- Sistema de clasificación APG III
-
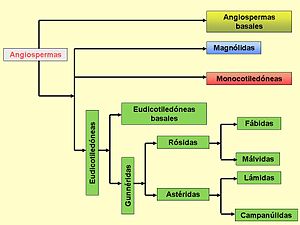 Diagrama simplificado que ilustra el árbol filogenético de las angiospermas. Se muestran los grandes grupos de angiospermas (angiospermas basales, magnólidas, monocotiledóneas y eudicotiledóneas) en diferentes colores. Basado en APG III.[1]
Diagrama simplificado que ilustra el árbol filogenético de las angiospermas. Se muestran los grandes grupos de angiospermas (angiospermas basales, magnólidas, monocotiledóneas y eudicotiledóneas) en diferentes colores. Basado en APG III.[1]
El sistema de clasificación APG III es la última versión del sistema para la clasificación de las angiospermas según criterios filogenéticos. Fue publicado en el 2009 por un vasto grupo de investigadores que se autodenominó «APG III» (del inglés Angiosperm Phylogeny Group, o sea, «grupo para la filogenia de las angiospermas»).[1] Esta versión sucede y reemplaza a aquellas publicadas en 1998 (denominada APG I)[2] y en 2003 (APG II).[1]
Este sistema de clasificación de plantas es diferente de las anteriores aproximaciones al ordenamiento de las angiospermas, que estaban basadas principalmente en criterios morfológicos. El sistema APG III, al igual que las dos versiones anteriores, se basa en datos moleculares —secuencias de ADN del núcleo celular, de la mitocondria y del cloroplasto— y en el análisis filogenético de los mismos. Intenta, de este modo, ordenar la diversidad de las angiospermas sobre la base de su filogenia, es decir, recuperando la evidencia de una serie de eventos únicos que comprende la historia evolutiva de este grupo de plantas. A través de la filogenia se puede comenzar a entender la diversificación, las regularidades en los patrones de la evolución, o simplemente sugerir cambios evolutivos individuales dentro de un clado. De este modo, se descubrieron relaciones entre las angiospermas que obligaban a deshacerse de muchas hipótesis largamente aceptadas acerca de su evolución. Debido a que el árbol filogenético que se desprendió de los análisis de la filogenia mostraba relaciones entre grupos de plantas muy diferentes a lo que se habían hipotetizado previamente (por ejemplo, que la angiosperma basal es Amborella), los botánicos se vieron obligados a rehacer de forma drástica la clasificación de las plantas. El esfuerzo conjunto derivó en las publicaciones firmadas por los tres sucesivos APG.[3]
APG III ordenó y agrupó a las angiospermas en 415 familias, la mayor parte de las cuales se halla incluida en algunos de los 59 órdenes aceptados por este sistema. Tales órdenes, a su vez, se distribuyen en clados.[1]
Concepto
Cada publicación del APG es una caracterización y ordenamiento de todos los órdenes y familias de las angiospermas existentes según su filogenia. Están diseñados para ayudar a la comprensión y a la enseñanza de la filogenia de las angiospermas en un momento dado del conocimiento de los principales clados y de las relaciones dentro y entre cada uno de ellos. Aún no existe una publicación definitiva del APG, ya que todavía se siguen haciendo descubrimientos acerca de la filogenia de varios grupos de angiospermas. De hecho, el árbol filogenético de las angiospermas todavía no está del todo consensuado, ya que hay aún nodos que no tienen un posicionamiento evidente en el árbol. Por esta razón, el sistema de clasificación basado en él todavía se encuentra en pleno desarrollo, y es revisado continuamente, por lo que el APG III no puede ser considerado el sistema de clasificación definitivo de las angiospermas, sino su más reciente aproximación. Una constante puesta al día de la clasificación se encuentra en el Angiosperm Phylogeny Website mantenido por Peter Stevens, uno de los coautores de APG III.[3]
Como particularidad, este sistema sólo abarca categorías taxonómicas desde las especies hasta los órdenes, y no categoriza los taxones por encima de orden. A los grupos de categoría superiores a orden se los denomina con nombres «informales» (no sujetos a las normas del Código Internacional de Nomenclatura Botánica), por lo que es más apropiado llamarlos clados en lugar de taxones. Esta situación puede cambiar en el futuro debido —por un lado— a la mayor certeza que se tiene acerca de las relaciones filogenéticas entre los grandes grupos de angiospermas y —por el otro— al establecimiento del Código Filogénetico de Nomenclatura Biológica («Filocódigo», en inglés «Phylocode»).[4] De hecho, la nomenclatura filogenética, tal como se contempla en el Filocódigo,[4] es una aproximación a la nomenclatura biológica específicamente diseñada para nombrar clados por referencia explícita a su filogenia.[5] [6] [7] [8]
Novedades introducidas
El sistema de clasificación APG III sigue la misma línea que el del APG II y el APG. Las novedades introducidas con respecto a las versiones previas están relacionadas con los recientes descubrimientos en el árbol filogenético, ya que el sistema trata de reflejarlos. Los conocimientos de la filogenia de las angiospermas generados durante la primera década del siglo XXI han determinado varios cambios en la circunscripción de muchas familias y en su clasificación, así como también la adición de algunos órdenes nuevos, tal como se ejemplifica y detalla a continuación.
Nuevos órdenes
En el APG III se adoptaron 14 órdenes nuevos, además de los 45 que ya habían sido aceptados en APG II. Los órdenes nuevos son: Amborellales, Berberidopsidales, Bruniales, Buxales, Chloranthales, Escalloniales, Huerteales, Nymphaeales, Paracryphiales, Petrosaviales, Picramniales, Trochodendrales, Vitales y Zygophyllales.
Filogenia de las angiospermas parásitas
En APG III se ubicó filogenéticamente a la mayoría de las angiospermas parásitas, buena parte de las cuales no se hallaban incluidas en ningún orden en las versiones previas de APG. Debido a la reducción o a la pérdida de muchos caracteres morfológicos taxonómicamente discriminantes, las afinidades de muchos grupos de plantas micoheterótrofas o parásitas han permanecido sin dilucidar durante décadas. Con el advenimiento de la sistemática molecular, se hicieron disponibles nuevas herramientas para identificar a los parientes fotosintéticos de las plantas micoheterótrofas. Así, los datos moleculares han sido utilizados para inferir las relaciones filogenéticas de muchos clados con especies micoheterótrofas.[9] No obstante, los datos moleculares no ofrecen tampoco una respuesta clara y definitiva a las relaciones filogenéticas de este tipo de organismos debido a que, al no ser organismos fotosintéticos, no poseen cloroplastos ni genes de cloroplasto amplificables (los cuales son frecuentemente usados en estudios filogenéticos), o tales genes —cuando existen— son altamente divergentes y las tasas de substitución de los genes nucleares o mitocondriales son muy elevadas, lo que causa un sesgo en las inferencias filogenéticas.[10] Asimismo, la adquisición de muestras representativas de muchos taxones, un factor crítico para realizar una reconstrucción filogenética, es todavía un obstáculo para muchos grupos de micoheterótrofos raros, tales como Corsiaceae, Triuridaceae, Epirixanthes (Polygalaceae), Cheilotheca (Ericaceae) y Thismiaceae.[11]
Nuevas familias y ubicación de géneros o familias de posición taxonómica incierta
Se ubicó en familias a géneros que no habían sido ubicados en familias en APG II, y familias en órdenes, reduciéndose considerablemente el número de taxones que no están incluidos en categorías de orden superior, con respecto a lo que se encontraba en APG II. En este sistema de clasificación quedan solamente 5 taxones (dos pequeñas familias de plantas parásitas y tres géneros) que todavía no se han ubicado en el árbol filogenético de las angiospermas y por lo tanto todavía no fueron ubicados en ningún orden en el APG III.
Gerrardina es un género con dos especies nativas de África. Hacia 1870 el género había sido incluido en las sapindáceas,[12] familia que luego fue incluida en las flacurtiáceas, en la que se retuvo a Gerrardina al momento de describirse la segunda especie del género en 1939.[13] Sobre la base de los aspectos morfológicos, Gerrardina presentaba varios caracteres en común con los restantes miembros de la familia, pero otros muchos para los cuales difería, por lo que se propuso en 2005 su exclusión de la mencionada familia y la creación de una nueva familia para incluirla, Gerrardinaceae. Esta nueva familia, no obstante, no era posible incluirla fácilmente en ningún orden.[14] En 2009, gracias a los análisis filogenéticos realizados sobre datos moleculares de una gran cantidad de taxones, fue posible concluir que Gerrardina en realidad se halla circunscrita en el orden Huerteales[15]
El ejemplo de Gerrardinaceae describe el derrotero de muchas otras familias cuya ubicación taxonómica era desconocida o dudosa y que, gracias a los análisis moleculares realizados en la primera década del año 2000, pudieron asignarse inequívocamente a nuevos órdenes. Estos estudios fueron recogidos por el APG III, de modo que algunas familias fueron reubicadas taxonómicamente, entre las que se encuentran: Hydatellaceae (ahora Nymphaeales), Haptanthaceae (ahora Buxales), Peridiscaceae (ahora Saxifragales), Huaceae (ahora Oxalidales), Centroplacaceae y Rafflesiaceae (ahora ambas del orden Malpighiales), Aphloiaceae, Geissolomataceae y Strasburgeriaceae (ahora todas del orden Crossosomatales), Picramniaceae (Picramniales), Dipentodontaceae y Gerrardinaceae (ahora ambas de Huerteales), Cytinaceae (ahora Malvales), Balanophoraceae (ahora Santalales), Mitrastemonaceae (ahora Ericales) y Boraginaceae (que ahora es considerada al menos un miembro del clado de las lámidas).
