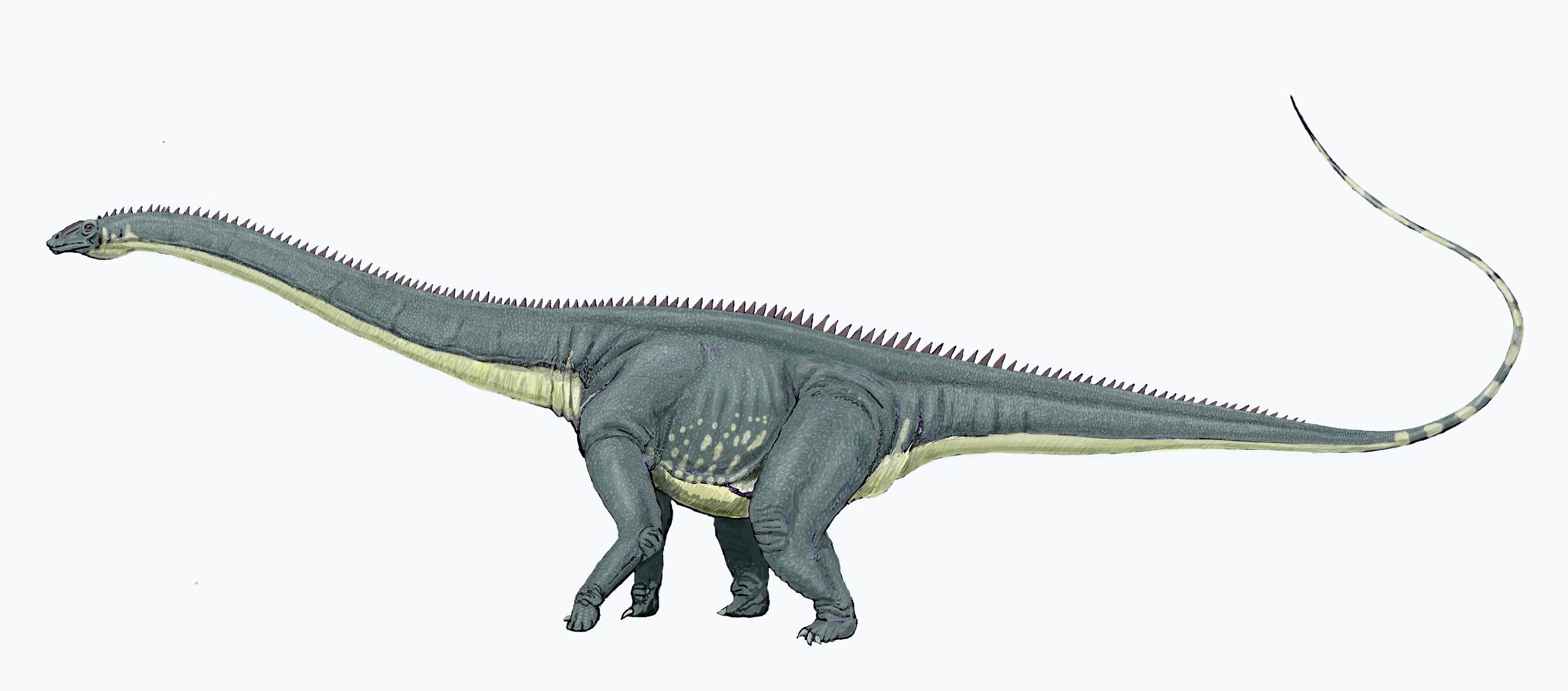
- Diplodocus
-
 Diplodocus
DiplodocusRango temporal: Jurásico superior 
Clasificación científica Reino: Animalia Filo: Chordata Clase: Sauropsida Superorden: Dinosauria Orden: Saurischia Suborden: Sauropodomorpha Infraorden: Sauropoda Superfamilia: Diplodocoidea Familia: Diplodocidae Subfamilia: Diplodocinae Género: Diplodocus
Marsh, 1878Especies Sinonimia - Seismosaurus Gillette, 1991
Diplodocus (gr. "doble viga")[1] es un género de dinosaurios saurópodos diplodócidos, que vivieron a finales del período Jurásico, hace aproximadamente 150 a 147 millones de años, en el Kimeridgiano y el Titoniano, en lo que hoy es Norteamérica. Los primeros fósiles fueron descubiertos en 1877 por S. W. Williston. El nombre genérico, acuñado por Othniel Charles Marsh en 1878, es la forma en neolatín derivada del griego διπλόος (diploos) "doble" y δοκός (dokos) "viga",[1] en referencia a la forma de los huesos a lo largo de la parte inferior de la cola. Estos huesos se pensaron exclusivos de los Diplodocus, sin embargo, han sido descubiertos en otros miembros de la familia de los diplodócidos y en saropódos no diplodócidos como Mamenchisaurus.
Vivió en lo que es hoy el oeste de Norteamérica durante el período Jurásico Superior. El Diplodocus es uno de los más comunes dinosaurios de la parte superior de la Formación Morrison, una secuencia de sedimentos marinos y aluvionales depositados alrededor de 150 a 147 millones de años atrás. La Formación Morrison muestra un ambiente dominado por gigantescos saurópodos como Camarasaurus, Barosaurus, Apatosaurus y Brachiosaurus además del Diplodocus.[2]
El Diplodocus está entre los dinosaurios más fácilmente identificables, con su forma clásica, cuello y cola larga y cuatro patas robustas. Durante muchos años, era el dinosaurio más grande conocido. Su gran tamaño pudo haber sido un impedimento a los depredadores Allosaurus y Ceratosaurus cuyos restos se han encontrado en los mismos estratos, lo que sugiere que coexistieron con el Diplodocus.
Contenido
Descripción
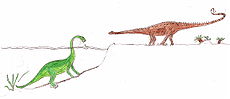
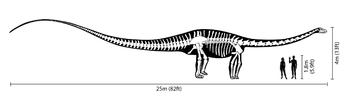 Comparación de tamaño con un ser humano.
Comparación de tamaño con un ser humano.
