontology 22(3):32A.
- ↑ Carroll, R. L. (1988) Vertebrate Paleontology and Evolution, WH Freeman & Co.
- ↑ Carroll, R. L. (1969) A new family of Carboniferous amphibians. Palaeontology 12:537–548.
- ↑ a b c Carroll, R. L. et al. (1998) Encyclopedia of Paleoherpetology Part 1. München:Pfeil.
- ↑ Colbert, E H. (1969) Evolution of the Vertebrates, John Wiley & Sons Inc (2nd ed.)
- ↑ a b Aldinger, K. (2006) Palaeobiology of Carboniferous/Permian aistopod amphibians. Technische Universität Bergakademie Freiberg
- ↑ Andrews, S. M. & Carroll, R. L. (1991) [Thttp://cat.inist.fr/?aModele=afficheN&cpsidt=4986823 he Order Adelospondyli: Carboniferous lepospondyl amphibians]. Transactions of the Royal Society of Edinburgh, Earth Sciences 82:239–275.
- ↑ a b c Carroll, R. L. (2007) The Palaeozoic Ancestry of Salamanders, Frogs and Caecilians. Zoological Journal of the Linnean Society 150: 1-140.
- ↑ a b c Benton, M. J. (2004) Vertebrate Paleontology, 3rd Ed. Blackwell Science Ltd.
- ↑ Anderson, J. S. et al. (2001) New information on Lethiscus stocki (Tetrapoda: Lepospondyli: Aistopoda) from high-resolution computed tomography and a phylogenetic analysis of Aistopoda. Canadian Journal of Earth Sciencies: Vol. 40, #8, pp. 1071-1083
- ↑ Anderson, J. S. (2002) Revision of the Aistopod Genus Phlegethontia (Tetrapoda: Lepospondyli). Journal of Paleontology.
- ↑ Germain, D. (2008) A new phlegethontiid specimen (Lepospondyli, Aistopoda) from the Late Carboniferous of Montceau-les-Mines (Saône-et-Loire, France). Geodiversitas 30 (4): 669-680.
- ↑ Cruickshank, A. R. I. & Skews, B. W. (1980) The Functional Significance of Nectridean Tabular Horns (Amphibia: Lepospondyli). Proc. R. Soc. Lond. B 209:513-537.
- ↑ Lombard R. E. & Bolt, J. R. (1999) A microsaur from the Mississippian of Illinois and a standard format for morphological characters. Journal of Paleontology 73(5):908-923.
- ↑ Carroll, R. L. & Gaskill, P. (1978) The order Microsauria, Mem. Amer. Phil. Soc., 126:1–211.
- ↑ Wellstead, C. F. (1991) Taxonomic revision of the Lysorophia, Permo-Carboniferous lepospondyl amphibians. Bull. Am. Mus. Nat. Hist. 209:1–90.
- ↑ Olson, E. C. (1971) A skeleton of Lysorophus tricarinatus (Amphibia:Lepospondyli) from the Hennessey Formation (Permian) of Oklahoma. Journal of Paleontology.
- ↑ Hembree, D. I. et al. (2005) Torridorefugium eskridgensis (new ichnogenus and ichnospecies): Amphibian aestivation burrows from the lower Permian Speiser Shale of Kansas. Journal of Paleontology 79(3):583-593.
- ↑ Hembree, D. I. et al. (2004) Amphibian burrows and ephemeral ponds of the Lower Permian Speiser Shale, Kansas: evidence for seasonality in the midcontinent. Palaeogeography, Palaeoclimatology, Palaeoecology 203:127–152.
- ↑ Braddy, S. J. et al. (2003) Amphibian swimming traces from the Lower Permian of southern New Mexico. Palaeontology 46, 671-683.
- ↑ Zardoya, R. & Meyer, A. (2001) On the origin of and phylogenetic relationships among living amphibians. PNAS 98:7380-7383.
- ↑ Laurin, M. (1998a) The importance of global parsimony and historical bias in understanding tetrapod evolution. Part I—systematics, middle ear evolution, and jaw suspension. Annales des Sciences Naturelles, Zoologie, Paris, 13e Série 19: 1-42.
- ↑ Laurin M. (1998b) The importance of global parsimony and historical bias in understanding tetrapod evolution. Part II—vertebral centrum, costal ventilation, and paedomorphosis. Annales des Sciences Naturelles, Zoologie, Paris, 13e Série 19: 99-114.