Schoepfia es un género de dicotiledóneas herbáceas y parásitas que abarca unas 30 especies nativas de América y Asia tropical. El género estuvo incluido durante muchos años dentro de las olacáceas pero los estudios filogenéticos recientes, basados en datos moleculares, indican que está muy poco relacionado con los restantes miembros de esa familia.[16] Por esa razón se lo ha transferido a una nueva familia, Schoepfiaceae en el orden Santalales.[17] Del mismo modo, muchos otros géneros fueron transferidos o segregados a nuevas familias, tales como Petermanniaceae (en el orden Liliales), Calophyllaceae (en Malpighiales), Capparaceae y Cleomaceae (ambas pertenecientes al orden Brassicales), Anacampserotaceae, Limeaceae, Lophiocarpaceae, Montiaceae y Talinaceae (todas de Caryophyllales) y Linderniaceae y Thomandersiaceae (ambas de Lamiales).[17]
Eliminación de las circunscripciones opcionales para muchas familias
El Grupo para la Clasificación Filogénetica de las Angiospermas (APG) en su trabajo de 1998[18] aceptó a las agapantáceas, aliáceas y amarilidáceas como tres familias independientes dentro del orden de las asparagales. En el trabajo del APG II, del 2003, el reconocimiento de Agapanthaceae y Amaryllidaceae como familias independientes era opcional. Es decir, las tres familias podían considerarse de un modo restringido (Alliaceae, Amaryllidaceae y Agapanthaceae en sentido estricto) o bien, podían también ser incluidas dentro de una circunscripción amplia de las aliáceas (lo que se denomina «Alliaceae sensu lato»).[19] De hecho, los tres grupos son plantas bulbosas con inflorescencias terminales umbeladas, que tienen por debajo brácteas espatáceas y nacen en un notorio escapo, todas estas características probablemente son sinapomórficas. Más aún, los análisis cladísticos basados en datos moleculares sostienen la estrecha relación entre ellas.[20] [21] [22] [23] [24] En la publicación de 2009, el sistema APG III reconfirmó el criterio de considerar a las tres familias como parte de una gran circunscripción de Alliaceae.[25] No obstante, en vez de conservarse el nombre Alliaceae, que tiene preeminencia, se aceptó la propuesta del taxónomo y especialista en plantas bulbosas Alan W. Meerow de utilizar para este grupo el nombre Amaryllidaceae.[26] Muchas otras familias podían ser consideradas en sentido amplio (es decir, una sola familia grande) o estricto (de dos a diez familias más pequeñas definidas de modo más restringido). En APG III, en cambio, se eliminaron esas alternativas debido a su impopularidad (los usuarios de la clasificación preferían que fueran los autores del APG los que decidieran cuál debían utilizar)[27] y, en la mayoría de los casos, se retuvo la circunscripción más amplia para estas familias, como se ha ejemplificado con el caso de las amarilidáceas. Otras familias, además de Amaryllidaceae, en las que se ha modificado la circunscripción son Asparagaceae y Xanthorrhoeaceae (todas Asparagales), Passifloraceae (Malpighiales), Primulaceae (Ericales) y otras familias más pequeñas. De este modo, en APG III se discontinuaron las siguientes familias: Illiciaceae, Alliaceae, Agapanthaceae, Agavaceae, Aphyllanthaceae, Hesperocallidaceae, Hyacinthaceae, Laxmanniaceae, Ruscaceae, Themidaceae, Asphodelaceae, Hemerocallidaceae, Kingdoniaceae, Fumariaceae, Pteridophyllaceae, Didymelaceae, Tetracentraceae, Pterostemonaceae, Hypseocharitaceae, Francoaceae, Memecylaceae, Lepuropetalaceae, Rhoipteleaceae, Medusagynaceae, Quiinaceae, Malesherbiaceae, Turneraceae, Bretschneideraceae, Diegodendraceae, Cochlospermaceae, Peganaceae, Tetradiclidaceae, Nyssaceae, Ternstroemiaceae, Pellicieraceae, Aucubaceae, Donatiaceae, Lobeliaceae, Desfontainiaceae, Diervillaceae, Dipsacaceae, Linnaeaceae, Morinaceae y Valerianaceae.[27]
Con independencia de la eliminación de las circunscripciones opcionales, las siguuientes 18 familias fueron discontinuadas debido a que fueron fusionadas con otras: Limnocharitaceae, Luzuriagaceae, Sparganiaceae, Ledocarpaceae, Heteropyxidaceae, Psiloxylaceae, Oliniaceae, Rhynchocalycaceae, Parnassiaceae, Maesaceae, Myrsinaceae, Theophrastaceae, Eremosynaceae, Polyosmaceae, Tribelaceae, Aralidiaceae, Mackinlayaceae y Melanophyllaceae.[17]
Impacto
Según la prensa del Real Jardín Botánico de Kew, este sistema de clasificación de plantas, que está basado en un conocimiento del árbol filogenético sin precedentes, tendrá un impacto fundamental, no sólo para los científicos, sino también en la forma en que los jardines botánicos organizan sus colecciones, y en el uso futuro de la información acerca de las plantas para mejorar la calidad de vida humana.[27]
El árbol filogenético de las angiospermas
El sistema de clasificación APG III está basado en un árbol filogenético o cladograma de las angiospermas que comprende 59 órdenes y 4 familias.[a] Dicho cladograma se provee a continuación y, debajo del mismo, se brinda el listado de las familias de angiospermas ordenadas en clados y en órdenes.[17]
Angiospermas Amborellales
Mesangiospermas Magnólidas Monocotiledóneas Acorales
Petrosaviales
Commelínidas Arecales
Ceratophyllales
Eudicotiledóneas Trochodendrales
Gunnéridas Pentapétalas Dilleniales
Rósidas Vitales
Eurrósidas Fábidas Málvidas Picramniales
Astéridas Genciánidas Lámidas Campanúlidas*
Campanúlidas Escalloniales
Bruniales
Paracryphiales
Ordenamiento líneal de los clados y de los órdenes de angiospermas

 Flor de Lilium henryi , un miembro del orden de las liliales, dentro del clado de las monocotiledóneas.
Flor de Lilium henryi , un miembro del orden de las liliales, dentro del clado de las monocotiledóneas.
 Flores de Malva sylvestris , un representante del orden de las malvales, dentro del clado de las málvidas.
Flores de Malva sylvestris , un representante del orden de las malvales, dentro del clado de las málvidas.- clado angiospermas
-
- orden Amborellales
- orden Nymphaeales
- orden Austrobaileyales
- orden Chloranthales
- clado magnólidas
-
- orden Canellales
- orden Laurales
- orden Magnoliales
- orden Piperales
-
- clado monocotiledóneas
-
- orden Acorales
- orden Alismatales
- orden Asparagales
- orden Dioscoreales
- orden Liliales
- orden Pandanales
- orden Petrosaviales
- clado commelínidas
-
-
- familia Dasypogonaceae -- Sin circunscribir a ningún orden
- orden Arecales
- orden Commelinales
- orden Poales
- orden Zingiberales
-
-
-
- Probable hermano de la las eudicotiledóneas
-
- orden Ceratophyllales
-
- clado eudicotiledóneas
-
-
- familia Sabiaceae -- Sin circunscribir a ningún orden
- orden Buxales
- orden Proteales
- orden Ranunculales
- orden Trochodendrales
-
- clado gunnéridas o núcleo de las dicotiledóneas verdaderas
-
-
- familia Dilleniaceae -- Sin circunscribir a ningún orden
- orden Gunnerales
- orden Saxifragales
-
- clado rósidas
-
- orden Vitales
- clado fábidas
-
- orden Celastrales
- orden Cucurbitales
- orden Fabales
- orden Fagales
- orden Malpighiales
- orden Oxalidales
- orden Rosales
- orden Zygophyllales
-
- clado málvidas
-
- orden Brassicales
- orden Crossosomatales
- orden Geraniales
- orden Huerteales
- orden Malvales
- orden Myrtales
- orden Picramniales
- orden Sapindales
-
- orden Berberidopsidales
- orden Caryophyllales
- orden Santalales
-
- clado astéridas
- clado lámidas
-
-
-
- familia Boraginaceae -- Sin circunscribir a ningún orden
- familia Vahliaceae -- Sin circunscribir a ningún orden
- familia Icacinaceae -- Sin circunscribir a ningún orden
- familia Metteniusaceae -- Sin circunscribir a ningún orden
- familia Oncothecaceae -- Sin circunscribir a ningún orden
- orden Garryales
- orden Gentianales
- orden Lamiales
- orden Solanales
-
-
-
- clado campanúlidas
-
-
- orden Apiales
- orden Aquifoliales
- orden Asterales
- orden Bruniales
- orden Dipsacales
- orden Escalloniales
- orden Paracryphiales
-
-
-
-
-
Las familias de angiospermas según APG III
APG III ordenó y agrupó a las angiospermas en 415 familias, la mayor parte de las cuales se halla incluida en algunos de los 59 órdenes aceptados por este sistema. Tales órdenes, a su vez, se distribuyen en clados. A continuación se provee el listado de las familias de angiospermas aceptadas por el sistema APG III ordenadas según los clados y los órdenes a los que pertenecen.[1] Además, se provee el nombre de los clados por encima de la categoría taxonómica de orden según la nomenclatura filogenética.[28] En cada caso se han destacado las diferencias con el sistema APG II según los siguientes símbolos: * = nueva ubicación de la familia;
† = nuevo orden reconocido por el sistema APG;
§ = familia con una nueva circunscripción;
º = familias que representan la circunscripción más amplia prevista en el sistema APG II, la cual se adopta en APGIII;
ºº = familias que representan la circunscripción más restringida del sistema APGII, la cual se adopta en APGIII.Angiospermae Lindley [P.D. Cantino & M.J. Donoghue][29]
- †Amborellales Melikyan, A.V.Bobrov & Zaytzeva
- Amborellaceae Pichon
- †Nymphaeales Salisb. ex Bercht. & J.Presl
- Cabombaceae Rich. ex A.Rich.
