- Autoincompatibilidad
-
Autoincompatibilidad

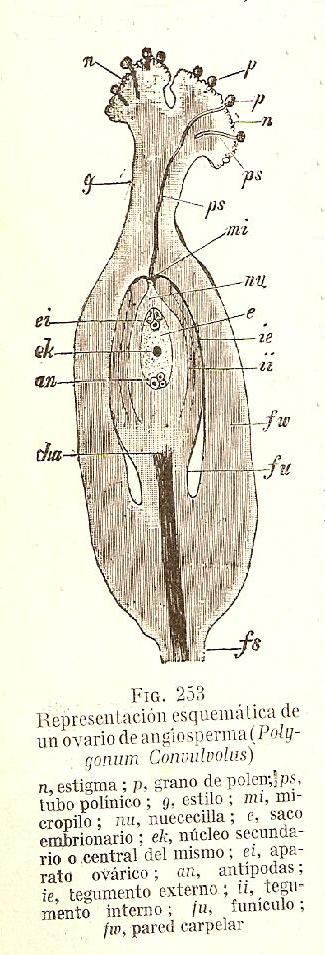
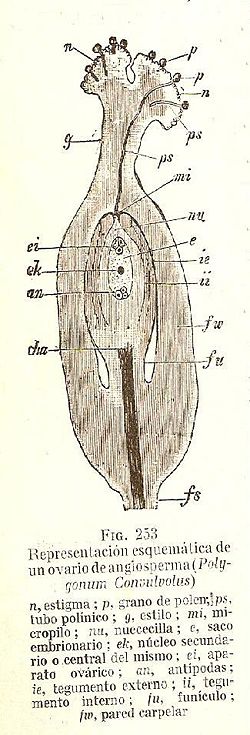 La fecundación en las Angiospermas es un proceso complejo que involucra varias etapas sucesivas. En el esquema se muestra como los granos de polen (p) aterrizan en el estigma de la flor (n) y germinan, emitiendo un tubo polínico (ps). El tubo polínico se elonga dentro del estilo (g) y atraviesa el micrópilo (mi). Ya dentro del óvulo, se orienta hacia el saco embrionario (e) para descargar los núcleos generativos que se fusionaran con la oósfera (la célula central de las 3 señaladas como (ei)) y con los núcleos polares (ek), produciéndose de ese modo la doble fecundación característica de las Angiospermas. La autoincompatibilidad impide la elongación de los tubos polínicos en algún punto de su trayecto hasta la oósfera.
La fecundación en las Angiospermas es un proceso complejo que involucra varias etapas sucesivas. En el esquema se muestra como los granos de polen (p) aterrizan en el estigma de la flor (n) y germinan, emitiendo un tubo polínico (ps). El tubo polínico se elonga dentro del estilo (g) y atraviesa el micrópilo (mi). Ya dentro del óvulo, se orienta hacia el saco embrionario (e) para descargar los núcleos generativos que se fusionaran con la oósfera (la célula central de las 3 señaladas como (ei)) y con los núcleos polares (ek), produciéndose de ese modo la doble fecundación característica de las Angiospermas. La autoincompatibilidad impide la elongación de los tubos polínicos en algún punto de su trayecto hasta la oósfera.
La autoincompatibilidad (AI) es la incapacidad de una planta hermafrodita para producir semillas por autopolinización aunque presente gametos viables. Es una estrategia reproductiva para promover la fecundación entre individuos que no estén relacionados y, por ende, es un mecanismo creador de nueva variabilidad genética.[1]
Durante la evolución de las angiospermas la AI ha surgido en varias ocasiones, en linajes totalmente diferentes. Más de 100 familias botánicas, entre las cuales se incluyen las Solanáceas, Poáceas, Asteráceas, Brasicáceas, Rosáceas y Fabáceas, presentan especies autoincompatibles. De hecho, se ha estimado que el 39% de las especies de angiospermas son AI.[2] Tan amplia distribución taxonómica es congruente con la existencia de varios mecanismos genéticos diferentes que regulan la AI, dependiendo de la familia considerada.