- ↑ Marjanović, D. & Laurin, M. (2009) The Origin(s) of Modern Amphibians: A Commentary. Evolutionary Biology 36:3, 336-338
- ↑ Pyron, R. A. (2011) Divergence time estimation using fossils as terminal taxa and the origins of lissamphibia. Syst Biol. 60(4):466-81.
- ↑ Bolt, J. R. (1991) Lissamphibian origins. In Origins of the higher groups of tetrapods: controversy and consensus (ed. H.-P. Schultze & L. Trueb), pp. 194–222. Ithaca, NY: Cornell University Press.
- ↑ Trueb, L. & Cloutier, R. in Origins of the Higher Groups of Tetrapods: Controversy and Consensus (eds Schultze, H.-P. & Trueb, L.) 174–193 (Comstock Publishing Associates, Ithaca, 1991).
- ↑ Sigurdsen & Bolt (2009) The lissamphibian humerus and elbow joint, and the origins of modern amphibians. J Morphol.
- ↑ a b c d Anderson, J. S. (2001) The phylogenetic trunk: Maximal inclusion of taxa with missing data in an analysis of the Lepospondyli. Syst. Biol. 50, 170–193.
- ↑ Carroll, R. L. & Currie, P. J. (1975) Microsaurs as possible apodan ancestors. Zool. J. Linn. Soc. 57, 229–247.
- ↑ Anderson, J. S. et al. (2008) A stem batrachian from the Early Permian of Texas and the origin of frogs and salamanders. Nature 453: 515-518.
- ↑ Jenkins, F. A. et al. (2007) Anatomy of Eocaecilia micropodia, A Limbed Caecilian of the Early Jurassic. Bulletin of the Museum of Comparative Zoology 158(6):285-365.
- ↑ Schoch, R. R. & Milner, A. R. (2004) Structure and implications of theories on the origins of lissamphibians. In: Arratia, G., Wilson, M.V. & Cloutier, R. (eds.): Recent advances in the origin and early radiation of vertebrates, pp. 345-377; München (Pfeil).
- ↑ Hedges, S. B. et al. (1990) Tetrapod phylogeny inferred from 18s and 28s ribosomal RNA sequences and a review of the evidence for amniote relationships. Molecular Phylogenetics and Evolution 7:607-633.
- ↑ Zhang, P. et al. (2005) Mitogenomic Perspectives on the Origin and Phylogeny of Living Amphibians. Systematic Biology 54:391-400.
- ↑ Hay, J. M. et al. (1995) Phylogenetic relationships of amphibian families inferred from DNA sequences of mitochondrial 12S and 16S ribosomal RNA genes. Molecular Biology and Evolution 12:928–937.
- ↑ a b c Ruta, M. & Coates, M. I. (2007) Dates, nodes and character conflict: addressing the lissamphibian origin problem. Journal of Systematic Palaeontology 5:69-122.
- ↑ Ruta, M. et al. (2003) Early tetrapod relationships revisited. Biological Reviews of the Cambridge Philosophical Society. 78: 251-345.
- ↑ Marjanović D. & Laurin, M. (2008b) A reevaluation of the evidence supporting an unorthodox hypothesis on the origin of extant amphibians. Contributions to Zoology 77: 149–199.
- ↑ Laurin M. & Reisz, R. R. (1997) A new perspective on tetrapod phylogeny. In: S. Sumida and K. Martin (eds.) Amniote Origins—Completing the Transition to Land: 9-59. London: Academic Press.
- ↑ Carroll, R. L. (1995) Problems of the phylogenetic analysis of Paleozoic choanates. Bulletin du Muséum national d'Histoire naturelle de Paris 4ème série 17: 389-445.
- ↑ a b Vallin, G & M Laurin (2004) Cranial morphology and affinities of Microbrachis, and a reappraisal of the phylogeny and lifestyle of the first amphibians. Journal of Paleontology 24:56-72.
Enlaces externos
 Wikimedia Commons alberga contenido multimedia sobre Lepospondyli. Commons
Wikimedia Commons alberga contenido multimedia sobre Lepospondyli. Commons Wikiespecies tiene un artículo sobre Lepospondyli. Wikispecies
Wikiespecies tiene un artículo sobre Lepospondyli. Wikispecies- Lepospondyli en Mikko's Phylogeny Archive.
- Lepospondyli en Palaeos.com




Wikimedia foundation. 2010.