- *Hydatellaceae U.Hamann
- ººNymphaeaceae Salisb.
- Austrobaileyales Takht. ex Reveal
- Austrobaileyaceae Croizat
- ºSchisandraceae Blume (incluye Illiciaceae A.C.Sm.)
- Trimeniaceae L.S.Gibbs
- †Chloranthales R.Br.
- Chloranthaceae R.Br. ex Sims
Magnoliidae Novák ex Takhtajan [W.S. Judd, P.S. Soltis & D.E. Soltis][30]
El clado de las magnólidas es un grupo de angiospermas que habían sido nombradas informalmente como «magnoliids» o «eumagnoliids» en muchas publicaciones filogenéticas recientes[31] [32] [33] [34] [1] y que difiere en su circunscripción con respecto a la subclase Magnollidae de los sistemas de clasificación de Cronquist y de Takhtajan. Las posibles sinapomorfias de este grupo incluyen caracteres fitoquímicos, como la presencia de los compuestos asarona, galbacina y licarina.[30] Las magnólidas comprenden cuatro órdenes:[17]
- Canellales Cronquist
- Canellaceae Mart.
- Winteraceae R.Br. ex Lindl.
- Piperales Bercht. & J.Presl
- Aristolochiaceae Juss.
- Hydnoraceae C.Agardh
- Lactoridaceae Engl.
- Piperaceae Giseke
- Saururaceae F.Voigt
- Laurales Juss. ex Bercht. & J.Presl
- Atherospermataceae R.Br.
- Calycanthaceae Lindl.
- Gomortegaceae Reiche
- Hernandiaceae Blume
- Lauraceae Juss.
- Monimiaceae Juss.
- Siparunaceae Schodde
- Magnoliales Juss. ex Bercht. & J.Presl
- Annonaceae Juss.
- Degeneriaceae I.W.Bailey & A.C.Sm.
- Eupomatiaceae Orb.
- Himantandraceae Diels
- Magnoliaceae Juss.
- Myristicaceae R.Br.
Monocotyledoneae de Candolle [W.S. Judd, P.S. Soltis, D.E. Soltis & S.W. Graham][35]
- Acorales Link
- Acoraceae Martinov
- Alismatales R.Br. ex Bercht. & J.Presl
- §Alismataceae Vent. (incluye Limnocharitaceae Takht. ex Cronquist)
- Aponogetonaceae Planch.
- Araceae Juss.
- Butomaceae Mirb.
- Cymodoceaceae Vines
- Hydrocharitaceae Juss.
- Juncaginaceae Rich.
- Posidoniaceae Vines
- Potamogetonaceae Bercht. & J.Presl
- Ruppiaceae Horan.
- Scheuchzeriaceae F.Rudolphi
- Tofieldiaceae Takht.
- Zosteraceae Dumort.
- †Petrosaviales Takht.
- Petrosaviaceae Hutch.
- Dioscoreales R.Br.
- Burmanniaceae Blume
- Dioscoreaceae R.Br.
- Nartheciaceae Fr. ex Bjurzon
- Pandanales R.Br. ex Bercht. & J.Presl
- Cyclanthaceae Poit. ex A.Rich.
- Pandanaceae R.Br.
- Stemonaceae Caruel
- Triuridaceae Gardner
- Velloziaceae J.Agardh
- Liliales Perleb
- §Alstroemeriaceae Dumort. (incluye Luzuriagaceae Lotsy)
- Campynemataceae Dumort.
- Colchicaceae DC.
- Corsiaceae Becc.
- Liliaceae Juss.
- Melanthiaceae Batsch ex Borkh.
- *Petermanniaceae Hutch.
- Philesiaceae Dumort.
- Ripogonaceae Conran & Clifford
- Smilacaceae Vent.
- Asparagales Link
- ºAmaryllidaceae J.St.-Hil. (incluye Agapanthaceae F.Voigt, Alliaceae Borkh.)
- ºAsparagaceae Juss. (incluye Agavaceae Dumort., Aphyllanthaceae Burnett, Hesperocallidaceae Traub, Hyacinthaceae Batsch ex Borkh., Laxmanniaceae Bubani, Ruscaceae M.Roem., Themidaceae Salisb.)
- Asteliaceae Dumort.
- Blandfordiaceae R.Dahlgren & Clifford
- Boryaceae M.W.Chase, Rudall & Conran
- Doryanthaceae R.Dahlgren & Clifford
- Hypoxidaceae R.Br.
- Iridaceae Juss.
- Ixioliriaceae Nakai
- Lanariaceae R.Dahlgren & A.E.van Wyk
- Orchidaceae Juss.
- Tecophilaeaceae Leyb.
- ºXanthorrhoeaceae Dumort. (incluye Asphodelaceae Juss. and Hemerocallidaceae R.Br.)
- Xeronemataceae M.W.Chase, Rudall & M.F.Fay
Commelinidae Takhtajan [S.W. Graham & W.S. Judd][36]
Las commelínidas son un gran clado dentro de las monocotiledóneas cuyas sinapomorfías incluyen las ceras epicuticulares de tipo Strelitzia, el polen con mucho almidón, las paredes celulares sin lignificar e impregnadas con compuestos ácidos fluorescentes a la luz UV (ácidos ferúlicos, diferúlicos y cumáricos), y las hojas con cuerpos de sílice (SiO2),[37] [38] [39] [40] [41] también los estomas para o tetracíticos, las inflorescencias rodeadas de brácteas y el embrión corto y ancho.[3]
- Dasypogonaceae Dumort.
- Arecales Bromhead
- Arecaceae Bercht. & J.Presl
- Commelinales Mirb. ex Bercht. & J.Presl
- Commelinaceae Mirb.
- Haemodoraceae R.Br.
- Hanguanaceae Airy Shaw
- Philydraceae Link
- Pontederiaceae Kunth
- Poales Small
- Anarthriaceae D.F.Cutler & Airy Shaw
- Bromeliaceae Juss.
- Centrolepidaceae Endl.
- Cyperaceae Juss.
- Ecdeiocoleaceae D.F.Cutler & Airy Shaw
- Eriocaulaceae Martinov
- Flagellariaceae Dumort.
- Joinvilleaceae Toml. & A.C.Sm.
- Juncaceae Juss.
- Mayacaceae Kunth
- Poaceae Barnhart
- Rapateaceae Dumort.
- Restionaceae R.Br.
- Thurniaceae Engl.
- §Typhaceae Juss. (incluye Sparganiaceae Hanin)
- Xyridaceae C.Agardh
- Zingiberales Griseb.
- Cannaceae Juss.
- Costaceae Nakai
- Heliconiaceae Vines
- Lowiaceae Ridl.
- Marantaceae R.Br.
- Musaceae Juss.
- Strelitziaceae Hutch.
- Zingiberaceae Martinov
Probable hermano de las eudicotiledóneas
- Ceratophyllales Link
- Ceratophyllaceae Gray
Eudicotyledoneae M.J. Donoghue, J.A. Doyle & P.D. Cantino[42]
Las eudicotiledóneas conforman un clado que incluye a la mayoría de las plantas antiguamente consideradas dentro del grupo parafilético de las dicotiledóneas. El término proviene del griego y significa "dicotiledóneas verdaderas" debido a que contiene a la mayoría de las plantas que habían sido llamadas anteriormente dicotiledóneas, y tienen caracteres de dicotiledóneas típicas. El clado se caracteriza por el polen tricolpado, por las flores cíclicas y la presencia diferenciada de las partes internas y externas del perianto (o sea, cáliz y corola).
- Ranunculales Juss. ex Bercht. & J.Presl
- Berberidaceae Juss.
- ºCircaeasteraceae Hutch. (incluye Kingdoniaceae Airy Shaw)
- Eupteleaceae K.Wilh.
- Lardizabalaceae R.Br.
- Menispermaceae Juss.
- ºpapaveraceae Juss. (incluye Fumariaceae Marquis, Pteridophyllaceae Nakai ex Reveal & Hoogland)
- Ranunculaceae Juss.
- Sabiaceae Blume
- Proteales Juss. ex Bercht. & J.Presl
- Nelumbonaceae A.Rich.