No obstante, en todos los casos se observa un mismo fenómeno: los granos de polen que llegan al estigma de la misma planta (o de plantas genéticamente relacionadas) son incapaces de efectuar la fecundación ya que detienen su desarrollo en alguna de las etapas del proceso (germinación de los granos de polen, desarrollo de los tubos polínicos en los pistilos o fertilización de la oósfera). Como consecuencia de este impedimento no se producen semillas tras la autopolinización.[1] Esta detención del crecimiento o desarrollo de los tubos polínicos dentro de los pistilos involucra, por un lado, el reconocimiento por parte del pistilo de cuáles son los tubos polínicos del mismo individuo y cuales son los de otras plantas. Por otro lado, tras ocurrir este reconocimiento, el desarrollo de los tubos polínicos propios (o de plantas genéticamente relacionadas) debe ser interrumpido. Ambos fenómenos, el reconocimiento y la interrupción del crecimiento, han sido estudiados extensivamente tanto a nivel citológico, fisiológico, genético y molecular.[3] [4]
Como se expresó previamente, los sistemas de AI en las plantas superiores han evolucionado varias veces en linajes independientes. A nivel molecular existen tres sistemas adecuadamente caracterizados: el sistema de autoincompatibilidad de las Solanáceas y Rosáceas; el de la amapola (Papaver) y el de Brassica. En dos de tales sistemas, se conocen en la actualidad los genes que codifican los componentes de reconocimiento polen-pistilo lo que ha demostrado claramente que las proteínas del pistilo y del polen involucradas en el reconocimiento son distintas entre si y que la AI es un mecanismo de "llave-cerradura" a nivel molecular.[5] [3]
Contenido
Tipos de mecanismos
Cuando las flores de la especie que presenta autoincompatibilidad son morfológicamente idénticas entre sí, se dice que la autoincompatibilidad es homomórfica. Por el contrario, cuando en la especie se presentan dos o tres tipos morfológicos diferentes de flores, se dice que la autoincompatibilidad es heteromórfica y el mecanismo se denomina heterostilia.[1]
Autoincompatibilidad homomórfica
El mecanismo mejor estudiado de AI en plantas actúa a través de la inhibición de la germinación o del crecimiento de los tubos polínicos en el estigma o en el estilo. Este mecanismo está basado en interacciones entre proteínas producidas por un único locus llamado S (del inglés Self-incompatibility, o sea, autoincompatibilidad). El locus S en realidad contiene 2 genes básicos, uno de los cuales se expresa en el pistilo y el otro en la antera o en los granos de polen. Se hace referencia a tales genes como determinantes femenino y masculino, respectivamente. Estos genes se hallan físicamente próximos en el genoma, genéticamente ligados y se los considera como un único alelo. Las proteínas producidas por los determinantes masculino y femenino del mismo alelo interactúan entre ellas en el momento de la germinación del grano de polen en el estigma y llevan a la detención del crecimiento del tubo polínico. En otras palabras, la identidad de las proteínas del grano de polen y del estigma genera una respuesta de AI que detiene el proceso de fertilización desde el inicio del mismo. Cuando, por el contrario, un determinante femenino interactúa con un determinante masculino proveniente de un alelo de AI diferente, no hay respuesta de AI y se produce la fertilización. Esta breve descripción de la respuesta de AI en plantas es usualmente más complicada y, muchas veces, están involucrados más de un locus de AI. Así, en las gramíneas, existen dos loci que gobiernan la reacción de AI denominados S y Z.[6] Existen dos tipos de incompatibilidad homómorfica, la AI gametofítica y la AI esporofítica, las cuales se describen a continuación.
Autoincompatibilidad gametofítica
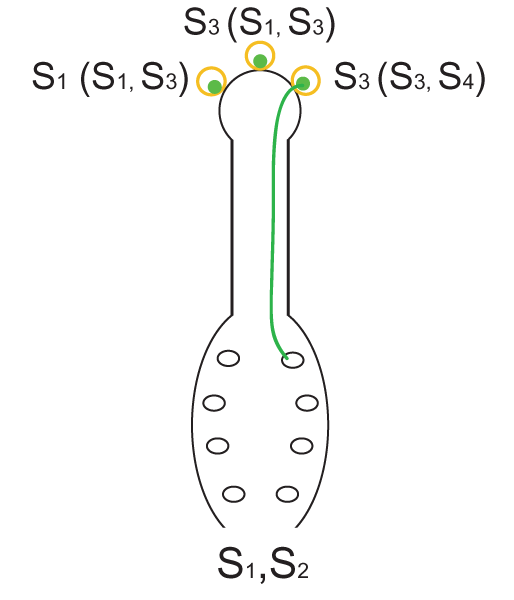
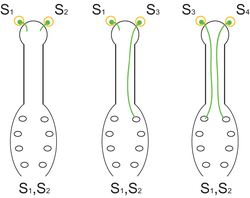 Fig. 1. El mecanismo de autoincompatibilidad gametofítica. La planta de genotipo S1S2 produce granos de polen con alelos S1 o S2, los cuales, al ser idénticos a los alelos S presentes en el estigma, no pueden germinar (izq.). Si sobre el estigma de una planta S1S2 llegan granos de polen de una planta con genotipo S1S3, el 50% de los mismos (aquellos con el alelo S3) podrán germinar y efectuar la polinización, la incompatibilidad se dice parcial (centro). Si sobre los estigmas de la planta de genotipo S1S2, finalmente, arriban granos de polen de otra planta con genotipo S3S4, la totalidad de los granos de polen podrán germinar ya que no hay identidad entre los alelos S de los granos de polen y del estigma (der.).
Fig. 1. El mecanismo de autoincompatibilidad gametofítica. La planta de genotipo S1S2 produce granos de polen con alelos S1 o S2, los cuales, al ser idénticos a los alelos S presentes en el estigma, no pueden germinar (izq.). Si sobre el estigma de una planta S1S2 llegan granos de polen de una planta con genotipo S1S3, el 50% de los mismos (aquellos con el alelo S3) podrán germinar y efectuar la polinización, la incompatibilidad se dice parcial (centro). Si sobre los estigmas de la planta de genotipo S1S2, finalmente, arriban granos de polen de otra planta con genotipo S3S4, la totalidad de los granos de polen podrán germinar ya que no hay identidad entre los alelos S de los granos de polen y del estigma (der.).El sistema de autoincompatibilidad gametofítica (AIG) es característico de varias familias de angiospermas, como por ejemplo Solanaceae, Rosaceae, Scrophulariaceae, Fabaceae, Onagraceae, Campanulaceae, Papaveraceae y Poaceae.[7] En la AIG, el fenotipo de AI del polen está determinado por su propio genotipo haploide, es decir, por la constitución genética del gametofito. En la figura 1 se describe como funciona este mecanismo mediante 3 ejemplos diferentes.