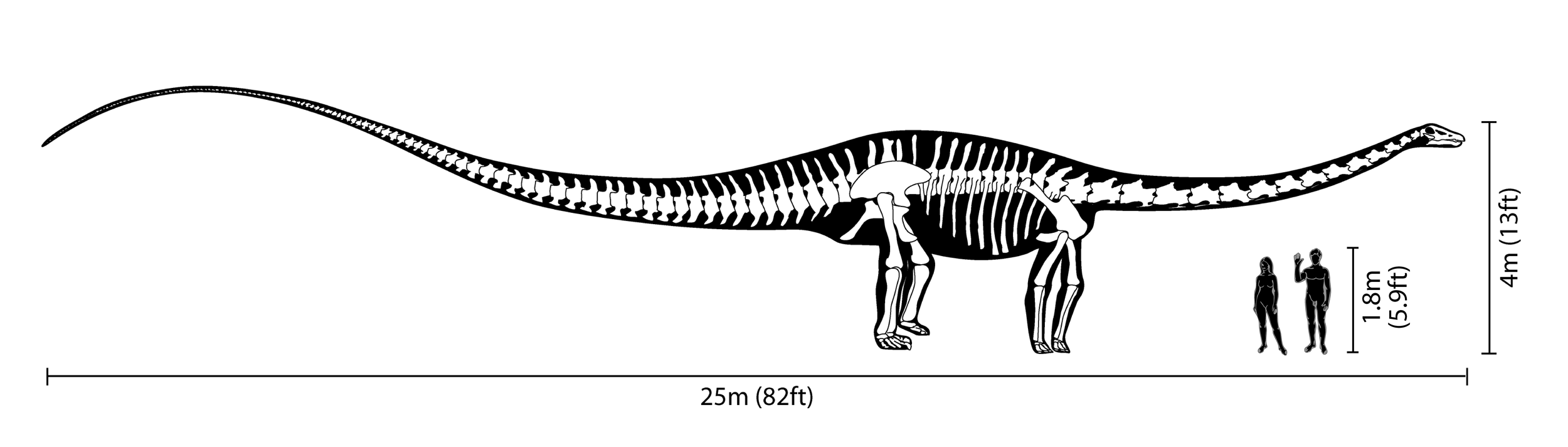
Los Diplodocus son uno de los dinosaurios mejor conocidos. Fueron animales cuadrúpedos muy grandes, de cuello largo y con una extensa cola en forma de látigo. Sus miembros delanteros eran ligeramente más cortos que sus miembros traseros, lo que configura una postura predominantemente horizontal. La mecánica de estos animales de cola y cuello largos, con cuatro patas macizas, ha sido comparada con la de un puente colgante.[3] De hecho, los Diplodocus es el dinosaurio más largo conocido del que se posee un esqueleto completo.[3] Aunque dinosaurios como el Supersaurus eran probablemente más largos, los restos fósiles hallados de estas otras especies son sólo parciales.[4] Los restos parciales de D. hallorum han aumentado la longitud estimada, aunque no tanto como se pensó previamente; cuando fue descrito en 1991, el descubridor, David Gillete, calculó que pudo haber medido hasta 54 metros de largo, haciéndole el dinosaurio más largo conocido, excepto los dudosos dinosaurios pobremente conocidos como Amphicoelias. Algunas estimaciones del peso se extendieron tanto como hasta 113 toneladas. Una revisión más reciente demuestra que las vértebras gigantes de la cola fueron colocadas realmente más adelante en la cola que lo que D. Gillete había calculado originalmente. El estudio demuestra que el esqueleto completo del Diplodocus en el Museo Carnegie de Historia Natural de Pittsburgh, Pensilvania, en el cual las estimaciones de Seismosaurus fueron basadas, incluía la decimotercera vértebra de la cola perteneciente a otro animal, lo cual redujo las estimaciones del tamaño en alrededor del 30%, quedando en alrededor de 37 metros de largo. Mientras que dinosaurios tales como los Supersaurus eran probablemente más largos, los fósiles encontrados hasta el momento que pertenecen a estos animales son solamente fragmentarios.[5]
El cráneo de los Diplodocus era muy pequeño en comparación con el tamaño del animal, los cuales llegaban a medir más de 37 metros de largo y 6,5 de alto al cuello.[6] Los Diplodocus poseían pequeños dientes con forma de clavija e inclinados hacia delante, los cuales estaban situados únicamente en la parte anterior de las mandíbulas.[7]
 Diplodocus carnegii, parte de la colección del Museo de La Plata.
Diplodocus carnegii, parte de la colección del Museo de La Plata.Su cuello estaba formado por al menos 15 vértebras, por lo que se cree que mantenían el cuello paralelo al suelo y que eran incapaces de elevarlo significativamente ( no más de 30 grados) por encima de su eje horizontal.[8] Por otro lado, las estimaciones modernas de su masa se sitúan en el rango de 10 a 16 toneladas: 10 t;[9] 11,5 t;[10] 12,7 t;[11] y 16 t.[12]
Los Diplodocus tenían una cola extremadamente larga, compuesta de más de 80 vértebras caudales,[13] cantidad que prácticamente duplica el número con el que contaban algunos saurópodos primitivos, como el Shunosaurus con 43, y que también supera a la que tenían macronarianos contemporáneos, como el Camarasaurus con 53. Existen especulaciones acerca de que los diplodocos podrían haber utilizado su cola como defensa[14] o para hacer ruido.[15] También podría haber servido como contrapeso para el cuello. La parte media de la cola tenía "dobles vigas" (huesos de forma extraña que le dan a los Diplodocus su nombre). Éstas pudieron haber servido para proporcionar apoyo para las vértebras, o quizás para evitar que los vasos sanguíneos se aplastasen en caso de que la pesada cola del animal chocase contra la tierra (o contra algún temerario depredador). Estas "dobles vigas" también aparecen en algunos dinosaurios emparentados con los Diplodocus.
Como otros saurópodos, la mano ("pie delantero") del Diplodocus estaba altamente modificada, con los huesos del dedo y de mano dispuestos en una columna vertical, con una sección de corte con forma de herradura. El Diplodocus carecía de garras a excepción del dedo 1 de la mano, que eran inusualmente grandes comparada a la de otros saurópodos, aplanada de lado a lado, y separados de los huesos de la mano. La función de esta garra inusualmente especializada es desconocida.[16]
Otros aspectos anatómicos
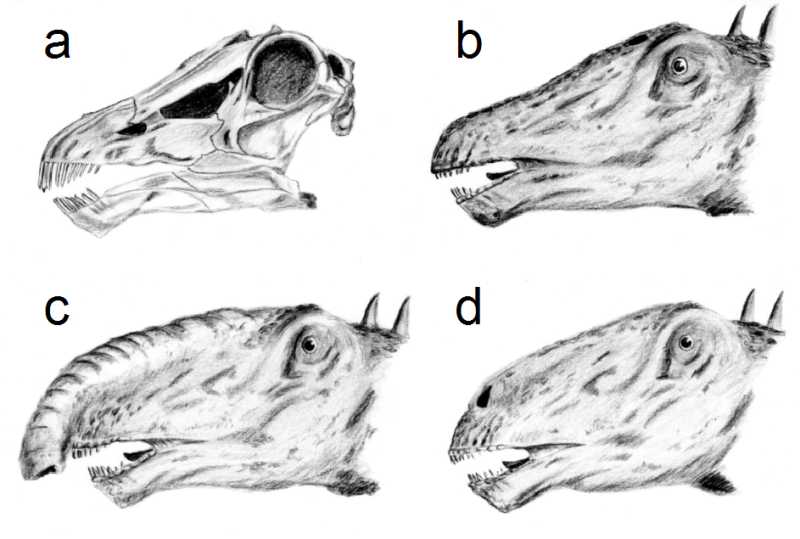
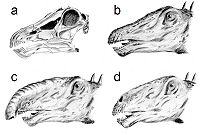 a) cráneo, b) clásica representación de la cabeza con las fosas nasales en lo alto de la misma, c) teoría de Bakker sobre una trompa, d) moderna representación con las fosas nasales abajo en el hocico y una posible cámara de resonancia.