- ººPlatanaceae T.Lestib.
- ººProteaceae Juss.
- †Trochodendrales Takht. ex Cronquist
- ºTrochodendraceae Eichler (incluye Tetracentraceae A.C.Sm.)
- †Buxales Takht. ex Reveal
- ºBuxaceae Dumort. (incluye Didymelaceae Leandri)
- *Haptanthaceae C.Nelson
Gunneridae D.E. Soltis, P.S. Soltis & W.S. Judd[43]
Las gunnéridas son un gran grupo de eudicotiledóneas que habían sido nombradas informalmente como «núcleo de las eudicotiledóneas» o «eudicotiledóneas nucleares» (en inglés "core eudicots") por casi todos los trabajos sobre filogenia de las angiospermas. Comprenden un grupo extremadamente amplio y diverso de angiospermas, con una gran variabilidad en el hábito, morfología, química, distribución geográfica y otros atributos. La sistemática clásica, basada únicamente en información morfológica, no fue capaz de reconocer este grupo. De hecho, la circunscripción de las gunnéridas como un clado está basada en una fuerte evidencia a partir de los datos de análisis moleculares.[44] [45] [46] La composición del clado es la siguiente: Gunnerales, Berberidopsidales (incluyendo a Aextoxicaceae;[47] [48] Dilleniaceae, Santalales, Saxifragales, Vitaceae, y todos los clados de Asteridae, Caryophyllales y Rosidae. En otras palabras, el clado está compuesto por Gunnerales más la totalidad del clado de las pentapétalas.[43]
- Gunnerales Takht. ex Reveal
- ººGunneraceae Meisn.
- ººMyrothamnaceae Nied.
- Dilleniaceae Salisb.
- Saxifragales Bercht. & J.Presl
- Altingiaceae Horan.
- Aphanopetalaceae Doweld
- Cercidiphyllaceae Engl.
- Crassulaceae J.St.-Hil.
- Daphniphyllaceae Müll.-Arg.
- Grossulariaceae DC.
- ººHaloragaceae R.Br.
- Hamamelidaceae R.Br.
- ºIteaceae J.Agardh (incluye Pterostemonaceae Small)
- Paeoniaceae Raf.
- ººPenthoraceae Rydb. ex Britt.
- *§Peridiscaceae Kuhlm. (incluye Medusandraceae Brenan, Soyauxia Oliver)
- Saxifragaceae Juss.
- ººTetracarpaeaceae Nakai
- †Berberidopsidales Doweld
- Aextoxicaceae Engl. & Gilg
- Berberidopsidaceae Takht.
- Santalales R.Br. ex Bercht. & J.Presl
- *Balanophoraceae Rich.
- Loranthaceae Juss.
- Misodendraceae J.Agardh
- Santalaceae R.Br.
- Olacaceae R.Br.
- Opiliaceae Valeton
- *Schoepfiaceae Blume
- Caryophyllales Juss. ex Bercht. & J.Presl
- Achatocarpaceae Heimerl
- Aizoaceae Martinov
- Amaranthaceae Juss.
- *Anacampserotaceae Eggli & Nyffeler
- Ancistrocladaceae Planch. ex Walp.
- Asteropeiaceae Takht. ex Reveal & Hoogland
- Barbeuiaceae Nakai
- Basellaceae Raf.
- Cactaceae Juss.
- Caryophyllaceae Juss.
- §Didiereaceae Radlk.
- Dioncophyllaceae Airy Shaw
- Droseraceae Salisb.
- Drosophyllaceae Chrtek, Slavíková & Studnicka
- Frankeniaceae Desv.
- Gisekiaceae Nakai
- Halophytaceae A.Soriano
- *Limeaceae Shipunov ex Reveal
- *Lophiocarpaceae Doweld & Reveal
- §Molluginaceae Bartl.
- *Montiaceae Raf.
- Nepenthaceae Dumort.
- Nyctaginaceae Juss.
- Physenaceae Takht.
- Phytolaccaceae R.Br.
- Plumbaginaceae Juss.
- Polygonaceae Juss.
- §Portulacaceae Juss.
- Rhabdodendraceae Prance
- Sarcobataceae Behnke
- Simmondsiaceae Tiegh.
- Stegnospermataceae Nakai
- *Talinaceae Doweld
- Tamaricaceae Link
Rosidae Takhtajan [W.S. Judd, P.D. Cantino, D.E. Soltis & P.S. Soltis][49]
Las rósidas son un gran grupo de eudicotiledóneas que contiene aproximadamente 70.000 especies,[50] más de la cuarta parte del total de especies de las angiospermas.[51] Estos órdenes, a su vez, comprenden unas 140 familias.[52] Conjuntamente con las astéridas, constituyen los dos grupos más grandes de eudicotiledóneas. Las rósidas comprenden al orden Vitales y a dos grandes clados: las fábidas y las málvidas.[49]
- †Vitales Juss. ex Bercht. & J.Presl
- Vitaceae Juss.
Fabidae W.S. Judd, D.E. Soltis & P.S. Soltis[53]
Las fábidas son un clado de eudicotiledóneas que habían sido nombradas informalmente como «eurrósidas I»[54] [55] [56] o «fabids».[57] No se han descubierto para este clado sinapomorfias que no estén basadas en los datos moleculares.[53]
- †Zygophyllales Link
- ººKrameriaceae Dumort.
- ººZygophyllaceae R.Br.
- Celastrales Link
- ºCelastraceae R.Br. (incluye Lepuropetalaceae Nakai, Parnassiaceae Martinov, Pottingeriaceae Takht.)
- Lepidobotryaceae J.Léonard
- Oxalidales Bercht. & J.Presl
- Brunelliaceae Engl.
- Cephalotaceae Dumort.
- Connaraceae R.Br.
- Cunoniaceae R.Br.
- Elaeocarpaceae Juss. ex DC.
- *Huaceae A.Chev.
- Oxalidaceae R.Br.
- Malpighiales Juss. ex Bercht. & J.Presl
- Achariaceae Harms
- Balanopaceae Benth. & Hook.f.
- Bonnetiaceae L.Beauvis. ex Nakai
- *Calophyllaceae J.Agardh
- Caryocaraceae Voigt
- *Centroplacaceae Doweld & Reveal
- ººChrysobalanaceae R.Br.
- §Clusiaceae Lindl.
- Ctenolophonaceae Exell & Mendonça
- ººDichapetalaceae Baill.
- Elatinaceae Dumort.
- ºº§Erythroxylaceae Kunth (incluye Aneulophus Benth.)
- Euphorbiaceae Juss.
- ººEuphroniaceae Marc.-Berti
- Goupiaceae Miers
- Humiriaceae A.Juss.
- Hypericaceae Juss.
- Irvingiaceae Exell & Mendonça
- Ixonanthaceae Planch. ex Miq.
- Lacistemataceae Mart.
- Linaceae DC. ex Perleb
- Lophopyxidaceae H.Pfeiff.
- Malpighiaceae Juss.
- ºOchnaceae DC. (incluye Medusagynaceae Engl. & Gilg, Quiinaceae Choisy)
- Pandaceae Engl. & Gilg
- ºPassifloraceae Juss. ex Roussel (incluye Malesherbiaceae D.Don, Turneraceae Kunth ex DC.)
- Phyllanthaceae Martinov
- Picrodendraceae Small
- Podostemaceae Rich. ex Kunth
- Putranjivaceae Meisn.
- *Rafflesiaceae Dumort.
- ººRhizophoraceae Pers.
- Salicaceae Mirb.
- ººTrigoniaceae A.Juss.
- Violaceae Batsch
- Cucurbitales Juss. ex Bercht. & J.Presl
- Anisophylleaceae Ridl.
- Begoniaceae C.Agardh
- Coriariaceae DC.
- Corynocarpaceae Engl.
- Cucurbitaceae Juss.
- Datiscaceae Dumort.
- Tetramelaceae Airy Shaw
- Fabales Bromhead
- Fabaceae Lindl.
- Polygalaceae Hoffmanns. & Link
- Quillajaceae D.Don
- Surianaceae Arn.
- Fagales Engl.
- Betulaceae Gray
- Casuarinaceae R.Br.
- Fagaceae Dumort.
- §Juglandaceae DC. ex Perleb (incluye Rhoipteleaceae Hand.-Mazz.)
- Myricaceae A.Rich. ex Kunth
- Nothofagaceae Kuprian.
- Ticodendraceae Gómez-Laur. & L.D.Gómez
- Rosales Bercht. & J.Presl
- Barbeyaceae Rendle
- Cannabaceae Martinov
- Dirachmaceae Hutch.
- Elaeagnaceae Juss.
- Moraceae Gaudich.
- Rhamnaceae Juss.
- Rosaceae Juss.
- Ulmaceae Mirb.
- Urticaceae Juss.
Malvidae W.S. Judd, D.E. Soltis & P.S. Soltis[58]
Las málvidas[59] son un grupo de eudicotiledóneas que habían sido nombradas informalmente como «eurrósidas II» en varios trabajos sobre filogenia de las angiospermas[60] [61] [62] [63] o “malvids”.[64] El nombre eurrósidas II también había sido aplicado de un clado más amplio que incluía a las mirtales.[65] [58]
- Geraniales Juss. ex Bercht. & J.Presl
- ºGeraniaceae Juss. (incluye Hypseocharitaceae Wedd.)