El mecanismo de la ARNasa
En este mecanismo, la elongación de los tubos polínicos es detenida cuando éstos han atravesado más o menos un tercio del pistilo.[8] El componente femenino de la reacción de AI es una ribonucleasa, denominada S-ARNasa,[9] la que aparentemente degrada el ARN ribosómico (ARNr) dentro de los tubos polínicos en el caso de que exista identidad entre los alelos S masculinos y femeninos. Al degradar el ARNr, el tubo polínico no puede sintetizar nuevas proteínas (y enzimas) por lo que su crecimiento se detiene en el tercio superior de los pistilos y termina colapsando.[8] El componente masculino de este sistema de AIG se ha identificado más tarde y se ha denominado proteína PiSLF, un miembro del grupo de proteínas con caja F.[10] Los miembros de este grupo de proteínas típicamente funcionan como ligasas de ubiquitina, o sea, "marcan" con ubiquitina a los productos correspondientes a alelos S distintos para dirigir su degradación en el proteosoma.[3] En las Solanáceas el locus S multialélico es altamente polimórfico y codifica para una serie alélica de S-ARNasas. La diversidad alélica intra e inter-poblacional de este locus ha sido examinada utilizando aproximaciones basadas en PCR. Tal aproximación, además, ha sido utilizada para demostrar que las S-ARNasas se hallan involucradas en la autoincompatibilidad observada en otras familias de angiospermas.[11] Las S-ARNasas muestran un patrón característico de 5 regiones conservadas y dos regiones hipervariables. Se considera que las primeras están relacionadas con la función catalítica de la enzima, mientras que las regiones hipervariables se hallan involucradas en la especificidad alélica.[12]
El mecanismo de la glicoproteína S
Este mecanismo, en el cual el crecimiento del tubo polínico es inhibido a los pocos minutos de ponerse en contacto con el estigma, ha sido descrito en detalle en la especie Papaver rhoeas (Papaveraceae).[8] El determinante femenino de la AI es una pequeña molécula extracelular que se expresa solamente en el estigma. El determinante masculino es, con toda probabilidad, un receptor de membrana (o sea, una proteína integral de membrana que responde a la unión del determinante femenino desencadenando una cascada de reacciones metabólicas dentro de la célula), pero no ha sido comprobado fehacientemente.[8] La interacción entre los determinantes masculinos y femeninos transmite una señal dentro del tubo polínico que da como resultado un fuerte influjo de cationes de calcio dentro del mismo lo que interfiere con su patrón de elongación normal.[13] [14] [15] El influjo de iones de calcio detiene el crecimiento del tubo polínico dentro de los dos primeros minutos. En ese estado, la inhibición del crecimiento todavía es reversible. De hecho, la elongación puede reanudarse si se manipulan adecuadamente las condiciones experimentales, lo que da como resultado la fertilización de los óvulos.[8] De modo subsiguiente a la detención de la elongación de los tubos polínicos, la proteína citosólica "p26", una pirofosfatasa, se inhibe por fosforilación,[16] lo que posiblemente determina la detención de los procesos de síntesis de moléculas dentro del tubo polínico, los que son imprescindibles para su elongación, y una reorganización de los filamentos de actina en el citoesqueleto de la célula vegetativa del tubo polínico.[17] [18] A los 10 minutos de su llegada al estigma, el tubo polínico ha sufrido una serie de cambios que lo conducen irreversiblemente a su muerte. A las 3-4 horas de la polinización, comienza la fragmentación del ADN nuclear del polen[19] y, finalmente, a las 10-14 horas, la célula vegetativa del tubo polínico muere por apoptosis.[8] [20] [3]
Autoincompatibilidad esporofítica
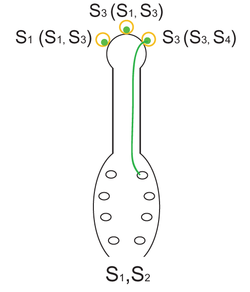 Fig.2. El mecanismo de autoincompatibilidad esporofítica. Los pistilos de la planta de genotipo S1S2 son polinizados con tres granos de polen, dos de ellos de genotipo S3 y el tercero de genotipo S1. La diferencia entre los dos granos de polen con genotipo S3 es que fueron producidos por dos plantas diferentes, una de genotipo S1S3, la otra de genotipo S3S4. Solamente pueden germinar en los estigmas, elongarse en los estilos y efectuar la fecundación los granos de polen que provienen de una planta que no posee alelos en común con el pistilo. Como la reacción de incompatibilidad depende del genotipo del esporofito que origina a los granos de polen y no del genotipo de los propios granos de polen, la reacción se denomina «esporofítica».
Fig.2. El mecanismo de autoincompatibilidad esporofítica. Los pistilos de la planta de genotipo S1S2 son polinizados con tres granos de polen, dos de ellos de genotipo S3 y el tercero de genotipo S1. La diferencia entre los dos granos de polen con genotipo S3 es que fueron producidos por dos plantas diferentes, una de genotipo S1S3, la otra de genotipo S3S4. Solamente pueden germinar en los estigmas, elongarse en los estilos y efectuar la fecundación los granos de polen que provienen de una planta que no posee alelos en común con el pistilo. Como la reacción de incompatibilidad depende del genotipo del esporofito que origina a los granos de polen y no del genotipo de los propios granos de polen, la reacción se denomina «esporofítica».En la autoincompatibilidad esporofítica (AIE) el fenotipo de AI de los granos de polen de una planta está determinado por el genotipo diploide de la antera (el esporófito) en la cual se originaron. Así, a diferencia del sistema de AIG en el cual cada grano de polen expresa su propio alelo de incompatibilidad, en este sistema cada grano de polen expresa los dos alelos de incompatibilidad presentes en la planta que lo originó. En otras palabras, en la AIG la reacción de incompatibilidad está determinada únicamente por el alelo presente en el gametofito, mientras que en la AIE queda gobernada por los dos alelos presentes en el esporófito. En esta diferencia sustancial radica, justamente, la denominación de ambos sistemas de AI. La AIE se ha identificado en las familias Brassicaceae, Asteraceae, Convolvulaceae, Betulaceae, Caryophyllaceae, Sterculiaceae y Polemoniaceae.[1] [21] En la figura 2 se describe mediante un esquema este mecanismo de incompatibilidad.