a) cráneo, b) clásica representación de la cabeza con las fosas nasales en lo alto de la misma, c) teoría de Bakker sobre una trompa, d) moderna representación con las fosas nasales abajo en el hocico y una posible cámara de resonancia.La cabeza de los Diplodocus ha sido ampliamente representada con las fosas nasales en la parte superior de la misma, debido a la posición de las aberturas nasales en el ápice del cráneo. Ha habido especulaciones acerca de si tal configuración significa que los Diplodocus tuvieron una trompa.[17] Pero un estudio reciente[18] afirmó que no hay evidencias paleoneuroanatómicas que respalden la hipótesis de la trompa. Se ha señalado que el nervio facial de animales como el elefante es largo, ya que inerva una trompa. Las pruebas sugieren que el nervio facial es muy pequeño en los Diplodocus. Estudios de Lawrence Witmer (2001) indicaron que, mientras que las aberturas nasales estaban arriba en la cabeza, las actuales fosas nasales carnosas estaban situadas mucho más abajo, en el hocico.[19]
Recientes descubrimientos han demostrado que los Diplodocus y otros diplodócidos pudieron haber tenido unas estrechas y puntiagudas espinas de queratina alrededor del dorso, como las de una iguana.[20] [21] Esta característica, radicalmente diferente de la imagen que se tenía de estos animales, ha sido incorporada en recientes reconstrucciones. Se desconoce exactamente cuántos diplodócidos presentaron esta característica, y si además estuvo presente en otros saurópodos.
Historia
 Diplodocus en el Museo de Historia Natural del Instituto Smithsoniano.
Diplodocus en el Museo de Historia Natural del Instituto Smithsoniano.Varias especies de Diplodocus fueron descritas entre 1878 y 1924. El primer esqueleto fue hallado en el año 1878 en Como Bluff, Wyoming, por Benjamin Mudge y Samuel Wendell Williston. Fue el paleontólogo Othniel Charles Marsh quien le puso el nombre de Diplodocus longus ("doble viga larga") ese mismo año.[22] Desde entonces, restos de Diplodocus han sido encontrados en la Formación de Morrison (al oeste de Estados Unidos), en Colorado, en Utah, Montana y Wyoming. Los fósiles de estos dinosaurios son relativamente comunes, a excepción del cráneo. Aunque no es la especie prototípica, la más conocida es el D. carnegie, siendo la más famosa debido al gran número de exhibiciones que se han realizado de su esqueleto en museos de todo el mundo.
Los dos géneros de saurópodos de la Formación de Morrison, Diplodocus y barosaurio, comparten huesos muy similares en sus miembros. En el pasado, muchos huesos aislados fueron automáticamente atribuidos a los Diplodocus pero pudieron, de hecho, haber pertenecido al Barosaurus.[23]
Clasificación
 Según los estudios realizados, los Diplodocus se paraban en dos patas y estiraban su largo cuello para llegar a las hojas más verdes ubicadas en la copa de los árboles.
Según los estudios realizados, los Diplodocus se paraban en dos patas y estiraban su largo cuello para llegar a las hojas más verdes ubicadas en la copa de los árboles.Diplodocus son el género tipo de la familia Diplodocidae.[24] Los miembros de esta familia, aunque de gran tamaño, poseen una estructura más delgada en comparación con la de otros saurópodos, como los titanosaurianos y los braquiosáuridos. Todos están caracterizados por largos cuellos y colas y una postura horizontal, con los miembros delanteros más cortos que los traseros. Los diplodócidos dominaron a finales del Jurásico en Norteamérica y posiblemente África[13] y parecen haber sido reemplazados por los titanosáuridos durante el Cretácico.
Una subfamilia, Diplodocinae, fue creada para incluir a los Diplodocus y a sus parientes más cercanos, incluyendo al barosaurio. Apatosaurus, aunque contemporáneo, tiene un parentesco más lejano, pero sigue siendo considerado un diplodócido aunque no un diplodocino, ya que es un miembro de la subfamilia Apatosaurinae.[25] [26] Los géneros Dinheirosaurus y Tornieria han sido también identificados como parientes cercanos de los Diplodocus por algunos autores.[27] [28]
El grupo Diplodocoidea abarca a los diplodócidos, así como los dicreosáuridos, rebaquisáuridos, Suuwassea,[25] [26] amficoelias[28] y posiblemente haplocantosaurio.[29] Este clado es el grupo hermano de los camarasáuridos, braquiosáuridos y titanosaurianos; los Macronaria.[30] [29] El conjunto de estos taxones conforman Neosauropoda, el más grande, diverso y exitoso grupo de dinosaurios sauropodomorfos.
Especies válidas
 D. longus.
D. longus. D. carnegiei.
D. carnegiei. D. hallorum.
D. hallorum.D. longus
Es la especie tipo, la cual es conocida gracias a dos cráneos y una serie caudal procedentes de la Formación Morrison de Colorado y de Utah.[31]
D. carnegiei
Nombrado en honor a Andrew Carnegie, es la especie mejor conocida, principalmente debido a un esqueleto casi completo recogido por Jacob Wortman, del Museo Carnegie de Historia Natural en Pittsburgh, Pensilvania. La especie fue descrita y nombrada por John Bell Hatcher en 1901.
D. hallorum
Es más conocida como Seismosaurus hallorum, ya que en el 2004, una presentación en la conferencia anual de la Sociedad Geológica de América demostró que el Seismosaurus era en realidad una especie del Diplodocus.[32] Este estudio fue seguido por una publicación mucho más detallada en el 2006, la cual no solo renombró la especie D. hallorum, sino que también especuló con que dicha especie podría ser la misma que la del D. longus.[33] La posición de que D. hallorum se debería mirar como un espécimen de D. longus también fue tomada por los autores de una redescripción de Supersaurus, refutando una hipótesis anterior que el Seismosaurus y Supersaurus eran sinónimos.[34]
Especies dudosas
D. lacustris
Es una especie dudosa, nombrada por Marsh en 1884, a partir de restos de un pequeño animal proveniente de Morrison, Colorado.[24] En la actualidad se cree que estos huesos pertenecieron a un animal inmaduro, en vez de una especie separada.[35]
D. hayi
Es una especie conocida gracias a un esqueleto parcial descubierto por William H. Utterback en 1902 cerca de Sheridan, Wyoming. Fue descrita por W. J. Holland en 1924.[36] Es un ejemplar joven, su cráneo muestra particularidades que indican que probablemente sea un Apatosaurus.
