- Majungasaurus
-
 Majungasaurus
MajungasaurusRango temporal: Cretácico superior 
Clasificación científica Reino: Animalia Filo: Chordata Clase: Sauropsida Superorden: Dinosauria Orden: Saurischia Suborden: Theropoda Infraorden: Ceratosauria Familia: Abelisauridae Género: Majungasaurus
Lavocat, 1955Especies - M. crenatissimus (tipo)
(Depéret, 1896)
Sinonimia Majungatholus
Sues and Taquet, 1979Majungasaurus ("Mahajanga" + gr. "lagarto" = "Lagarto de Mahajanga") es un género de dinosaurios terópodos abelisáuridos que existieron a finales del período Cretácico, hace aproximadamente 80 millones de años, en el Campaniano, en lo que hoy es Madagascar. Solo una especie, M. crenatissimus, ha sido asignada. A este género solía llamárselo Majungatholus, nombre que ahora se considera sinónimo de Majungasaurus.
Al igual que otros abelisáuridos, el Majungasaurus era un depredador bípedo provisto de un hocico corto. Aunque los brazos no disponen de mucha evidencia, se reconoce que estos eran muy pequeños, mientras las patas eran largas y robustas. Por otro lado, se diferencia de los demás abelisáuridos al contar con un cráneo más ancho, dotado de una textura muy áspera y endurecida en el tope del hocico, más un cuerno redondeado arriba del cráneo, el cual fue originalmente confundido con el domo de un paquicefalosauriano. También poseía más dientes en ambas mandíbulas que la mayoría de abelisáuridos.
Tras ser conocido por numerosos cráneos y esqueletos bien preservados, el Majungasaurus ha llegado a ser uno de los dinosaurios terópodos más estudiados del Hemisferio Sur. Aparentemente estaba más relacionado con los abelisáuridos de la India que con los de Sudamérica o África continental, lo cual tiene importantes repercusiones biogeográficas. El Majungasaurus era el superpredador de su ecosistema, cazando principalmente saurópodos como el Rapetosaurus, y siendo también el único dinosaurio del cual hay evidencia directa de canibalismo (revisar coelophysis).
Contenido
Descripción
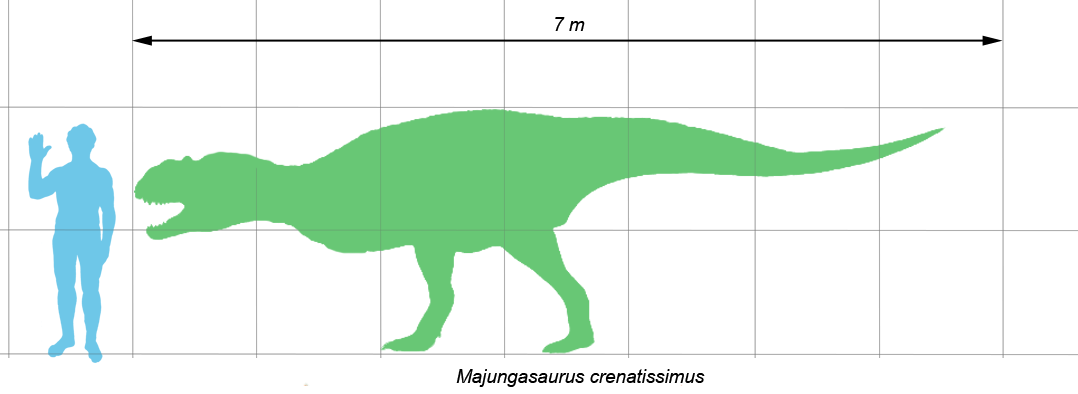
El Majungasaurus era el típico terópodo mediano que alcanzaba entre 6 y 7 metros de longitud, incluyendo su cola.[1] Restos incompletos de grandes individuos indican que algunos adultos llegaban a medir una longitud de más de 8 metros.[2] Los científicos estiman que un Majungasaurus adulto promedio pesaba más de 1100 kilogramos, sin embargo los más grandes individuos habrían pesado más.[2]
El cráneo del Majungasaurus ha sido excepcional y detalladamente estudiado a diferencia del de muchos terópodos. Como en los cráneos de otros abelisáuridos, su longitud era pequeña en proporción a su altura, sin embargo no tan pequeña como en el Carnotaurus. Cráneos de grandes individuos tenían una longitud que oscilaba entre los 60 y 70 cm. Un alto premaxilar, el cual embotaba la punta del hocico, era también común en su familia. Sin embargo, el cráneo del Majungasaurus era apreciablemente más ancho que el de otros abelisáuridos.