- ºMelianthaceae Horan. (incluye Francoaceae A.Juss.)
- §Vivianiaceae Klotzsch (incluye Ledocarpaceae Meyen)
- Myrtales Juss. ex Bercht. & J.Presl
- Alzateaceae S.A.Graham
- Combretaceae R.Br.
- Crypteroniaceae A.DC.
- Lythraceae J.St.-Hil.
- ºMelastomataceae Juss. (incluye Memecylaceae DC.)
- §Myrtaceae Juss. (incluye Heteropyxidaceae Engl. & Gilg, Psiloxylaceae Croizat)
- Onagraceae Juss.
- §Penaeaceae Sweet ex Guill. (incluye Oliniaceae Arn., Rhynchocalycaceae L.A.S.Johnson & B.G.Briggs)
- Vochysiaceae A.St.-Hil.
- Crossosomatales Takht. ex Reveal
- *Aphloiaceae Takht.
- Crossosomataceae Engl.
- *Geissolomataceae A.DC.
- *Guamatelaceae S.Oh & D.Potter
- Stachyuraceae J.Agardh
- Staphyleaceae Martinov
- *§Strasburgeriaceae Soler. (incluye Ixerbaceae Griseb. ex Doweld & Reveal)
- †Picramniales Doweld
- *Picramniaceae Fernando & Quinn
- †Huerteales Doweld
- *Dipentodontaceae Merr.
- *Gerrardinaceae Alford
- Tapisciaceae Takht.
- Brassicales Bromhead
- ºAkaniaceae Stapf (incluye Bretschneideraceae Engl. & Gilg)
- Bataceae Mart. ex Perleb
- §Brassicaceae Burnett
- *Capparaceae Juss.
- Caricaceae Dumort.
- *Cleomaceae Bercht. & J.Presl
- Emblingiaceae J.Agardh
- Gyrostemonaceae A.Juss.
- Koeberliniaceae Engl.
- Limnanthaceae R.Br.
- Moringaceae Martinov
- Pentadiplandraceae Hutch. & Dalziel
- Resedaceae Martinov
- Salvadoraceae Lindl.
- Setchellanthaceae Iltis
- Tovariaceae Pax
- Tropaeolaceae Juss. ex DC.
- Malvales Juss. ex Bercht. & J.Presl
- ºBixaceae Kunth (incluye Cochlospermaceae Planch., Diegodendraceae Capuron)
- Cistaceae Juss.
- *Cytinaceae A.Rich.
- Dipterocarpaceae Blume
- Malvaceae Juss.
- Muntingiaceae C.Bayer, M.W.Chase & M.F.Fay
- Neuradaceae Kostel.
- Sarcolaenaceae Caruel
- Sphaerosepalaceae Tiegh. ex Bullock
- Thymelaeaceae Juss.
- Sapindales Juss. ex Bercht. & J.Presl
- Anacardiaceae R.Br.
- Biebersteiniaceae Schnizl.
- Burseraceae Kunth
- Kirkiaceae Takht.
- Meliaceae Juss.
- ºNitrariaceae Lindl. (incluye Peganaceae Tiegh. ex Takht., Tetradiclidaceae Takht.)
- Rutaceae Juss.
- Sapindaceae Juss.
- Simaroubaceae DC.
Asteridae Takhtajan [R.G. Olmstead & W.S. Judd][66]
Las astéridas son un gran grupo de eudicotiledóneas que incluye aproximadamente 80.000 especies, agrupadas en 13 órdenes y más de cien familias[50] y entre la tercera a la cuarta parte del total de especies de las angiospermas.[51] [52] Conjuntamente con las rósidas, constituyen los dos grupos más grandes de eudicotiledóneas. Las plantas pertenecientes a este clado se caracterizan por ser herbáceas, con flores hermafroditas, cigomorfas (de simetría bilateral), polinizadas por insectos, por los estambres dispuestos en un círculo y los pétalos de la corola unidos entre sí formando un tubo. El gineceo está formado por dos carpelos soldados. Las flores se organizan a menudo en inflorescencias apretadas, como las espigas de las labiadas y las plantagináceas o los capítulos (cabezuelas) de las Compuestas. Cuatro de las familias más grandes de angiospermas pertenecen a este clado: las asteráceas, las rubiáceas, las lamiáceas y las apocináceas.[3] [66]
- Cornales Link.
- Cornaceae Bercht. & J.Presl (incluye Nyssaceae Juss. ex Dumort.)
- Curtisiaceae Takht.
- Grubbiaceae Endl. ex Meisn.
- Hydrangeaceae Dumort.
- Hydrostachyaceae Engl.
- Loasaceae Juss.
- Ericales Bercht. & J.Presl
- Actinidiaceae Engl. & Gilg.
- Balsaminaceae A.Rich.
- Clethraceae Klotzsch
- Cyrillaceae Lindl.
- Diapensiaceae Lindl.
- Ebenaceae Gürke
- Ericaceae Juss.
- Fouquieriaceae DC.
- Lecythidaceae A.Rich.
- Marcgraviaceae Bercht. & J.Presl
- *Mitrastemonaceae Makino
- ºPentaphylacaceae Engl. (incluye Ternstroemiaceae Mirb. ex DC.)
- Polemoniaceae Juss.
- §Primulaceae Batsch ex Borkh. (incluye Maesaceae Anderb., B.Ståhl & Källersjö, Myrsinaceae R.Br., Theophrastaceae G.Don)
- Roridulaceae Martinov
- Sapotaceae Juss.
- Sarraceniaceae Dumort.
- ººSladeniaceae Airy Shaw
- Styracaceae DC. & Spreng.
- Symplocaceae Desf.
- ºTetrameristaceae Hutch. (incluye Pellicieraceae L.Beauvis.)
- Theaceae Mirb. ex Ker Gawl.
Lamiidae Takhtajan [R.G. Olmstead & W.S. Judd][67]
Las lámidas[68] son un grupo de eudicotiledóneas que habían sido nombradas informalmente como «euastéridas I».[69] [70] El clado, según ha sido circunscripto en APG III, comprende cuatro órdenes y cinco familias que todavía no han sido incluidas en ningún orden.[71] [17] La sinapomorfia más evidente de las lámidas es la presencia de vasos del xilema con perforaciones simples.[72] [67]
- §*Boraginaceae Juss. (incluye Hoplestigmataceae Gilg)
- Vahliaceae Dandy
- Icacinaceae Miers
- Metteniusaceae H.Karst. ex Schnizl.
- Oncothecaceae Kobuski ex Airy Shaw
- Garryales Lindl.
- Eucommiaceae Engl.
- ºGarryaceae Lindl. (incluye Aucubaceae Bercht. & J.Presl)
- Gentianales Juss. ex Bercht. & J.Presl
- Apocynaceae Juss.
- Gelsemiaceae Struwe & V.A.Albert
- Gentianaceae Juss.
- Loganiaceae R.Br. ex Mart.
- Rubiaceae Juss.
- Lamiales Bromhead
- §Acanthaceae Juss.
- Bignoniaceae Juss.
- Byblidaceae Domin
- Calceolariaceae Olmstead
- Carlemanniaceae Airy Shaw
- Gesneriaceae Rich. & Juss.
- Lamiaceae Martinov
- *Linderniaceae Borsch, K.Müll., & Eb.Fisch.
- Lentibulariaceae Rich.
- Martyniaceae Horan.
- Oleaceae Hoffmanns. & Link
- Orobanchaceae Vent.
- Paulowniaceae Nakai
- Pedaliaceae R.Br.
- Phrymaceae Schauer
- §Plantaginaceae Juss.
- Plocospermataceae Hutch.
- Schlegeliaceae Reveal
- Scrophulariaceae Juss.
- Stilbaceae Kunth
- Tetrachondraceae Wettst.
- *Thomandersiaceae Sreem.
- Verbenaceae J.St.-Hil.
- Solanales Juss. ex Bercht. & J.Presl
- Convolvulaceae Juss.
- Hydroleaceae R.Br. ex Edwards
- Montiniaceae Nakai
- Solanaceae Juss.
- Sphenocleaceae T.Baskerv.
Campanulidae M.J. Donoghue & P.D. Cantino[73]
Las campanúlidas son un grupo de eudicotiledóneas que habían sido nombradas informalmente como «euastéridas II».[74] [75] Algunos atributos son característicos del grupo: las hojas alternas, con márgenes serrados o dentados, las flores epíginas, con una fusión tardía de los pétalos, filamentos de los estambres libres y los frutos indehiscentes.[76] [77] [73]
- Aquifoliales Senft
- Aquifoliaceae Bercht. & J.Presl
- §Cardiopteridaceae Blume (incluye Leptaulaceae Tiegh.)
- Helwingiaceae Decne.
- Phyllonomaceae Small
- Stemonuraceae Kårehed
- Asterales Link
- Alseuosmiaceae Airy Shaw
- Argophyllaceae Takht.
- Asteraceae Bercht. & J.Presl
- Calyceraceae R.Br. ex Rich.
- ºCampanulaceae Juss. (incluye Lobeliaceae Juss.)
- Goodeniaceae R.Br.
- Menyanthaceae Dumort.
- Pentaphragmataceae J.Agardh
- Phellinaceae Takht.
- Rousseaceae DC.