Solo uno de los muchos mecanismos de AIE ha sido descrito con detalle a nivel molecular, el de Brassica (Brassicaceae). Debido a que la AIE está determinada por un genotipo diploide, tanto el polen como el pistilo expresan los productos de la traducción de dos alelos diferentes, o sea, expresan dos determinantes femeninos y dos determinantes masculinos. Entre los dos alelos de un mismo locus de AIE existen relaciones de dominancia que determinan la existencia de patrones extremadamente complejos de compatiblidad/incompatibilidad. Estas relaciones de dominancia incluso permiten que se generen individuos homocigóticos para alelos S recesivos.[22] Comparada con una población en la cual los alelos S son codominantes, la presencia de relaciones de dominancia en la población eleva la probabilidad de que ocurran cruzamientos compatibles entre sus miembros.[22] La relación entre las frecuencias de los alelos S recesivos y dominantes refleja un balance dinámico. La relación entre las frecuencias de los alelos S recesivos y dominantes refleja un balance dinámico entre el aseguramiento de la reproducción (favorecido por los alelos recesivos) y el impedimento de la autofecundación (favorecido por los alelos dominantes).[23]
El mecanismo de AI en Brassica
Como se mencionó previamente, el fenotipo de AIE del polen está determinado por el genotipo diploide de la antera. En Brassica, la cubierta del polen, derivada del tapete de la antera, lleva los productos génicos de dos alelos S. Estos son proteínas pequeñas, ricas en cisteína. El gen que codifica estas proteínas se denomina SCR o SP11, y se expresa en el tapete de la antera (es decir, en el esporófito) como así también en la microspora y en el polen (o sea, en el gametofito).[24] [25] El determinante femenino de la respuesta de AI en Brassica es una proteína transmembranal denominada SRK, la cual presenta un dominio quinasa intracelular y un dominio extracelular variable.[26] [27] SRK se expresa en el estigma y probablemente funcione como un receptor de la proteína SCR/SP11 de la cubierta del polen. Otra proteína estigmática, denominada SLG, presenta una secuencia altamente similar a SRK y parece funcionar como un «coreceptor» del determinante masculino, amplificando la respuesta de autoincompatibilidad.[28] Las interacciones entre las proteínas SRK y SCR/SP11 dan como resultado la autofosforilación del dominio quinasa intracelular de SRK,[29] [30] y se transmite una señal dentro de la célula de la papila estigmática. Otra proteína esencial para la respuesta de autoincompatibilidad es MLPK, una quinasa serina-treonina, la cual se halla anclada del lado intracelular de la membrana plasmática.[31] Los eventos moleculares y celulares subsecuentes que llevan a la inhibición del crecimiento de los tubos polínicos todavía están pobremente descritos.[3]
Autoincompatibilidad heteromórfica
 Distilia en Primula vulgaris. Las flores Pin (A) presentan los filamentos de los estambres cortos y los pistilos largos. Las flores Thrum (B) muestran, en cambio, pistilos cortos y filamentos largos. Notas: 1.- pétalos, 2.-sépalos, 3.- anteras de los estambres y 4.- pistilo. Las únicas polinizaciones legítimas son las que se producen en cruzamientos Pin x Thrum o Thrum x Pin.
Distilia en Primula vulgaris. Las flores Pin (A) presentan los filamentos de los estambres cortos y los pistilos largos. Las flores Thrum (B) muestran, en cambio, pistilos cortos y filamentos largos. Notas: 1.- pétalos, 2.-sépalos, 3.- anteras de los estambres y 4.- pistilo. Las únicas polinizaciones legítimas son las que se producen en cruzamientos Pin x Thrum o Thrum x Pin.La heterostilia es una forma única de polimorfismo morfológico en las flores de ciertas especies que, como mecanismo, impide la autofecundación de cada flor. En las especies heterostílicas existen dos o tres tipos de flores (llamadas morfos). En una planta individual todas las flores presentan el mismo morfo. Estos morfos difieren cualitativamente entre sí en las longitudes de los pistilos y de los filamentos de las anteras. El fenotipo para cada morfo está genéticamente determinado. Cuando una especie presenta dos morfos florales se denomina distílica. En uno de los morfos, llamado Pin, los estambres son cortos y los pistilos largos. En el otro morfo, llamado Thrum, los estambres son largos y los pistilos cortos. En las especies que presentan heterostilia existe un mecanismo diferente de AI denominado autoincompatibilidad heteromórfica. Este mecanismo probablemente no se halle relacionado evolutivamente con los sistemas de incompatibilidad homomórfica.[32]
Genética de la autoincompatibilidad heteromórfica
Las especies del género Primula son posiblemente el ejemplo mejor conocido de flores heteromórficas y el mecanismo reproductivo que estas especies poseen ha atraído considerable atención entre botánicos, genetistas y evolucionistas, incluyendo a Charles Darwin ya para el año 1862.[33] Casi todas las especies heterostílicas presentan AI. Los loci responsables de la AI en estas especies están estrechamente ligados con aquellos genes responsables del polimorfismo floral, por lo que ambos caracteres se heredan en forma conjunta. La distilia está determinada por un solo gen con dos alelos, la tristilia por dos genes con dos alelos cada uno.[1]