 Majungasaurus crenatissimus comparaddo en tamaño con un humano.
Majungasaurus crenatissimus comparaddo en tamaño con un humano.
Todos los abelisáuridos tenían una textura rugosa y esculpida en las caras exteriores de los huesos del cráneo, y Majungasaurus no era una excepción. Esto fue llevada a un extremo en los huesos nasales de Majungasaurus, que eran extremadamente gruesos estando fundidos entre sí, con un canto central bajo corriendo a lo largo de la mitad del hueso más cercano a las narinas. Un distintivo cuerno con forma de bóveda se asomaba de los huesos frontales también fundidos en la parte superior del cráneo. En vida, estas estructuras habrían sido cubiertas con una cierta clase de integumento, posiblemente de queratina. Una tomografía computada del cráneo muestra que la estructura nasal y el cuerno frontal tuvieron las cavidades huecas similares a senos, quizás para reducir el peso.[2] Los dientes eran típicos de abelisáuridos en tener coronas cortas, sin embargo Majungasaurus tenía 17 en cada maxilar de la mandíbula superior y en el dentario de la mandíbula inferior más que cualquier otro abelisáurido a excepción de Rugops.[3]
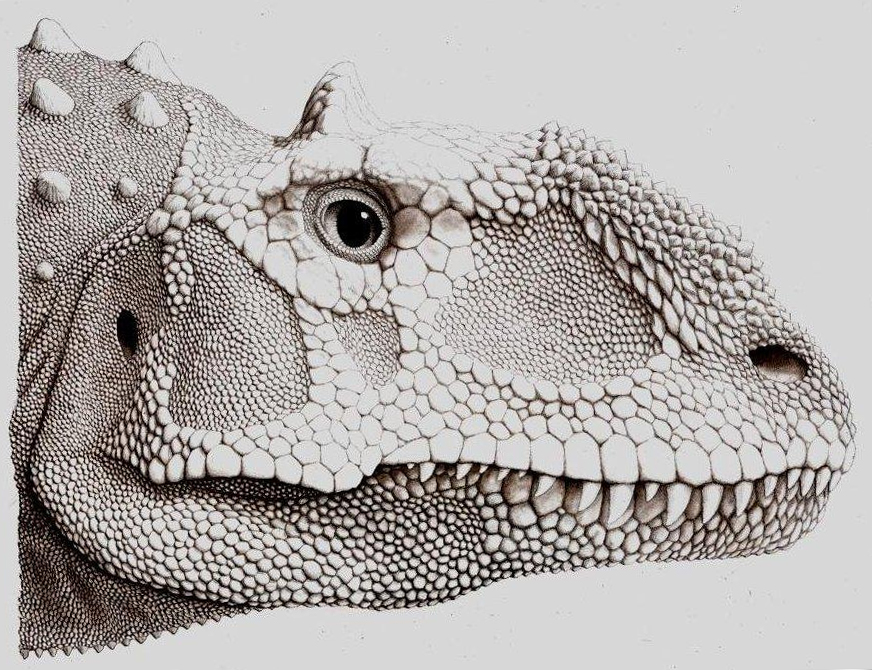
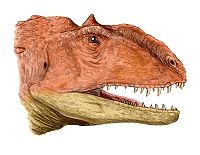 reconstrucción de la cabeza del Majungasaurus.