- ºStylidiaceae R.Br. (incluye Donatiaceae B.Chandler)
- †Escalloniales R.Br.
- §Escalloniaceae R.Br. ex Dumort. (incluye Eremosynaceae Dandy, Polyosmaceae Blume, Tribelaceae Airy Shaw)
- †Bruniales Dumort.
- Bruniaceae R.Br. ex DC.
- §Columelliaceae D.Don (incluye Desfontainiaceae Endl.)
- †Paracryphiales Takht. ex Reveal
- §Paracryphiaceae Airy Shaw (incluye *Quintiniaceae Doweld, Sphenostemonaceae P.Royen & Airy Shaw)
- Dipsacales Juss. ex Bercht. & J.Presl
- Adoxaceae E.Mey.
- §Caprifoliaceae Juss. (incluye Diervillaceae Pyck, Dipsacaceae Juss., Linnaeaceae Backlund, Morinaceae Raf., Valerianaceae Batsch)
- Apiales Nakai
- Apiaceae Lindl.
- Araliaceae Juss.
- Griseliniaceae J.R.Forst. & G.Forst. ex A.Cunn.
- Myodocarpaceae Doweld
- Pennantiaceae J.Agardh
- Pittosporaceae R.Br.
- §Torricelliaceae Hu (incluye Aralidiaceae Philipson & B.C.Stone, Melanophyllaceae Takht. ex Airy Shaw)
Taxones con posición incierta
- Apodanthaceae Takht. (tres géneros)
- Cynomoriaceae Endl. ex Lindl.
- Gumillea Ruiz & Pav.
- Petenaea Lundell (probablemente en Malvales)
- Nicobariodendron (probablemente en Celastraceae).
Véase también
Notas
a. ↑ Adicionalmente, existen seis familias con una posición filogenética incierta, las que no se muestran en el cladograma pero que aparecen listadas en la sección «Taxones con posición taxonómica incierta».
Referencias
- ↑ a b c d e f APG II. 2003. "An Update of the Angiosperm Phylogeny Group Classification for the orders and families of flowering plants: APG II". Botanical Journal of the Linnean Society. 141, 399-436 (pdf aquí)
- ↑ APG I Botanical Journal of the Linnean Society Volume 141 Issue 4 Page 399 - April 2003 doi:10.1046/j.1095-8339.2003.t01-1-00158.x
- ↑ a b c d Stevens, P. F. (2001 en adelante) Angiosperm Phylogeny Website Versión 9, Junio 2008 y actualizado desde entonces. http://www.mobot.org/MOBOT/research/APweb/ Fecha de acceso: 10 de mayo de 2010.
- ↑ a b Cantino, P.D. & de Queiroz, K. 2006. PhyloCode: a Phylogenetic Code of Biological Nomenclature. Version 3a. Website: http://www.phylocode.org. Revised June 16, 2006.
- ↑ De Queiroz, K. & Cantino, P.D. 2001. Phylogenetic nomenclature and the PhyloCode. Bull. Zool. Nomencl. 58: 254–271.
- ↑ De Queiroz, K. & Gauthier, J. 1990. Phylogeny as a central principle in taxonomy: phylogenetic definitions of taxon names. Syst. Zool. 39: 307–322.
- ↑ De Queiroz, K. & Gauthier, J. 1992. Phylogenetic taxonomy. Annual Rev. Ecol. Syst. 23: 449–480.
- ↑ Pickett, K.M. 2005. The new and improved PhyloCode, now with types, ranks, and even polyphyly: a conference report from the First International Phylogenetic Nomenclature Meeting. Cladistics 21 (2005) 79–82
- ↑ Cameron KM, Chase MW, Rudall PJ. Recircumscription of the monocotyledonous family Petrosaviaceae to include Japanolirion. Brittonia (2003) 55:214–225
- ↑ Merckx V, Bakker F, Huysmans S, Smets E. Bias and conflict in phylogenetic inference of myco-heterotrophic plants: a case study in Thismiaceae. Cladistics (2009) 25:64–77
- ↑ Smith SE, Read DJ. Mycorrhizal symbiosis (2008) 3rd edn. London: Academic Press
- ↑ Oliver, D. 1870. Gerrardina foliosa, Oliv. Hooker's Icon. Pl., Third Series 1: 60. t. 1075
- ↑ Milne-Redhead, E. 1939. Gerrardina eylesiana Milne-Redhead. Hooker's Icon. Pl., Fifth Series 4: 1-2, t. 3390.
- ↑ Alford, M. H. 2006. Gerrardinaceae: a new family of African flowering plants unresolved among Brassicales, Huerteales, Malvales, and Sapindales. Taxon 55: 959-964.
- ↑ Worberg, Andreas; Alford, Mac H.; Quandt, Dietmar; Borsch, Thomas. 2009. Huerteales sister to Brassicales plus Malvales, and newly circumscribed to include Dipentodon, Gerrardina, Huertea, Perrottetia, and Tapiscia. Taxon, Volume 58, Number 2, May 2009 , pp. 468-478(11)
- ↑ Valéry Malécot and Daniel L. Nickrent. Molecular Phylogenetic Relationships of Olacaceae and Related Santalales. Systematic Botany (2008), 33(1): pp. 97–106
- ↑ a b c d e f The Angiosperm Phylogeny Group III ("APG III", en orden alfabético: Brigitta Bremer, Kåre Bremer, Mark W. Chase, Michael F. Fay, James L. Reveal, Douglas E. Soltis, Pamela S. Soltis y Peter F. Stevens, además colaboraron Arne A. Anderberg, Michael J. Moore, Richard G. Olmstead, Paula J. Rudall, Kenneth J. Sytsma, David C. Tank, Kenneth Wurdack, Jenny Q.-Y. Xiang y Sue Zmarzty) (2009). «An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III.» (pdf). Botanical Journal of the Linnean Society (161): pp. 105-121. http://www3.interscience.wiley.com/journal/122630309/abstract.
- ↑ Angiosperm Phylogeny Group. (1998). «An ordinal classification for the families of flowering plants» (PDF). Ann. Missouri Bot. Gard. 85: pp. 531-553. http://www.bergianska.se/pub/publikationer/Bremer/Angiosperm_et_al_1998.pdf. Consultado el 26/12/2008.
- ↑ APG II. 2003. An Update of the Angiosperm Phylogeny Group Classification for the orders and families of flowering plants: APG II. Botanical Journal of the Linnean Society. 141, 399-436
- ↑ Chase, M. W.; Duvall, M. R., Hills, H. G., Conran, J. G., Cox, A. V., Eguiarte, L. E., Hartwell, J., Fay, M. F., Caddick, L. R., Cameron, K. M., y Hoot, S. (1995). «Molecular systematics of Lilianae.». En Rudall, P. J., Cribb, P. J., Cutler, D. F.. Monocotyledons: Systematics and evolution. (Royal Botanic Gardens edición). Kew. pp. 109-137.
- ↑ Chase, M.W.; Stevenson, D. W., Wilkin, P., y Rudall, P. J. (1995b). «Monocot systematics: a combined analysis.». En Rudall, P. J., Cribb, P. J., Cutler, D. F.. Monocotyledons: Systematics and evolution. (Royal Botanic Gardens edición). Kew. pp. 685-730.
- ↑ Fay, M. y Chase, M. 1996. Resurrection of Themidaceae for the Brodiaea alliance, and Recircumscription of Alliaceae, Amaryllidaceae and Agapanthoideae. Taxon 45: 441-451
- ↑ Fay, M. F. (2000). «Phylogenetic studies of Asparagales based on four plastid DNA regions.». En K. L. Wilson y D. A. Morrison. Monocots: Systematics and evolution. (Royal Botanic Gardens edición). Kollingwood, Australia: CSIRO. pp. 360-371.
- ↑ Pires, J. C.; Sytsma, K. J. (2002). «:A phylogenetic evaluation of a biosystematic framework: Brodiaea and related petaloid monocots (Themidaceae).». Amer. J. Bot. (89): pp. 1342-1359. http://www.amjbot.org/cgi/content/full/89/8/1342.
- ↑ THE ANGIOSPERM PHYLOGENY GROUP (Birgitta Bremer, Kåre Bremer, Mark W. Chase, Michael F. Fay, James L. Reveal, Douglas E. Soltis, Pamela S. Soltis and Peter F. Stevens, Arne A. Anderberg, Michael J. Moore, Richard G. Olmstead, Paula J. Rudall, Kenneth J. Sytsma, David C. Tank, Kenneth Wurdack, Jenny Q.-Y. Xiang & Sue Zmarzty). 2009. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III. Botanical Journal of the Linnean Society 161, 105–121.
- ↑ Meerow, A.W., Reveal, J.L., Snijman, D.A., Dutilh, J.H. Superconservation proposal to conserve Amaryllidaceae (1805) over Alliaceae (1797). Taxon 56: 1299-1300. 2007
- ↑ a b c Bronwyn Friedlander, Bryony Phillips & Tarryn Barrowman. 2009. As easy as APG III – Scientists revise the system of classifying flowering plants. Royal Botanical Gardens, Kew. Press Release.
- ↑ P.D. Cantino, J.A. Doyle, S.W. Graham, W. S. Judd, R.G. Olmstead, D. E. Soltis, P.S. Soltis & M.J. Donoghue. 2007. Towards a phylogenetic nomenclature of Tracheophyta. Taxon 56 (3) 822–846.