Paleobiología
Postura
 Antes, algunos científicos creían que los Diplodocus caminaban con las piernas flexionadas, pero en la actualidad, esta teoría ya ha sido descartada.
Antes, algunos científicos creían que los Diplodocus caminaban con las piernas flexionadas, pero en la actualidad, esta teoría ya ha sido descartada.La representación de la postura de los Diplodocus ha cambiado considerablemente con los años. Por ejemplo, una obra clásica de 1910 de Oliver P. Hay representa a dos Diplodocus merodeando en la orilla de un río. Dichos animales cuentan con miembros abiertos hacia los lados, similares a los de los lagartos. Hay argumentó que los Diplodocus tenían un paso parecido al de un lagarto con patas sobresaliendo a ambos lados,[37] afirmación que fue apoyada por Gustav Tornier. Sin embargo, esta hipótesis fue rebatida por William J. Holland, quien demostró que un Diplodocus con esa postura, habría necesitado un foso para hacer pasar su vientre.[38]
Más adelante, los diplodócidos fueron retratados con frecuencia con sus cuellos elevados, lo que les permitiría comer de árboles altos, pero, recientemente, los científicos argumentaron que el corazón habría tenido problemas para mantener la presión arterial necesaria para oxigenar el cerebro. Además, estudios posteriores comprobaron que la estructura de las vértebras cervicales no pudieron permitir que el cuello se moviese tan arriba.[39] [40]
Al igual que con el Barosaurus, el cuello muy largo de los Diplodocus es fuente de mucha controversia entre los científicos. En 1992, un estudio de la Universidad de Columbia sobre la estructura del cuello de un diplodócido, indicó que los cuellos más largos debieron requerir de un corazón de 1.6 t. El estudio propuso que animales como éste debieron tener "corazones" auxiliares rudimentarios en sus cuellos, cuyo único propósito sería el de bombear la sangre al siguiente "corazón".[3]
Aunque el cuello largo ha sido tradicionalmente interpretado como una adaptación alimenticia, un reciente estudio[41] sugiere que los Diplodocus y sus parientes lo debieron utilizar, en primer lugar, como reclamo sexual. El posible uso alimenticio aparecería con posterioridad.
Galería de posturas
-

Esqueleto de un Diplodocus con la postura reciente, expuesto en el Museo de Historia Natural de Londres.
Alimentación
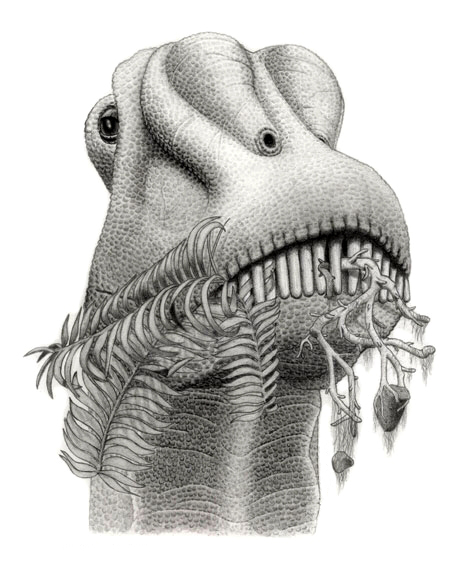
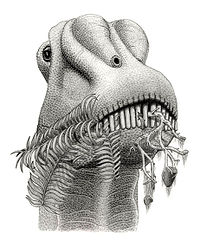 Un diplodócido comiendo helechos.
Un diplodócido comiendo helechos.Comparados con los dientes de otros saurópodos, los del Diplodocus eran muy extraños. Las coronas dentales eran largas, delgadas y elípticas en sección transversal, mientras que el ápice formaba un punto triangular embotado.[43] La faceta de uso más prominente está en el ápice; sin embargo, a diferencia de los demás patrones de uso observados dentro de los saurópodos, los del Diplodocus se ubican en el lado labial (mejilla) de los dientes superiores e inferiores.[43] Esto significa que los Diplodocus y otros diplodócidos tenían un mecanismo de alimentación radicalmente diferente al de otros saurópodos. El deshojamiento de ramas usando un lado del hocico es la forma de alimentación más probable del Diplodocus,[44] [45] [46] ya que explica los poco habituales patrones de desgaste de los dientes (a partir del contacto entre diente y comida). Al deshojar ramas usando un lado del hocico, una fila de dientes debió usarse para arrancar el follaje del tallo, mientras que la otra debió actuar como guía y estabilizador. Gracias a que contaba con una alongada región preorbital del cráneo (delante de los ojos), podría deshojar porciones más largas de tallos en una sola acción.[43] El movimiento hacia atrás de las mandíbulas inferiores pudo haber contribuido con dos significantes funciones en el comportamiento de alimentación: aumentar la apertura de la boca y permitir ajustes finos en las posiciones relativas de las filas de dientes para afinar el deshoje.[43]
Con un cuello lateral y dorsoventralmente flexible y la posibilidad de utilizar su cola para erguirse apoyándose en sus miembros traseros (habilidad "trípode"), los Diplodocus habría tenido la capacidad de alimentarse de hojas en muchos niveles (bajo, intermedio y alto), hasta aproximadamente 10 metros sobre el suelo, desde los pequeños ginkgos a las colosales araucarias.[47] El rango de movimiento del cuello también puede haberles permitido alimentarse de plantas acuáticas sumergidas. Esta hipótesis está apoyada por la relación de longitud entre los miembros delanteros y los traseros. Además, pudo haber usado sus dientes con forma de clavija para comer plantas marinas blandas y algas de litoral.[39]
Reproducción y crecimiento
Aunque no hay evidencia de hábitos de anidamiento de los Diplodocus, otros saurópodos como el titanosauriano Saltasaurus, han sido asociados con sitios de anidamiento.[48] [49] Los sitios de anidamiento de los titanosaurianos, indican que pudo haber depositado sus huevos comunalmente a lo largo de una gran área en muchos hoyos bajos, cada uno de ellos cubierto con vegetación. Esto ha generado la especulación de que los Diplodocus pudieron presentar un comportamiento similar. Otra teoría es la inclusión de las hembras en zonas de arboleda más frondosa, una vez fertilizadas, para la deposición de los huevos en un lugar "más seguro" que la desamparada planicie en la que habitualmente subsistían
Según varios estudios de histología de huesos, los Diplodocus, al igual que otros saurópodos, crecían a un ritmo muy rápido, alcanzando la madurez sexual en tan sólo una década y continuando con su crecimiento a lo largo de toda su vida.[50] [51] [52] Esta postura es radicalmente distinta de la que se atribuía a los saurópodos, los cuales se pensaba que crecían lentamente durante su vida, tomando décadas para alcanzar su madurez.