reconstrucción de la cabeza del Majungasaurus.El esqueleto postcraneal de Majungasaurus es muy parecido al de Carnotaurus y Aucasaurus, los otros dos únicos géneros de abelisáuridos de los que se conoce el esqueleto completo. Majungasaurus era bípedo, con una larga cola para balancear el torso y la cabeza, poniendo el centro de gravedad sobre las caderas. Aunque las vértebras cervicales (del cuello) tuvieran cavidades y excavaciones numerosas (pleurocoelas) para reducir su peso, eran robustas, con los sitios para la inserciones musculares bien marcados y las costillas que se entrecruzaban para dar fuerza. Tendones osificados unidos a las costillas cervicales, dándoles un aspecto bifurcado, según lo visto en Carnotaurus. Todas estas características dieron lugar a un cuello muy fuerte y muscular. Se diferenciaban en que las costillas cervicales de Majungasaurus tenían depresiones largas a lo largo de los lados para la reducción de peso.[4] El humero es el único hueso del brazo descrito, era corto y curvado, asemejándose a los de Aucasaurus y de Carnotaurus . Esto puede indicar que Majungasaurus tenía los miembros superiores muy cortos similares a los de estos dos géneros, con cuatro dedos. Como otros abelisáuridos, los miembros inferiores eran gruesos y cortos comparados a la longitud de cuerpo. La tibia de Majungasaurus era incluso más gruesa que la de Carnotaurus, con una cresta prominente en la rodilla. El astrágalo y el calcáneo estaban fundidos juntos, y los pies tenían tres dígitos funcionales, con un primer dígito más pequeño que no entraba en contacto con el suelo.[5]
Historia
El paleontólogo frances Charles Depéret describió los primeros restos de terópodo provenientes del noroeste de Madagascar en 1896. Estos incluían dos dientes, una garra, y algunas vértebras descubiertas a lo largo del Río Betsiboka por un oficial del ejército frances y depositadosen la colección del que es hoy la Université Claude Bernard Lyon 1. Depéret refirió estos fósiles al género Megalosaurus, que en ese tiempo era un taxón cajón de sastre conteniendo un número de terópodos pocos conocidos, creando una nueva especie Megalosaurus crenatissimus.[6] Este nombre deriva del latín crenatus ("muescas") y el sufijo -issimus ("muchas"), en referencias a las numerosas endentaduras en los bordes anterir y posterior del diente.[1] Depéret posteriormente reasigno la especie al género norteaméricano Dryptosaurus, otro taxón pobremente conocido.[7]
 El holotipo de Majungatholus, un hueso frontal fundido, alguna vez considerado parte de un paquicefalosáurido.
El holotipo de Majungatholus, un hueso frontal fundido, alguna vez considerado parte de un paquicefalosáurido.Numerosos fragmentos provenientes de la Provincia de Mahajanga en el noroeste de Madagascar donde fueron recobrados por coleccionistas franceses por más de 100 años, muchos de ellos depositados en el Muséum National d'Histoire Naturelle en París.[1] En 1955, René Lavocat describió un dentario con dientes de la Formación Maevarano en la misma región de los descubrimiento originales. Los dientes coincidian con los descritos por Depéret, pero el hueso fuertemente curvado, era diferente a los de Megalosaurus y Dryptosaurus. Lavocat renombro al género Majungasaurus, usando un antiguo nombre de Mahajanga y la conocida palabra griega σαυρος/sauros , haciendo del hueso de la mandíbula (MNHN.MAJ 1) el espécimen tipo.[8] Hans-Dieter Sues y Philippe Taquet describieron un domo craneal (MNHN. MAJ 4) como un paquicefalosauriano (Majungatholus atopus) en 1979. Este fue el primer reporte de un paquicefalosaurido en el Hemisferio Sur.[9]
 Mandíbula deMajungasaurus crenatissimus, Muséum national d'histoire naturelle, París.