La AI es esporofítica, o sea que los determinantes masculinos de la reacción de AI están gobernados por el genotipo del esporofito que produce los granos de polen. Los loci de AI casi siempre contienen dos alelos en la población de plantas, uno de los cuales es dominante sobre el otro tanto en el polen como en el pistilo. El polimorfismo en los alelos de AI se corresponde con el que gobierna la morfología floral, por lo que el polen de un morfo puede fertilizar solo los pistilos del otro morfo. En las especies con flores tristilias, cada flor contiene dos tipos de estambres, cada uno de los cuales produce polen capaz de fertilizar solo uno de los otros dos morfos de la población.[32] Las poblaciones de las especie distílicas contienen solo dos genotipos para el locus de AI: ss y Ss. La fertilización solo es posible entre plantas que presenten distinto genotipo para este locus, por lo que los cruzamientos legítimos (aquellos que producirán descendientes) son siempre Ss x ss o ss x Ss.[32] Esta restricción mantiene constantemente una relación de 1:1 entre ambos genotipos en la población, los que se hallan aleatoriamente distribuidos en el espacio que ocupa la población.[34] [35] En 2005 se logró determinar la secuencia de bases de uno de los alelos S de Primula vulgaris, la cual demostró ser altamente similar a la secuencias de alelos S de otras especies AI.[36]
Las especies tristílicas presentan, además de locus S, otro locus con solo dos alelos denominado M.[32] En este caso el número de genotipos posibles es mayor que en el caso de la distilia pero siempre se mantiene una relación de 1:1 entre los individuos de cada tipo de AI.[37]
Otros mecanismos de autoincompatibilidad
Autoincompatibilidad retardada
 El tulipanero africano (Spathodea campanulata) es una especie con incompatilibidad retardada.
El tulipanero africano (Spathodea campanulata) es una especie con incompatilibidad retardada.Como se describió anteriormente, en la mayoría de las plantas autoincompatibles la inhibición del crecimiento de los tubos polínicos se produce en el estigma o en el estilo. Los mecanismos de AI que actúan en el ovario han sido descriptos como autoincompatibilidad retardada (AIR) y son bastante comunes en plantas.[38] Los sistemas de AIR pueden dividirse según el momento en que ocurre la inhibición de la auto-fertilización en las siguientes categorías:[38] [39]
- la inhibición de los tubos polínicos incompatibles ocurre antes de su llegada a los óvulos.
- la reacción de autoincompatibilidad ocurre por inhibición de la fertilización, dentro del óvulo.
- la autoincompatibilidad se produce por la inhibición del crecimiento del embrión. De hecho, este es un mecanismo de AIR post-cigótico debido a que la fertilización ya ha ocurrido.
Especies que exhiben este tipo de mecanismo de AI son Narcissus triandrus (Amaryllidaceae), algunas del género Asclepias (Asclepiadaceae) y Spathodea campanulata (Bignoniaceae).[40] [41] [42] [43] [44]
Autoincompatibilidad críptica
La autoincompatibilidad críptica (AIC) se ha descrito en un número limitado de especies. Existe evidencia, por ejemplo, que Silene vulgaris (Caryophyllaceae), Amsinckia grandiflora (Boraginaceae), Decodon verticillatus (Lythraceae) y Cheiranthus cheiri (Brassicaceae) presentan este mecanismo.[45] [46] [47] [48] En este tipo de autoincompatibilidad, la presencia simultánea en el estigma de polen extraño y polen de la misma planta da como resultado una mayor cantidad de progenie proveniente de la polinización con el polen extraño.[48] No obstante, y al contrario de lo que ocurre con los sistemas de autoincompatibilidad «completos» o «absolutos» descritos previamente, en la AIC puede existir fertilización y producción de descendencia en ausencia de polen extraño. De este modo se asegura la reproducción del individuo aunque éste se halle aislado de otro individuo de la misma especie. La AIC actúa, al menos en algunas de las especies estudiadas, en el estadio de elongación de los tubos polínicos dentro de los pistilos y lleva a una tasa de crecimiento o elongación mayor en los tubos polínicos provenientes del polen extraño, los que arriban a los óvulos y realizan la fertilización de las oósferas antes que los tubos polínicos provenientes del polen de la propia planta.[48] Los mecanismos celulares y moleculares determinantes de la AIC no han sido descriptos hasta abril de 2008. La fuerza de la respuesta de AIC puede ser definida como la relación existente entre el número de oósferas fertilizadas por polen extraño con respecto al número de oósferas fertilizadas por el propio polen de la planta cuando se aplica la misma cantidad de polen extraño como propio de la planta sobre los estigmas de la misma. Esta relación va desde 3,2 hasta 11,5 en los taxa estudiados hasta el momento.[49]
La protrusión del estigma como mecanismo de autoincompatibilidad y su base genética
 Estigmas protruídos en las flores de Solanum crispum. Los pistilos de cada flor se observan como columnas blanquecinas que surgen desde el centro del anillo que forman los estambres.
Estigmas protruídos en las flores de Solanum crispum. Los pistilos de cada flor se observan como columnas blanquecinas que surgen desde el centro del anillo que forman los estambres. Estigmas insertos en las flores de Solanum lycopersicum (tomate). Los pistilos de cada flor quedan dentro del anillo de estambres o sobresalen mínimamente.