Paleoecología
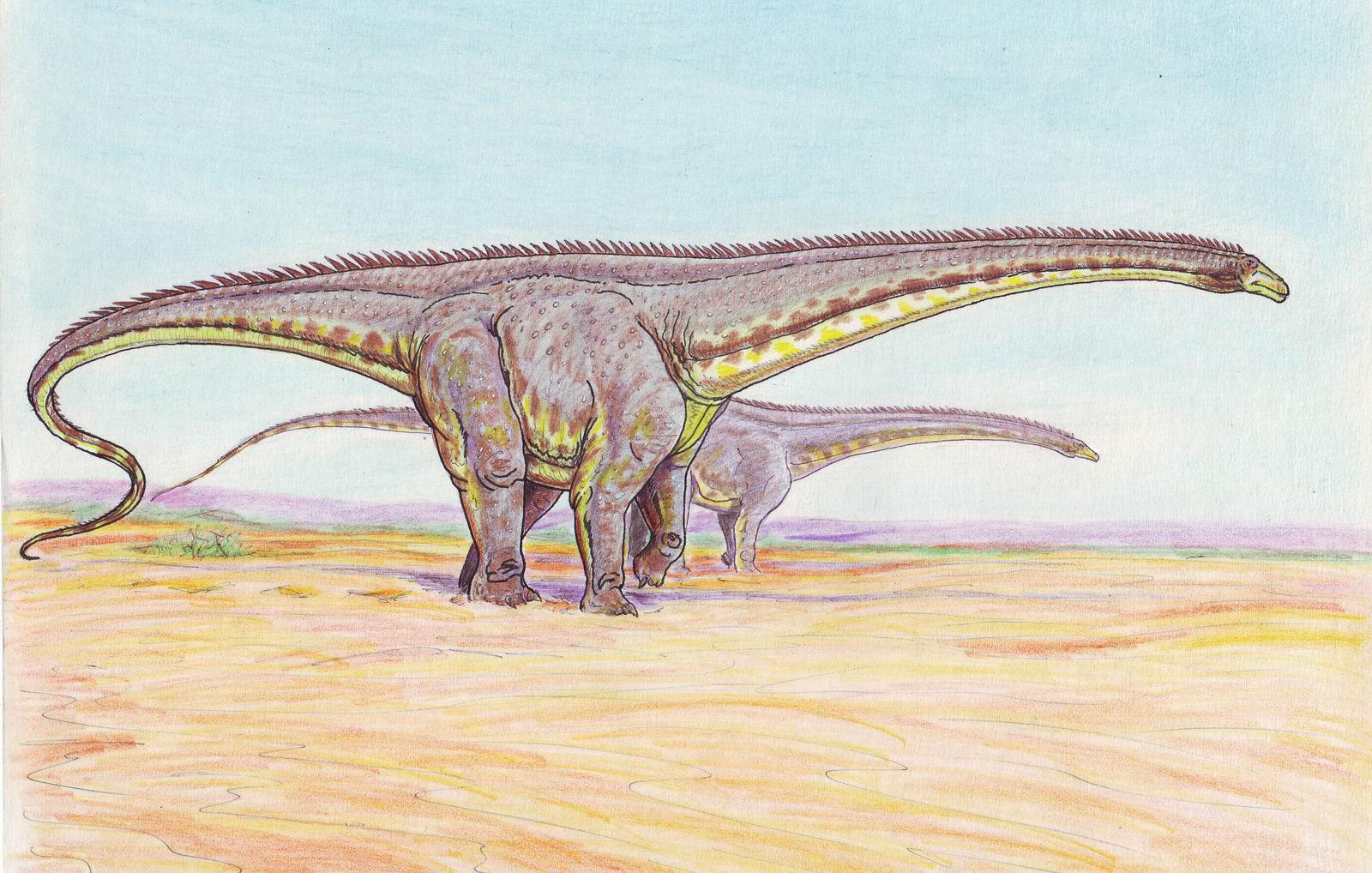
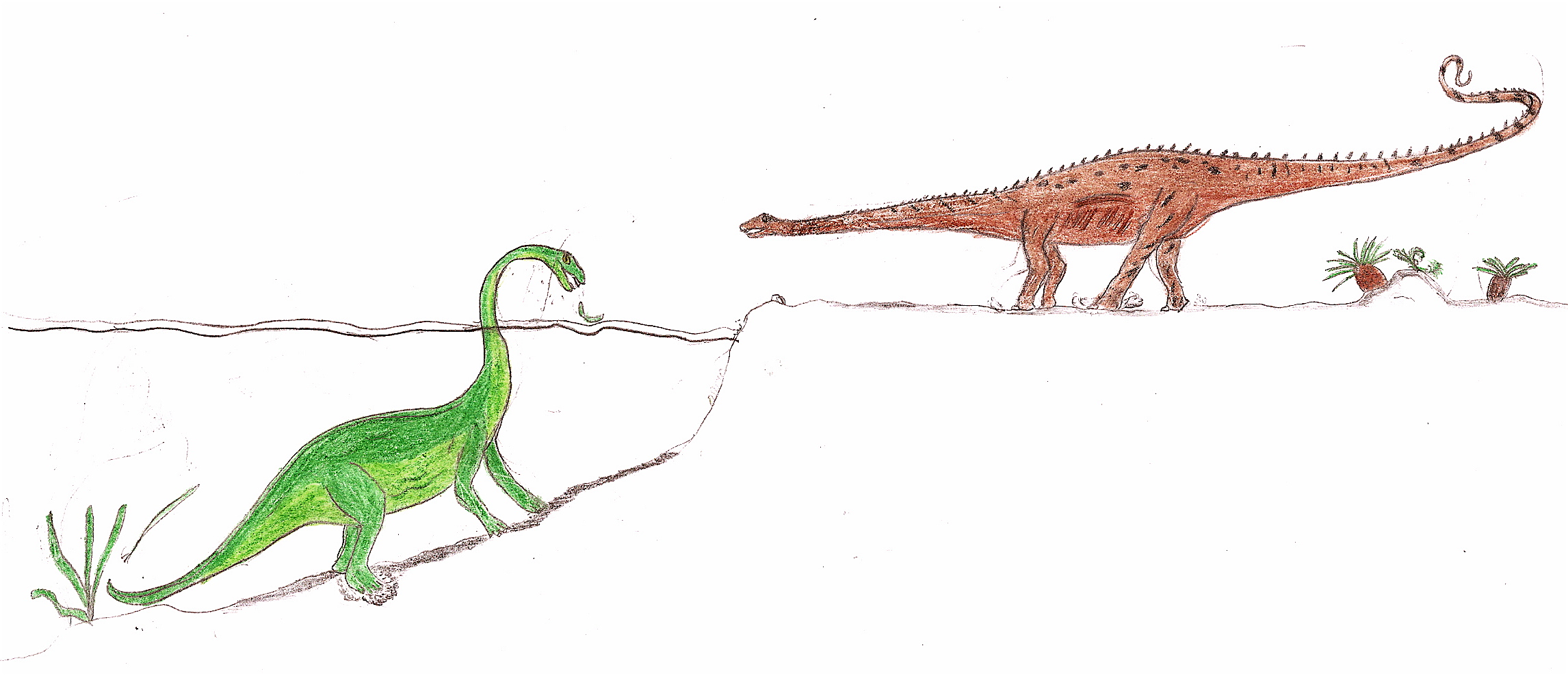
 Comparación de dos modos de vida de Diplodocus propuestos a lo largo de la Historia.