Mandíbula deMajungasaurus crenatissimus, Muséum national d'histoire naturelle, París.En 1993, científicos de la State University of New York at Stony Brook y de la Université d’Antananarivo comenzaroncon el Proyecto del Lecho Mahajanga, una serie expediciones para examinas los fósiles y la geología de lso sedimentos del Cretácico Superior cerca de la Villa de Berivotra, en la Provincia de Mahajanga.[1] La primer expedición se topo con centenares de dientes del terópodos idéntico a los de Majungasaurus, algunos incertados a un premaxilar aislado que fue descrito en 1996.[10] Las siete expediciones siguientes encontraron cerca de de diez mil fósiles, muchos pertenecientes a especies nuevas para la ciencia. Al Proyecto del Lecho de Mahajanga se le acredita el crédito para quintuplicar la diversidad sabida de taxones fósiles en la región.[1] Fieldwork en 1996 encontró una cráneo exquisitamente conservado (FMNH PR 2100). En el techo del cráneo se encontraba un domo idéntico al descrito por Sues yTaquet para su Majungatholus atopus. Majungatholus fue redescrito como un abelisáurido, no como un paquicefalosáurido en 1998. Sin embargo aun cuando el nombre Majungasaurus crenatissimus era más viejo que Majungatholus atopus, los autores juzgaron que el dentario tipo de Majungasaurus era muy fragmentariopara aseverar que pertenecia la mismo género del cráneo.[11] El trabajo adicional en el terreno durante la década siguiente dio con una serie de cráneos menos completos, así como docenas de esqueletos parciales de individuos que se extendían de jóvenes a los adultos. Miembros del proyecto, también recogieron centenares de dientes deMajungasaurus. Tomándolos juntos, estos restos representan casi la totalidad del esqueleto, sin embargo, la mayor parte de los brazos, la mayoría de la pelvis y el extremo de la cola siguen sin conocerse.[1] Elte trabajo de campo culmino en 2007 con una monografía consistente en siete trabajos cientificos que abarcan todos los aspectos de la biología del animal publicado en las Memorias de la Society of Vertebrate Paleontology. Los trabajos están escritos en inglés con un resumen en magalache.[12] En este volumen, el dentario descrito por Lavocat es re evaluado y considerado diagnostico para la especie. Por eso, el nombre Majungatholus es remplazado por el anterior Majungasaurus.[1] Aunque la monografía es muy completa, los editores apuntan que solo se describió el material hallado entre 1993 y 2001. Un significativo número de especímenes en muy buen estado, fueron excavados entre 2003 y 2005 y aguardan preparación, descripción y publicación en futuros trabajos.[1]
Clasificación
Majungasaurus es clasificado como miembro del clado de Theropoda Abelisauridae, que es considerado una familia en taxonomía linneana. Junto con la familia Noasauridae, los abelisáuridos son incluidos en la Superfamilia Abelisauroidea, que a su vez es una subdivisión del infraorden Ceratosauria.[1] [13] Abelisáuridos es conocida por sus cráneos altos, hocicos cortos con una gran ornamentación de los huesos de la cara, un rasgo convergente con los carcarodontosáuridos, miembros anteriores muy reducidos, rasgo convergente con los tiranosáuridos, y proporciones cortas y gruesas de los miembros inferiores, entre otras características.[14]
Como muchos otros grupos de dinosaurios, la sistemática dentro de la familia Abelisauridae es confusa. Varios análisis cladisticos han indicado que Majungasaurus comparte una relación estrecha con Carnotaurus fde Sudamérica,[13] [14] mientras que otros no encuentran lugar dentro de la filogenia donde ubicarlo.[15] El análisis más reciente, usando la más completa información reciente, se ha ubicado a Majungasaurus en un clado junto a Rajasaurus y Indosaurus de la India, pero excluyendo géneros como Carnotaurus, Ilokelesia, Ekrixinatosaurus, Aucasaurus y Abelisaurus de Sudamérica, así como Rugops del continente de África. Esto deja abierto la posibilidad de dos clados separados de abelisáuridos en el oeste y el este Gondwana.[1] La descripción detallada de abelisáuridos conocidos como Aucasaurus así como los descubrimientos y los análisis futuros puede ayudar a resolver el cuadro filogenético.