Estigmas insertos en las flores de Solanum lycopersicum (tomate). Los pistilos de cada flor quedan dentro del anillo de estambres o sobresalen mínimamente.Las especies silvestres de tomate (Solanum lycopersicum) se diferencian entre sí por su modo de reproducción. Así, hay especies AI y, por lo tanto, son «alógamas». Por otro lado, hay especies que usualmente se autopolinizan y autofecundan, se dice que son autocompatibles (AC) y, por ende, autógamas. Entre ambos extremos, finalmente, hay especies que presentan poblaciones AI y AC, y otras que son usualmente AI pero pueden dar una proporción de semillas por autofecundación (se las llama alógamas facultativas).
El grado en que los estigmas se elongan por arriba de los estambres (variable que se denomina «protrusión de los estigmas») es un determinante central en la posibilidad de polinización cruzada y, por ende, de la alogamia en estas especies. Las especies relacionadas con el tomate que son AI obligadas o facultativas presentan flores con los estigmas muy protruídos. En contraste, otras especies tales como el tomate cultivado, presentan los estigmas insertos lo que promueve la autopolinización. Se ha observado que una región del genoma del tomate (un QTL, acrónimo inglés para designar un locus que determina o gobierna un carácter cuantitativo) denominada se2.1 es responsable de una gran proporción de la variabilidad fenotípica observada para este carácter y que las mutaciones en este locus están involucradas en la evolución desde la alogamia hacia la autogamia en estas especies. La localización de este QTL en el genoma del tomate ha permitido concluir que se2.1 es un locus complejo que comprende por lo menos 5 genes estrechamente ligados, uno que controla la longitud del estilo, 3 que controlan el tamaño de los estambres y el último que gobierna la dehiscencia de las anteras. Este grupo de genes representaría los vestigios de un complejo ancestral de genes co-adaptados que controlan el tipo de apareamiento en estas especies de Solanum.[50]
En 2007 se ha avanzado un paso más en la comprensión de este grupo de genes ya que se ha determinado la secuencia de bases del locus Style2.1, el gen responsable de la longitud del estilo. Este gen codifica un factor de transcripción que regula el alargamiento celular de los estilos en desarrollo. La transición desde la AI a la AC estuvo acompañada, más que por un cambio en la proteína STYLE2.1, por una mutación en el promotor del gen style2.1 que dio como resultado la disminución en la expresión de ese gen durante el desarrollo floral y, por ende, el acortamiento consecuente de la longitud de los estilos.[51]
La evolución de la autocompatibilidad (AC)
La AI frecuentemente se ha perdido durante la diversificación de las angiospermas, un fenómeno que explica el origen de la autocompatibilidad (es decir, de la aceptación del propio polen y, por ende, de la producción de semillas por autopolinización). Sin embargo, no existe evidencia alguna que lo contrario haya alguna vez ocurrido, es decir, una vez que en el transcurso de la evolución se pierde un sistema de AI, éste no se vuelve a recobrar. Se considera que la pérdida irreversible de los sistemas de AI ocurre debido a que la transición hacia la AC está acompañada por el colapso en la variabilidad del locus S y por la acumulación de mutaciones con pérdida de función en muchos de los loci involucrados en la respuesta de autoincompatibilidad. Esta asimetría en las transiciones entre uno y otro sistema implica que la frecuencia de la AI en las angiospermas está declinando, o bien, que la misma confiere una ventaja macroevolutiva.[2]
Referencias
- ↑ a b c d e Frankel, R. & Galun, E. (1977). Pollination mechanisms, reproduction, and plant breeding. Monographs on Theoretical and Applied Genetics no. 2. Springer-Verlag. Berlin. ISBN 0387079343.
- ↑ a b Igic, B., Russell Lande, & Joshua R. Kohn, (2008). «Loss of Self-Incompatibility and Its Evolutionary Consequences» Int. J. Plant Sci.. Vol. 169. n.º 1. pp. 93-104.
- ↑ a b c d e Franklin-Tong, Vernonica E. (Ed.) (2008). Self-Incompatibility in Flowering Plants. Evolution, Diversity, and Mechanisms. Springer-Verlag. Berlin. ISBN 978-3-540-68485-5.
- ↑ Boavida LC, Vieira AM, Becker JD, Feijó JA. (2005). «Gametophyte interaction and sexual reproduction: how plants make a zygote» Int. J. Dev. Biol.. Vol. 49. pp. 615-632. doi: 10.1387/ijdb.052023lb.
- ↑ Deborah Charlesworth, Xavier Vekemans, Vincent Castric, Sylvain Glémin. 2005. Plant self-incompatibility systems: a molecular evolutionary perspective New Phytologist 168 (1) , 61–69 doi:10.1111/j.1469-8137.2005.01443.x.
- ↑ Charlesworth, D., X. Vekemans, V. Castric and S. Glemin (2005). «Plant self-incompatibility systems: a molecular evolutionary perspective.» New Phytologist. Vol. 168. n.º 12. pp. 61-69.
- ↑ Franklin, F. C. H., M. J. Lawrence, and V. E. Franklin-Tong (1995). «Cell and molecular biology of self-incompatibility in flowering plants» Int. Rev. Cytol.. Vol. 158. pp. 1-64.
- ↑ a b c d e f Franklin-Tong, V. E., and F. C. H. Franklin (2003). "The different mechanisms of gametophytic self-incompatibility." Philos. Trans. R. Soc. Lond. B. Biol. Sci. 358(1434): 1025–1032.