Comparación de dos modos de vida de Diplodocus propuestos a lo largo de la Historia.Debido a la posición de las aberturas nasales en el techo del cráneo, Marsh, y luego Hatcher,[53] supusieron que se trataba de un animal acuático. Este tipo de suposición también se realizó con otros saurópodos, como el braquiosaurio y el apatosaurio. La idea del entorno acuático fue posteriormente descartada, ya que la presión del agua en la pared pectoral de los Diplodocus habría sido demasiado grande como para que pudiesen respirar.[54] A partir de la década de 1970, existe un consenso generalizado acerca de que los saurópodos eran animales terrestres, que deambularon entre los árboles. Sin embargo, una teoría más reciente sugiere un hábitat ribereño para los Diplodocus.
El Diplodocus es el saurópodo más común en la zona extensa de rocas fósiles en el Suroeste Americano conocido la Formación de Morrison. La Formación Morrison es interpretada como un ambiente húmedo, y llanuras inundables.[55] La vegetación variaba de bosques a los lados de los ríos de coníferas, Helechos arborescentes y helechos, pasando a sabanas de helechos con algunos árboles dispersos.[56] La Formación Morrison es un área muy rica en fósiles, en el se encuentran clorofitos, hongos, musgos, equisetos, helechos, cycadales, ginkgos, y varias familias de coníferas. Otros fósiles incluyen bivalvos, caracoles, Actinopterigios, ranas, salamandras, tortugas, esfenodontos, lagartos, cocodrilomorfos terrestres y acuáticos, varias especies de pterosaurios, numerosas especies de dinosaurios, y mamíferos primitivos docodontes, multituberculados, symmetrodontes, y triconodontes. Algunos de los dinosaurios terópodos fueron Allosaurus Ceratosaurus, Ornitholestes, y Torvosaurus, los saurópodos junto con Diplodocus, Apatosaurus, Brachiosaurusy Camarasaurus y los ornitisquios Camptosaurus, Dryosaurus, y Stegosaurus son conocidos en Morrison.[57]
En la cultura popular
 Presentación de la primera réplica de Diplodocus carnegiei a los miembros del Museo de Historia Natural de Londres, 12 de mayo, 1905. Lord Avebury aparece pronunciando un discurso.
Presentación de la primera réplica de Diplodocus carnegiei a los miembros del Museo de Historia Natural de Londres, 12 de mayo, 1905. Lord Avebury aparece pronunciando un discurso.Diplodocus es un dinosaurio famoso y muy representado. Mucho de esto ha sido probablemente debido a su abundancia de restos esqueléticos y del estado anterior como el dinosaurio más largo. Sin embargo, la donación de muchos moldes esqueléticos montados por Andrew Carnegie a los potentados en todo el mundo al principio del siglo XX.[58] hizo mucho para que la gente del mundo se familiarice con este dinosaurio. Moldes de los esqueletos de Diplodocus todavía se exhiben en muchos museos por todo el mundo, incluyendo un inusual D. hayi en el Museo Houston de Ciencia Natural, y D. carnegiei en instituciones como elMuseo de Historia Natural de Londres, Museo Nacional de Ciencias Naturales de Madrid, España,[59] Museo Humboldt de Berlín, Alemania, el Museo Senckenberg de Fráncfort, Alemania, Museo Field de Historia Natural de Chicago, en el Museo de La Plata, en Argentina y, por supuesto, el original sigue siendo el centro de la exhibición en Museo Carnegie de Historia Natural en Pittsburgh. Un esqueleto montado de D. longus está en el Museo Nacional de Historia Natural de la Institución Smithsoniana en Washington D. C., mientras que un esqueleto montado de D. hallorum (antes Seismosaurus), que puede ser el mismo que D. longus, puede ser encontrado en Museo de Nuevo México de Historia Natural y Ciencia.
 Modelo de Diplodocus colocándose en sus piernas traseras en Bałtów, Polonia.
Modelo de Diplodocus colocándose en sus piernas traseras en Bałtów, Polonia.Diplodocus ha sido un tema frecuente en las películas del dinosaurio, documentales y ficticias. Fue mostrado en el segundo episodio de la serie de televisión premiada de la BBC; Walking with Dinosaurs. El episodio "Época del Titanes" sigue la vida de un Diplodocus hace 152 millones de años, enfrentandose a ataques por parte de Ornitholestes, Stegosaurus y Allosaurus. La película animada Fantasía ofrece muchos saurópodos en el cuadro de La consagración de la primavera, uno de ellos parece ser un Diplodocus. En literatura, James A. Michener libro Centennial tiene un capítulo dedicado a Diplodocus, narrando la vida y la muerte de un individuo. Diplodocus es una figura comúnmente vista entre los dinosaurios de juguete y maquetas. Ha tenido dos figuras separadas en Colección de Carnegie. En El mundo perdido: Parque Jurásico II, aparecen varios saurópodos que puede que sean Diplodocus y Mamenchisaurus.
Referencias
- ↑ a b Liddell & Scott (1980). Greek-English Lexicon, Abridged Edition. Oxford University Press, Oxford, UK. ISBN 0-19-910207-4.
- ↑ Christine C.E. & Peterson, F. (2004). "Reconstruction of the Upper Jurassic Morrison Formation extinct ecosystem—a synthesis". Sedimentary Geology 167, 309–355
- ↑ a b c Lambert D. (1993)The Ultimate Dinosaur Book ISBN 0-86438-417-3
- ↑ Wedel, M.J. and Cifelli, R.L. Sauroposeidon: Oklahoma’s Native Giant. 2005. Oklahoma Geology Notes 65:2.
- ↑ Wedel, M.J. and Cifelli, R.L. Sauroposeidon: Oklahoma's Native Giant. 2005. Oklahoma Geology Notes 65:2.