- ↑ Angiospermae Lindley 1830: xxxvi [P.D. Cantino & M.J. Donoghue], converted clade name.. En: P.D. Cantino, J.A. Doyle, S.W. Graham, W. S. Judd, R.G. Olmstead, D. E. Soltis, P.S. Soltis & M.J. Donoghue. 2007. Towards a phylogenetic nomenclature of Tracheophyta. Taxon 56 (3) 822–846. Pag. E 22.
- ↑ a b Magnoliidae Novák ex Takhtajan 1967: 51 [W.S. Judd, P.S. Soltis & D.E. Soltis], converted clade name. En: P.D. Cantino, J.A. Doyle, S.W. Graham, W. S. Judd, R.G. Olmstead, D. E. Soltis, P.S. Soltis & M.J. Donoghue. 2007. Towards a phylogenetic nomenclature of Tracheophyta. Taxon 56 (3) 822–846. Pag. E 24.
- ↑ Simpson, M.G. 2006. Plant Systematics. Elsevier, Amsterdam
- ↑ Qiu, Y.-L., Li, L., Hendry, T.A., Li, R., Taylor, D.W., Issa, M.J., Ronen, A.J., Vekaria, M.L. & White, A.M. 2006. Reconstructing the basal angiosperm phylogeny: evaluating information content of mitochondrial genes. Taxon 55: 837-856.
- ↑ Soltis, D. E., P. S. Soltis, M. W. Chase, M. E. Mort, D. C. Albach, M. Zanis, V. Savolainen, W. J. Hahn, S. B. Hoot, M. F. Fay, M. Axtell, S. M. Swensen, L. M. Prince, W. J. Kress, K. C. Nixon, and J. S. Farris. 2000. Angiosperm phylogeny inferred from 18S rDNA, rbcL, and atpB sequences. Botanical Journal of the Linnean Society 133: 381-461.
- ↑ P.D. Cantino, J.A. Doyle, S.W. Graham, W. S. Judd, R.G. Olmstead, D. E. Soltis, P.S. Soltis & M.J. Donoghue. 2007. Towards a phylogenetic nomenclature of Tracheophyta. Taxon 56 (3) 822–846
- ↑ Monocotyledoneae de Candolle 1817: 122 [W.S. Judd, P.S. Soltis, D.E. Soltis & S.W. Graham], converted clade name. En: P.D. Cantino, J.A. Doyle, S.W. Graham, W. S. Judd, R.G. Olmstead, D. E. Soltis, P.S. Soltis & M.J. Donoghue. 2007. Towards a phylogenetic nomenclature of Tracheophyta. Taxon 56 (3) 822–846. Pag. E 24.
- ↑ Commelinidae Takhtajan 1967: 514 [S.W. Graham & W.S. Judd], converted clade name. En: P.D. Cantino, J.A. Doyle, S.W. Graham, W. S. Judd, R.G. Olmstead, D. E. Soltis, P.S. Soltis & M.J. Donoghue. 2007. Towards a phylogenetic nomenclature of Tracheophyta. Taxon 56 (3) 822–846. Pag. E 25.
- ↑ Dahlgren, R. M.; Clifford, H. T., Yeo, P. F. (1985). The families of the monocotyledons. (Springer-Verlag edición). Berlín.
- ↑ Harley, M. M.; Ferguson, I. K. (1990). «The role of the SEM in pollen morphology and plant systematics.». En D. Claugher. Scanning electron microscope in taxonomy and functional morphology. Systematics Association Special Vol. 41.. Oxford.: Clarendon Press. pp. 45-68.
- ↑ Barthlott, W.; Fröhlich, D. (1983). «
- Micromorphologie und Orientierungs muster epicuticularer Wachs-Kristalloide: ein neues systematisches Merkmal bei Monokotylen.». Plant Syst. Evol. (142): pp. 171-185.
- ↑ Harris, P. J.; Hartley, R. D. (1980). «
- Phenolic constituents of the cell walls of monocotyledons.». Biochem. Syst. Ecol. (8): pp. 153-160.
- ↑ Zona, S. (2001). «:Starchy pollen in commelinoid monocots.» (pdf). Ann. Bot. II (87): pp. 109-116. http://aob.oxfordjournals.org/cgi/reprint/87/1/109. Consultado el 25/02/2008.
- ↑ Eudicotyledoneae M.J. Donoghue, J.A. Doyle & P.D. Cantino, new clade name. En: P.D. Cantino, J.A. Doyle, S.W. Graham, W. S. Judd, R.G. Olmstead, D. E. Soltis, P.S. Soltis & M.J. Donoghue. 2007. Towards a phylogenetic nomenclature of Tracheophyta. Taxon 56 (3) 822–846. Pag. E 26.
- ↑ a b Gunneridae D.E. Soltis, P.S. Soltis & W.S. Judd, new clade name. En: P.D. Cantino, J.A. Doyle, S.W. Graham, W. S. Judd, R.G. Olmstead, D. E. Soltis, P.S. Soltis & M.J. Donoghue. 2007. Towards a phylogenetic nomenclature of Tracheophyta. Taxon 56 (3) 822–846. Pag. E 27.
- ↑ Hoot, S. B., S. Magallón, and P. R. Crane. 1999. Phylogeny of basal eudicots based on three molecular data sets: atpB, rbcL, and 18S nuclear ribosomal DNA sequences. Annals of the Missouri Botanical Garden 86: 1-32
- ↑ Savolainen, V., M. W. Chase, C. M. Morton, D. E. Soltis, C. Bayer, M. F. Fay, A. De Bruijn, S. Sullivan, and Y.-L. Qiu. 2000a. Phylogenetics of flowering plants based upon a combined analysis of plastid atpB and rbcL gene sequences. Systematic Biology 49: 306-362
- ↑ Soltis, D. E., P. S. Soltis, M. W. Chase, M. E. Mort, D. C. Albach, M. Zanis, V. Savolainen, W. J. Hahn, S. B. Hoot, M. F. Fay, M. Axtell, S. M. Swensen, L. M. Prince, W. J. Kress, K. C. Nixon, and J. S. Farris. 2000. Angiosperm phylogeny inferred from 18S rDNA, rbcL, and atpB sequences. Botanical Journal of the Linnean Society 133: 381-461.
- ↑ Hilu, K.W., Borsch, T., Müller, K., Soltis, D.E., Soltis, P.S., Savolainen, V., Chase, M.W., Powell, M.P., Alice, L.A., Evans, R., Sauquet, H., Neinhuis, C., Slotta, T.A.B., Rohwer, J.G., Campbell, C.S. & Chatrou, L.W. 2003. Angiosperm phylogeny based on matK sequence information. Amer. J. Bot. 90: 1758–1776.
- ↑ Soltis, D.E., Soltis, P.S., Chase, M.W., Mort, M.E., Albach, D.C., Zanis, M., Savolainen, V., Hahn, W.H., Hoot, S.B., Fay, M.F., Axtell, M., Swensen, S.M., Prince, L.M., Kress, W.J., Nixon, K.C. & Farris, J.S. 2000. Angiosperm phylogeny inferred from 18S rDNA, rbcL, and atpB sequences. Bot. J. Linn. Soc. 133: 381–461.
- ↑ a b Rosidae Takhtajan 1967: 264 [W.S. Judd, P.D. Cantino, D.E. Soltis & P.S. Soltis], converted clade name. En: P.D. Cantino, J.A. Doyle, S.W. Graham, W. S. Judd, R.G. Olmstead, D. E. Soltis, P.S. Soltis & M.J. Donoghue. 2007. Towards a phylogenetic nomenclature of Tracheophyta. Taxon 56 (3) 822–846. Pag. E 27.
- ↑ a b Birgitta Bremer, Kare Bremer,Nahid Heidari,Per Erixon, Richard G. Olmstead, Arne A. Anderberg, Mari Kallersjo, and Edit Barkhordariana (10Mar2009), «Phylogenetics of asterids based on 3 coding and 3 non-coding chloroplast DNA markers and the utility of non-coding DNA at higher taxonomic levels», Molecular Phylogenetics and Evolution 106 (10): 3853–3858, http://www.bergianska.se/pub/publikationer/Bremer/Bremer_etal_6-gen_2002.pdf
- ↑ a b Robert W. Scotland and Alexandra H. Wortley (2003), «How many species of seed plants are there?», Taxon 52 (1): 101–104, doi:
- ↑ a b Douglas E. Soltis, Pamela S. Soltis, Peter K. Endress, and Mark W. Chase (2005), Phylogeny and Evolution of the Angiosperms, Sunderland, MA, USA: Sinauer, ISBN 978-0878938179
- ↑ a b Fabidae W.S. Judd, D.E. Soltis & P.S. Soltis, new clade name. En: P.D. Cantino, J.A. Doyle, S.W. Graham, W. S. Judd, R.G. Olmstead, D. E. Soltis, P.S. Soltis & M.J. Donoghue. 2007. Towards a phylogenetic nomenclature of Tracheophyta. Taxon 56 (3) 822–846. Pag. E 29.