- ↑ McClure, B. A., V. Haring, , P. R. Ebert, M. A. Anderson, R. J. Simpson, F. Sakiyama, and A. E. Clarke (1989). "Style self-incompatibility gene products of Nicotiana alata are ribonucleases." Nature 342: 955–957.
- ↑ Sijacic, P., X. Wang, A. L. Skirpan, Y. Wang, P. E. Dowd, A. G. McCubbin, S. Huang, & T. Kao (2004). "Identification of the pollen determinant of S-RNase-mediated self-incompatibility." Nature 429: 302-305.
- ↑ Newbigin, E. (1996). The evolution of self-incompatibility: a molecular voyeur’s perspective. Sexual Plant Reproduction Volume 9, Number 6:357-361.[1]
- ↑ Matton, D.P., O Maes, G Laublin, Q Xike, C Bertrand, D Morse, and M Cappadocia (1996). «Hypervariable Domains of Self-Incompatibility RNases Mediate Allele-Specific Pollen Recognition.» Plant Cell. Vol. 9(10). pp. 1757–1766. PMCID 157019.
- ↑ Franklin-Tong, V. E., J. P. Ride, N. D. Read, A. J. Trewawas, & F. C. H. Franklin (1993). "The self-incompatibility response in Papaver rhoeas is mediated by cytosolic free calcium." Plant J. 4: 163–177.
- ↑ Franklin-Tong, V. E., G. Hackett, and P. K. Hepler (1997). "Ratioimaging of Ca21 in the self-incompatibility response in pollen tubes of Papaver rhoeas." Plant J. 12: 1375–1386.
- ↑ Franklin-Tong, V. E., T. L. Holdaway-Clarke, K. R. Straatman, J. G. Kunkel, and P. K. Hepler (2002). "Involvement of extracellular calcium influx in the self-incompatibility response of Papaver rhoeas." Plant J. 29: 333–345.
- ↑ Rudd, J. J., F. C. H. Franklin, J. M. Lord, & V. E. Franklin-Tong (1996). "Increased phosphorylation of a 26-kD pollen protein is induced by the self-incompatibility response in Papaver rhoeas." Plant Cell 8: 713–724.
- ↑ Geitmann, A., B. N. Snowman, , A. M. C. Emons, & V. E. Franklin-Tong (2000). "Alterations to the actin cytoskeleton of pollen tubes are induced by the self-incompatibility reaction in Papaver rhoeas." Plant Cell 12: 1239–1252.
- ↑ Snowman, B. N., D. R. Kovar, G. Shevchenko, V. E. Franklin-Tong, & C. J. Staiger (2002). "Signal-mediated depolymerization of actin in pollen during the self-incompatibility response." Plant Cell 14: 2613–2626.
- ↑ Jordan, N. D., F. C. H. Franklin, and V. E. Franklin-Tong (2000). "Evidence for DNA fragmentation triggered in the self-incompatibility response in pollen of Papaver rhoeas." Plant J. 23: 471–479.
- ↑ Thomas, S. G., and V. E. Franklin-Tong (2004). "Self-incompatibility triggers programmed cell death in Papaver pollen." Nature 429: 305-309.
- ↑ Goodwillie, C. (1997). "The genetic control of self-incompatibility in Linanthus parviflorus (Polemoniaceae)." Heredity 79: 424–432.
- ↑ a b Hiscock, S. J., and D. A. Tabah (2003). "The different mechanisms of sporophytic self-incompatibility." Philos. Trans. R. Soc. Lond. B. Biol. Sci. 358(1434): 1037-1045.
- ↑ Ockendon, D. J. (1974). "Distribution of self-incompatibility alleles and breeding structure of open-pollinated cultivars of Brussels sprouts." Heredity 32: 159–171.
- ↑ Schopfer, C. R., M. E. Nasrallah, and J. B. Nasrallah, (1999). "The male determinant of self-incompatibility in Brassica." Science 266: 1697–1700.
- ↑ Takayama, S., H. Shiba, M. Iwano, H. Shimosato, F.-S. Che, N. Kai, M. Watanabe, G. Suzuki, K. Hinata, and A. Isogai (2000). "The pollen determinant of self-incompatibility in Brassica campestris." Proc. Natl Acad. Sci USA 97: 1920–1925.
- ↑ Stein, J. C., B. Howlett, D. C. Boyes, M. E. Nasrallah, and J. B. Nasrallah (1991). "Molecular cloning of a putative receptor kinase gene encoded by the self-incompatibility locus of Brassica oleracea." Proc. Natl Acad. Sci. USA 88: 8816–8820.
- ↑ Nasrallah, J. B., and M. E. Nasrallah (1993). "Pollen–stigma signalling in the sporophytic self-incompatibility response." Pl. Cell 5: 1325–1335.
- ↑ Takasaki, T., K. Hatakeyama, G. Suzuki, M. Watanabe, A. Isogai, and K. Hinata (2000). "The S receptor kinase determines self-incompatibility in Brassica stigma." Nature 403: 913–916.
- ↑ Schopfer, C. R., and J. B. Nasrallah (2000). "Self-incompatibility. Prospects for a novel putative peptide-signaling molecule." Pl. Physiol. 124: 935–939.
- ↑ Takayama, S., H. Shimosato, H. Shiba, M. Funato, F.-E. Che, M. Watanabe, M. Iwano, and A. Isogai (2001). "Direct ligand–receptor complex interaction controls Brassica self-incompatibility." Nature 413: 534–538.
- ↑ Murase, K., H. Shiba, M. Iwano, F. S. Che, M. Watanabe, A. Isogai, and S. Takayama (2004). "A membrane-anchored protein kinase involved in Brassica self-incompatibility signaling." Science 303(5663): 1516-9.