- ↑ Upchurch P, Barrett PM, Dodson P (2004). «Sauropoda». En Weishampel DB, Dodson P, Osmólska H. The Dinosauria (2nd Edition). University of California Press. pp. 316. ISBN 0-520-24209-2.
- ↑ Upchurch, P. & Barrett, P.M. (2000). Chapter 4: The evolution of sauropod feeding mechanism. IN: Evolution of Herbivory in Terrestrial Vertebrates ISBN 0-521-59449-9
- ↑ Stevens, K.A. & Parrish, M. (1999). "Neck Posture and Feeding Habits of Two Jurassic Sauropod Dinosaurs". Science 284, 798-800
- ↑ Dodson, P., Behrensmeyer, A.K., Bakker, R.T., and McIntosh, J.S. (1980). Taphonomy and paleoecology of the dinosaur beds of the Jurassic Morrison Formation. Paleobiology 6:208-232.
- ↑ Paul, G.S. (1994). Big sauropods - really, really big sauropods. The Dinosaur Report, The Dinosaur Society Fall:12-13.
- ↑ Foster, J.R. (2003). Paleoecological Analysis of the Vertebrate Fauna of the Morrison Formation (Upper Jurassic), Rocky Mountain Region, U.S.A. New Mexico Museum of Natural History and Science:Albuquerque, New Mexico. Bulletin 23.
- ↑ Coe, M.J., Dilcher, D.L., Farlow, J.O., Jarzen, D.M., and Russell, D.A. (1987). Dinosaurs and land plants. In: Friis, E.M., Chaloner, W.G., and Crane, P.R. (eds.). The Origins of Angiosperms and Their Biological Consequences. Cambridge University Press:New York, 225-258. ISBN 0-521-32357-6.
- ↑ a b Wilson JA (2005). «Overview of Sauropod Phylogeny and Evolution». En Rogers KA & Wilson JA(eds). The Sauropods:Evolution and Paleobiology. Indiana University Press. p. 15-49. ISBN 0-520-24623-3.
- ↑ Holland WJ (1915). «Heads and Tails: a few notes relating to the structure of sauropod dinosaurs.». Annals of the Carnegie Museum 9: pp. 273-278.
- ↑ Myhrvold NP and Currie PJ (1997). «Supersonic sauropods? Tail dynamics in the diplodocids». Paleobiology 23: pp. 393-409.
- ↑ Bonnan, M. F. (2003). "The evolution of manus shape in sauropod dinosaurs: implications for functional morphology, forelimb orientation, and phylogeny." Journal of Vertebrate Paleontology, 23: 595-613.
- ↑ Bakker, Robert T. (1986) The Dinosaur Heresies: New Theories Unlocking the Mystery of the Dinosaurs and their Extinction. New York: Morrow.
- ↑ Knoll, F., Galton, P.M., López-Antoñanzas, R. (2006). "Paleoneurological evidence against a proboscis in the sauropod dinosaur Diplodocus." Geobios, 39: 215-221
- ↑ Lawrence M. Witmer et al., "Nostril Position in Dinosaurs and other Vertebrates and its Significance for Nasal Function." Science 293, 850 (2001)
- ↑ Czerkas, S. A. (1993). "Discovery of dermal spines reveals a new look for sauropod dinosaurs." Geology 20, 1068-1070
- ↑ Czerkas, S. A. (1994). "The history and interpretation of sauropod skin impressions." In Aspects of Sauropod Paleobiology (M. G. Lockley, V. F. dos Santos, C. A. Meyer, and A. P. Hunt, Eds.), Gaia No. 10. (Lisbon, Portugal).
- ↑ Marsh OC. Principal characters of American Jurassic dinosaurs. Part I. American Journal of Science 3; 411-416 (1878).
- ↑ McIntosh (2005). «The Genus Barosaurus (Marsh)». En Carpenter, Kenneth and Tidswell, Virginia (ed.). Thunder Lizards: The Sauropodomorph Dinosaurs. Indiana University Press. pp. 38–77. ISBN 0-253-34542-1.
- ↑ a b Marsh, O.C. 1884. Principal characters of American Jurassic dinosaurs. Part VII. On the Diplodocidae, a new family of the Sauropoda. American Journal of Science 3: 160-168.
- ↑ a b Taylor, M.P. & Naish, D. 2005. The phylogenetic taxonomy of Diplodocoidea (Dinosauria: Sauropoda). PaleoBios 25(2): 1-7. (download here)
- ↑ a b Harris, J.D. 2006. The significance of Suuwassea emiliae (Dinosauria: Sauropoda) for flagellicaudatan intrarelationships and evolution. Journal of Systematic Palaeontology 4(2): 185–198.
- ↑ Bonaparte, J.F. & Mateus, O. 1999. A new diplodocid, Dinheirosaurus lourinhanensis gen. et sp. nov., from the Late Jurassic beds of Portugal. Revista del Museo Argentino de Ciencias Naturales. 5(2):13-29. (download here)
- ↑ a b Rauhut, O.W.M., Remes, K., Fechner, R., Cladera, G., & Puerta, P. 2005. Discovery of a short-necked sauropod dinosaur from the Late Jurassic period of Patagonia. Nature 435: 670-672.
- ↑ a b Wilson, J. A., 2002, "Sauropod dinosaur phylogeny: critique and cladistica analysis". Zoological Journal of the Linnean Society 136: 217-276.
- ↑ Upchurch P, Barrett PM, Dodson P (2004). «Sauropoda». En Weishampel DB, Osmólska H, Dodson P. The Dinosauria (2nd Edition). University of California Press. pp. 316. ISBN 0-520-24209-2.
- ↑ Upchurch P, Barrett PM, Dodson P (2004). «Sauropoda». En Weishampel DB, Osmólska H, Dodson P. The Dinosauria (2nd Edition). University of California Press. pp. 305. ISBN 0-520-24209-2.
- ↑ Reappraisal of Seismosaurus, A Late Jurassic Sauropod Dinosaur from New Mexico
- ↑ Lucas, S.G., Spielman, J.A., Rinehart, L.A., Heckert, A.B., Herne, M.C., Hunt, A.P., Foster, J.R., and Sullivan, R.M. (2006). «Taxonomic status of Seismosaurus hallorum, a Late Jurassic sauropod dinosaur from New Mexico». En Foster, J.R., and Lucas, S.G.. Paleontology and Geology of the Upper Morrison Formation. New Mexico Museum of Natural History and Science (bulletin 36). pp. 149-161. ISSN 1524-4156.
- ↑ Lovelace, David M.; Hartman, Scott A.; and Wahl, William R. (2007). «Morphology of a specimen of Supersaurus (Dinosauria, Sauropoda) from the Morrison Formation of Wyoming, and a re-evaluation of diplodocid phylogeny». Arquivos do Museu Nacional 65 (4): pp. 527–544.