- ↑ Savolainen, V., M. W. Chase, C. M. Morton, D. E. Soltis, C. Bayer, M. F. Fay, A. De Bruijn, S. Sullivan, and Y.-L. Qiu. 2000a. Phylogenetics of flowering plants based upon a combined analysis of plastid atpB and rbcL gene sequences. Systematic Biology 49: 306-362
- ↑ Savolainen, V., Fay, M.F., Albach, D.C., Bachlund, A., van der Bank, M., Cameron, K.M., Johnson, S.A., Lledo, M.D., Pintaud, J.-C., Powell, M., Sheahan, M.C., Soltis, D.E., Soltis, P.S., Weston, P., Whitten, W.M., Wurdack, K.J. & Chase, M.W. 2000b. Phylogeny of the eudicots: a nearly complete familial analysis based on rbcL gene sequences. Kew Bull. 55: 257-309.
- ↑ Soltis, D. E., P. S. Soltis, M. W. Chase, M. E. Mort, D. C. Albach, M. Zanis, V. Savolainen, W. J. Hahn, S. B. Hoot, M. F. Fay, M. Axtell, S. M. Swensen, L. M. Prince, W. J. Kress, K. C. Nixon, and J. S. Farris. 2000. Angiosperm phylogeny inferred from 18S rDNA, rbcL, and atpB sequences. Botanical Journal of the Linnean Society 133: 381-461.
- ↑ Judd, W.S. & Olmstead, R.G. 2004. A survey of tricolpate (eudicot) phylogenetic relationships. Amer. J. Bot. 91:1627-1644.
- ↑ a b Malvidae W.S. Judd, D.E. Soltis & P.S. Soltis, new clade name. En: P.D. Cantino, J.A. Doyle, S.W. Graham, W. S. Judd, R.G. Olmstead, D. E. Soltis, P.S. Soltis & M.J. Donoghue. 2007. Towards a phylogenetic nomenclature of Tracheophyta. Taxon 56 (3) 822–846. Pag. E 29.
- ↑ Malvidae D.E. Soltis, P.S. Soltis & W.S. Judd. En: P.D. Cantino, J.A. Doyle, S.W. Graham, W. S. Judd, R.G. Olmstead, D. E. Soltis, P.S. Soltis & M.J. Donoghue. 2007. Towards a phylogenetic nomenclature of Tracheophyta. Taxon 56 (3) 822–846
- ↑ Soltis, D.E., Soltis, P.S., Chase, M.W., Mort, M.E., Albach, D.C., Zanis, M., Savolainen, V., Hahn, W.H., Hoot, S.B., Fay, M.F., Axtell, M., Swensen, S.M., Prince, L.M., Kress, W.J., Nixon, K.C. & Farris, J.S. 2000. Angiosperm phylogeny inferred from 18S rDNA, rbcL, and atpB sequences. Bot. J. Linn. Soc. 133: 381–461.
- ↑ Soltis, D.E., Soltis, P.S., Endress, P.K. & Chase, M.W. 2005. Phylogeny and Evolution of Angiosperms. Sinauer Associates, Sunderland.
- ↑ Judd, W.S., Campbell, C.S., Kellogg, E.A., Stevens, P.F. & Donoghue, M.J. 2002. Plant Systematics—A Phylogenetic Approach, 2nd ed. Sinauer Associates, Sunderland.
- ↑ APG II (Angiosperm Phylogeny Group II). 2003. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG II. Bot. J. Linn. Soc. 141: 399–436.
- ↑ Judd, W.S. & Olmstead, R.G. 2004. A survey of tricolpate (eudicot) phylogenetic relationships. Amer. J. Bot. 91: 1627–1644 ).
- ↑ Savolainen, V., Chase, M.W., Hoot, S.B., Morton, C.M., Soltis, D.E., Bayer, C., Fay, M.F., de Bruijn, A.Y., Sullivan, S. & Qiu, Y.-L. 2000a. Phylogenetics of flowering plants based on combined analysis of plastid atpB and rbcL gene sequences. Syst. Biol. 49: 306–362.
- ↑ a b Asteridae Takhtajan 1967: 405 [R.G. Olmstead & W.S. Judd], converted clade name. En: P.D. Cantino, J.A. Doyle, S.W. Graham, W. S. Judd, R.G. Olmstead, D. E. Soltis, P.S. Soltis & M.J. Donoghue. 2007. Towards a phylogenetic nomenclature of Tracheophyta. Taxon 56 (3) 822–846. Pag. E 29.
- ↑ a b Lamiidae Takhtajan 1987: 228 [R.G. Olmstead & W.S. Judd], converted clade name. En: P.D. Cantino, J.A. Doyle, S.W. Graham, W. S. Judd, R.G. Olmstead, D. E. Soltis, P.S. Soltis & M.J. Donoghue. 2007. Towards a phylogenetic nomenclature of Tracheophyta. Taxon 56 (3) 822–846. Pag. E 29.
- ↑ Lamiidae R.G. Olmstead & W.S. Judd. En: P.D. Cantino, J.A. Doyle, S.W. Graham, W. S. Judd, R.G. Olmstead, D. E. Soltis, P.S. Soltis & M.J. Donoghue. 2007. Towards a phylogenetic nomenclature of Tracheophyta. Taxon 56 (3) 822–846
- ↑ Savolainen, V., M. W. Chase, C. M. Morton, D. E. Soltis, C. Bayer, M. F. Fay, A. De Bruijn, S. Sullivan, and Y.-L. Qiu. 2000a. Phylogenetics of flowering plants based upon a combined analysis of plastid atpB and rbcL gene sequences. Systematic Biology 49: 306-362
- ↑ Soltis, D. E., P. S. Soltis, M. W. Chase, M. E. Mort, D. C. Albach, M. Zanis, V. Savolainen, W. J. Hahn, S. B. Hoot, M. F. Fay, M. Axtell, S. M. Swensen, L. M. Prince, W. J. Kress, K. C. Nixon, and J. S. Farris. 2000. Angiosperm phylogeny inferred from 18S rDNA, rbcL, and atpB sequences. Botanical Journal of the Linnean Society 133: 381-461.
- ↑ P.D. Cantino, J.A. Doyle, S.W. Graham, W. S. Judd, R.G. Olmstead, D. E. Soltis, P.S. Soltis & M.J. Donoghue. 2007. Towards a phylogenetic nomenclature of Tracheophyta. Taxon 56 (3) 822–846
- ↑ Baas, P., Jansen, S. & Wheeler, E.A. 2003. Ecological adaptations and deep phylogenetic splits-evidence and questions from the secondary xylem. Pp. 221-239 En: Stuessy, T.E, Mayer, V. & Horandl, E. (eds.), Deep Morphology: Toward a Renaissance of Morphology in Plant Systematics. A.R.G. Gantner Verlag, Liechtenstein. [Regnum Veg. 141]
- ↑ a b Campanulidae M.J. Donoghue & P.D. Cantino, new clade name. En: P.D. Cantino, J.A. Doyle, S.W. Graham, W. S. Judd, R.G. Olmstead, D. E. Soltis, P.S. Soltis & M.J. Donoghue. 2007. Towards a phylogenetic nomenclature of Tracheophyta. Taxon 56 (3) 822–846. Pag. E 30.
- ↑ Savolainen, V., M. W. Chase, C. M. Morton, D. E. Soltis, C. Bayer, M. F. Fay, A. De Bruijn, S. Sullivan, and Y.-L. Qiu. 2000a. Phylogenetics of flowering plants based upon a combined analysis of plastid atpB and rbcL gene sequences. Systematic Biology 49: 306-362
- ↑ Soltis, D. E., P. S. Soltis, M. W. Chase, M. E. Mort, D. C. Albach, M. Zanis, V. Savolainen, W. J. Hahn, S. B. Hoot, M. F. Fay, M. Axtell, S. M. Swensen, L. M. Prince, W. J. Kress, K. C. Nixon, and J. S. Farris. 2000. Angiosperm phylogeny inferred from 18S rDNA, rbcL, and atpB sequences. Botanical Journal of the Linnean Society 133: 381-461.
- ↑ Judd, W. S., C. S. Campbell, E. A. Kellogg, P. F. Stevens, and M. J. Donoghue. 2002. Plant systematics: a phylogenetic approach. Sinauer Associates, Inc., Sunderland, Massachusetts, USA.
- ↑ Bremer, K., A. Backlund, B. Sennblad, U. Swenson, K. Andreasen, M. Hjertson, J. Lundberg, M. Backlund, and B. Bremer. 2001. A phylogenetic analysis of 100+ genera and 50+ families of euasterids based on morphological and molecular data with notes on possible higher level morphological synapomorphies. Plant Systematics and Evolution 229: 137-169.
Bibliografía
- The Angiosperm Phylogeny Group III ("APG III", en orden alfabético: Brigitta Bremer, Kåre Bremer, Mark W. Chase, Michael F. Fay, James L. Reveal, Douglas E. Soltis, Pamela S. Soltis y Peter F. Stevens, además colaboraron Arne A. Anderberg, Michael J. Moore, Richard G. Olmstead, Paula J. Rudall, Kenneth J. Sytsma, David C. Tank, Kenneth Wurdack, Jenny Q.-Y. Xiang y Sue Zmarzty) (2009). «An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III.» (pdf). Botanical Journal of the Linnean Society (161): pp. 105-121. http://www3.interscience.wiley.com/journal/122630309/abstract.
Categorías:- Sistemas botánicos
- Teoría de la información
- clado angiospermas






Wikimedia foundation. 2010.