- ↑ a b c d Ganders, F. R. (1979). "The biology of heterostyly." New Zealand Journal of Botany 17: 607-635.
- ↑ Darwin, C. R. 1862. On the two forms, or dimorphic condition, in the species of Primula, and on their remarkable sexual relations. Journal of the Proceedings of the Linnean Society of London (Botany) 6: 77-96. Reimpresión del trabajo original
- ↑ Ornduff, R., & S. G. Weller (1975). "Pattern diversity of incompatibility groups in Jepsonia heterandra (Saxifragaceae)." Evolution 29: 373-5.
- ↑ Ganders, F. R. (1976). "Pollen flow in distylous populations of Amsinckia (Boraginaceae)." Canadian Journal of Botany 54: 2530-5.
- ↑ Iain W. Manfield, Vassily K. Pavlov, Jinhong Li, Holly E. Cook, Florian Hummel, & Philip M. Gilmartin. 2005. Molecular characterization of DNA sequences from the Primula vulgaris S-locus. J. Exp. Bot. 56: 1177-1188.
- ↑ Spieth, P. T. (1971). "A necessary condition for equilibrium in systems exhibiting self-incompatible mating." Theoretical Population Biology 2: 404-18.
- ↑ a b Seavey, S. F., & K. S. Bawa (1986). "Late-acting self-incompatibility in angiosperms." Botanical Review 52: 195–218.
- ↑ Sage, T. L., R. I. Bertin, and E. G. Williams (1994). "Ovarian and other late-acting self-incompatibility systems." In E. G. Williams, R. B. Knox, and A. E. Clarke [eds.], Genetic control of self-incompatibility and reproductive development in flowering plants, 116–140. Kluwer Academic, Amsterdam.
- ↑ Sage, T. L., F. Strumas, W. W. Cole, and S. C. H. Barrett (1999). "Differential ovule development following self- and cross-pollination: the basis of self-sterility in Narcissus triandrus (Amaryllidaceae)." American Journal of Botany 86(6): 855–870.
- ↑ Sage, T. L., and E. G. Williams (1991). "Self-incompatibility in Asclepias." Plant Cell Incomp. Newsl. 23: 55–57.
- ↑ Sparrow, F. K., and N. L. Pearson (1948). "Pollen compatibility in Asclepias syriaca." J. Agric. Res. 77: 187–199.
- ↑ Lipow, S. R., and R. Wyatt (2000). "Single Gene Control of Postzygotic Self-Incompatibility in Poke Milkweed, Asclepias exaltata L." Genetics 154: 893–907.
- ↑ Bittencourt JR, N. S., P. E. Gibbs, and J. Semir (2003). "Histological study of post-pollination events in Spathodea campanulata Beauv. (Bignoniaceae), a species with late-acting self-incompatibility." Annals of Botany 91: 827-834.
- ↑ Glaettli, M. (2004). Mechanisms involved in the maintenance of inbreeding depression in gynodioecious Silene vulgaris (Caryophyllaceae): an experimental investigation. PhD dissertation, University of Lausanne.
- ↑ Weller, S. & R. Ornduff. 1977. Cryptic Self-Incompatibility in Amsinckia grandiflora. Evolution, Vol. 31, No. 1 pp. 47-51. resumen
- ↑ Eckert, E.G. & M. Allen. 1997. Cryptic self-incompatibility in tristylous Decodon verticillatus (Lythraceae). American Journal of Botany, Vol 84, 1391. resumen
- ↑ a b c Bateman, A. J. (1956). "Cryptic self-incompatibility in the wallflower: Cheiranthus cheiri L." Heredity 10: 257–261.
- ↑ Travers, S. E., and S. J. Mazer (2000). "The absence of cryptic self-incompatibility in Clarkia unguiculata (Onagraceae)." American Journal of Botany 87(2): 191–196.
- ↑ Chen KY, Tanksley SD. 2004. High-resolution mapping and functional analysis of se2.1: a major stigma exsertion quantitative trait locus associated with the evolution from allogamy to autogamy in the genus Lycopersicon. Genetics 168(3):1563-73.Resumen en inglés.
- ↑ Chen KY, Cong B, Wing R, Vrebalov J, Tanksley SD. Changes in regulation of a transcription factor lead to autogamy in cultivated tomatoes. Science 26;318(5850):643-645.Resumen en inglés.
Bibliografía
- Boavida LC, Vieira AM, Becker JD, Feijó JA. (2005). «Gametophyte interaction and sexual reproduction: how plants make a zygote» Int. J. Dev. Biol.. Vol. 49. pp. 615-632. doi: 10.1387/ijdb.052023lb.
- Frankel, R. & Galun, E. (1977). Pollination mechanisms, reproduction, and plant breeding. Monographs on Theoretical and Applied Genetics no. 2. Springer-Verlag. Berlin. ISBN 0387079343.
- Franklin-Tong, Vernonica E. (Ed.) (2008). Self-Incompatibility in Flowering Plants. Evolution, Diversity, and Mechanisms. Springer-Verlag. Berlin. ISBN 978-3-540-68485-5.
Enlaces externos
- University of Paisley, "Heterostyly in the Cowslip (Primula veris L.) "
- Cátedra de Morfología de Planta Vasculares, Facultad de ciencias Agrarias, Universidad Nacional de Nordeste, Corrientes, Argentina. "Alogamia"
Categorías: Reproducción vegetal | Polinización | Fisiología vegetal





Wikimedia foundation. 2010.