- ↑ Upchurch, P., Barrett, P.M., and Dodson, P. (2004). "Sauropoda." In D. B. Weishampel, H. Osmólska, and P. Dodson (eds.), The Dinosauria (2nd edition). University of California Press, Berkeley 259-322.
- ↑ Holland WJ. The skull of Diplodocus. Memoirs of the Carnegie Museum IX; 379-403 (1924).
- ↑ Hay, Dr. Oliver P., "On the Habits and Pose of the Sauropod Dinosaurs, especially of Diplodocus." The American Naturalist, Vol. XLII, Oct. 1908
- ↑ Holland, Dr. W. J., "A Review of Some Recent Criticisms of the Restorations of Sauropod Dinosaurs Existing in the Museums of the United States, with Special Reference to that of Diplodocus carnegiei in the Carnegie Museum", The American Naturalist, 44:259–283. 1910.
- ↑ a b Stevens KA, Parrish JM (2005). «Neck Posture, Dentition and Feeding Strategies in Jurassic Sauropod Dinosaurs». En Carpenter, Kenneth and Tidswell, Virginia (ed.). Thunder Lizards: The Sauropodomorph Dinosaurs. Indiana University Press. pp. 212–232. ISBN 0-253-34542-1.
- ↑ Upchurch, P, et al. (2000). «"Neck Posture of Sauropod Dinosaurs"». Science 287, 547b (2000);DOI: 10.1126/science.287.5453.547b. Consultado el 28-11-2006.
- ↑ Senter,P. "Necks for Sex: Sexual Selection as an Explanation for Sauropod Neck Elongation". Journal of Zoology, 2006
- ↑ Hay, O. P., 1910, Proceedings of the
- ↑ a b c d Upchurch, P. & Barrett, P.M. (2000). Chapter 4: The evolution of sauropod feeding mechanism. IN: Evolution of Herbivory in Terrestrial Vertebrates ISBN 0-521-59449-9
- ↑ Norman, D.B. (1985). "The illustrated Encyclopedia of Dinosaurs". London: Salamander Books Ltd
- ↑ Dodson, P. (1990). Sauropod paleoecology. IN: "The Dinosauria" 1st Edition, (Eds. Weishampel, D.B., Dodson, P. & Osmólska, H.)
- ↑ Barrett, P.M. & Upchurch, P. (1994). Feeding mechanisms of Diplodocus. Gaia 10, 195-204
- ↑ Barrett, P.M. & Upchurch, P. (2005). Sauropodomorph Diversity through Time, Paleoecological and Macroevolutionary Implications. IN: "The Sauropods: Evolution and Paleobiology" (Eds. Curry, K. C.)
- ↑ Walking on Eggs: The Astonishing Discovery of Thousands of Dinosaur Eggs in the Badlands of Patagonia, by Luis Chiappe and Lowell Dingus. June 19, 2001, Scribner
- ↑ Grellet-Tinner, Chiappe, & Coria, "Eggs of titanosaurid sauropods from the Upper Cretaceous of Auca Mahuevo (Argentina)", Can. J. Earth Sci. 41(8): 949-960 (2004)
- ↑ Sander, P. M. (2000). "Long bone histology of the Tendaguru sauropods: Implications for growth and biology". Paleobiology 26, 466-488
- ↑ Sander, P. M., N. Klein, E. Buffetaut, G. Cuny, V. Suteethorn, and J. Le Loeuff (2004). "Adaptive radiation in sauropod dinosaurs: Bone histology indicates rapid evolution of giant body size through acceleration". Organisms, Diversity & Evolution 4, 165-173
- ↑ Sander, P. M., and N. Klein (2005). "Developmental plasticity in the life history of a prosauropod dinosaur". Science 310 1800-1802
- ↑ Hatcher JB. "Diplodocus (Marsh): Its osteology, taxonomy, and probable habits, with a restoration of the skeleton,". Memoirs of the Carnegie Museum, vol. 1 (1901), pp. 1-63
- ↑ Kermack, Kenneth A. (1951). «A note on the habits of sauropods». Annals and Magazine of Natural History 12 (4): pp. 830–832.
- ↑ Russell, Dale A. (1989). An Odyssey in Time: Dinosaurs of North America. Minocqua, Wisconsin: NorthWord Press. pp. 175–176. ISBN 1-55971-038-1.
- ↑ Carpenter, Kenneth (2006). «Biggest of the big: a critical re-evaluation of the mega-sauropod Amphicoelias fragillimus» (pdf). Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin, 36. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science. pp. 131–138. https://scientists.dmns.org/sites/kencarpenter/PDFs%20of%20publications/Amphicoelias.pdf.
- ↑ Chure, Daniel J.; Litwin, Ron; Hasiotis, Stephen T.; Evanoff, Emmett; and Carpenter, Kenneth (2006). «The fauna and flora of the Morrison Formation: 2006». En Foster, John R.; and Lucas, Spencer G. (eds.). Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin, 36. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science. pp. 233–248.
- ↑ Bakker, Robert T. (1986). The Dinosaur Heresies: New Theories Unlocking The Mystery of the Dinosaurs and Their Extinction. New York: William Morrow. pp. 203. ISBN 0140100555.
- ↑ Pérez García, A. y Sánchez Chillón, B. (2009). «Historia de Diplodocus carnegii del MNCN: primer esqueleto de dinosaurio montado en la Península Ibérica». Revista Española de Paleontología 24 (2): pp. 133-148.
Véase también
Enlaces externos
 Portal:Dinosaurios. Contenido relacionado con Dinosaurios.
Portal:Dinosaurios. Contenido relacionado con Dinosaurios. Wikimedia Commons alberga contenido multimedia sobre DiplodocusCommons.
Wikimedia Commons alberga contenido multimedia sobre DiplodocusCommons. Wikiespecies tiene un artículo sobre Diplodocus. Wikispecies
Wikiespecies tiene un artículo sobre Diplodocus. Wikispecies- Diplodocus in the Dino Directory (en inglés)
- Museo Nacional de Ciencias Naturales, Madrid
- Diplodocus Marsh, by J.B. Hatcher 1901 - Su Osteología, Taxonomía, y Probables Hábitos, con una Reconstrucción del Esqueleto. Memorias del Museo Carnegie, Volume 1, Number 1, 1901. Texto completo, de lectura libre. (en inglés)
- Museo Carnegie de Historia Natural - Historia (en inglés)
- Reconstrucciones esqueléticas de diplodócidos incluyendo D. carnegii, D. longus, y D. hallorum, de Scott Hartman's Skeletal Drawing website (en inglés).
- Diplodoco en duiops
- Diplodoco en Dinodata (en inglés)
Categorías:- Fósiles
- Diplodocoidea
- Dinosaurios del Jurásico
- Dinosaurios norteamericanos










Wikimedia foundation. 2010.