Paleobiología
Ornamentación craneana
Majungasaurus quizás es el más distintivo de ornamentación de craneal, incluyendo los gruesos nasales fusionados y el cuerno frontal. Otros ceratosaurianos, incluyendo al Carnotaurus, Rajasaurus y Ceratosaurus tenían crestas en su cabeza. Estas estructuras se han visto generalmente como usadas en la competición intraespecífica, aunque su función exacta dentro de ese contexto sea desconocida. La cavidad hueca dentro del cuerno frontal de Majungasaurus habría debilitado la estructura y había imposibilitado probablemente su uso en combate físico directo, aunque el cuerno pudo haber servido para el propósito de exhibición .[14] Mientras que hay variación en la ornamentación entre individuos de Majungasaurus, no hay evidencias de dimorfismo sexual.[2]
Comportamiento alimenticio
Científicos han sugerido que la forma única del cráneo de Majungasaurus y otros abelisáuridos indican comportamientos de cacería distintos que los otros terópodos. Considerando que la mayoría de los terópodos se caracterizaban cráneos largos, bajos y estrechos, los cráneos de abelisáuridos eran más altos y más anchos, y a menudo más cortos en longitud también.[2] The los cráneos estrechos de otros terópodos estaban bien equipados soportar la tensión vertical de una mordedura larga, pero son buenos en soportar torsión.[16] En comparación con los modernos mamíferos predadores, la mayoría de los terópodos pudieron haber utilizado una estrategia similar en cierto modo a la de los modernos cánidos con un hocico largo y estrecho, con la daban de muchas mordeduras que debilitan el animal presa.[17]
 Restauración del la cabeza del Majungasaurus crenatissimus
Restauración del la cabeza del Majungasaurus crenatissimusAbelisáuridos, especialmente Majungasaurus, se puede en lugar de otro haber adaptado para una estrategia la alimentación más similar a moderno felinos, con un corto y ancho hocico, con el que muerden una sola vez hasta someter a la presa. Majungasaurus tenía un hocico incluso más amplio que otros abelisáuridos, que junto a otros aspectos de su anatomía apoyan la teoría de morder y sostener. El cuello es muy fuerte, con robustas vértebras, con las costillas cervicales entrelazadas con tendones osificados, así como profundos sitios de inserción para los fuertes músculos en el cuelo y la parte posterior del cráneo. Estos músculos habrían podido mantener la mordida en forma constante a pesar de las luchas de su presa. Los cráneos de los abelisáuridos también fueron consolidados en muchas áreas por el hueso mineralizado fuera de piel, creando la textura áspera característica de los huesos. Esto es particularmente cierto en Majungasaurus, donde los huesos nasales fueron fusionados y engrosados para aumentar la fortaleza del cráneo. Por otra parte, la quijada inferior de Majungasaurus soportaba una gran fenestra (abertura) en cada lado, según lo visto en otro ceratosaurianos, así como una articulación sinovial entre ciertos huesos que permitieron un alto nivel de flexibilidad en la mandíbula inferior, aunque no tanto como en las serpientes. Esto pudo haber sido una adaptación para prevenir la fractura de la mandíbula inferior al sostener una presa durante la lucha. Los dientes delanteros de la quijada superior eran más robustos que el resto, proporcionando un punto de anclaje para la mordedura, mientras que la altura baja de la corona de los dientes de Majungasaurus habría prevenido que se rompan durante los forcejeos. Finalmente, a diferencia de los dientes de Allosaurus y la mayoría de otros terópodos, que eran curvados tanto en el frente como detrás, abelisáuridos como Majungasaurus tenía dientes curvados en el borde delantero pero más rectos en el borde trasero. Esta estructura pudo haber servido evitar rebanar en lugar de sostener con los dientes al morder.[2]
 Majungasaurus crenatissimus
Majungasaurus crenatissimusMajungasaurus fue el mayor depredador de su ambiente, donde los únicos grandes herbívoros eran saurópodos como Rapetosaurus. Los expertos han sugerido que Majungasaurus, y quizás otros abelisáuridos, se hayan especializado en la caza de saurópodos. Las adaptaciones para consolidar la cabeza y el cuello para una mordedura el tipo de morder y sostener pudieron haber sido muy útiles contra los saurópodos, que habrían sido animales muy fuertes. Esta hipótesis también es apoyada por la forma de las patas de Majungasaurus, que eran cortas y robustas, en oposición de las largas y delgadas de otros terópodos. Debido a esto Majungasaurus no debió moverse tan rápido como otros terópodos de tamaño similar, pero esto no presentaba ningún problema ya que los saurópodos eran más lentos que él. Los huesos robustos de las patas sugieren las piernas muy fuertes, y su longitud más corta habría bajado el centro de gravedad del animal. Así Majungasaurus habría sacrificado velocidad por poder.[2] Marcas de dientes de Majungasaurus en huesos de Rapetosaurusconfirman que estos fueron partes de la dieta del dinosaurio, independientemente de si los mató realmente.[18]
Canibalismo
Aunque los saurópodos pudieron haber sido la presa de la opción para Majungasaurus, recientes descubrimientos en Madagascar indican otro componente sorpresa en la dieta: otros Majungasaurus. Numerosos huesos de Majungasaurus se han descubierto con marcas de dientes idénticas a ésas encontradas en los huesos de saurópodos de los mismos lugares. Estas marcas tienen el mismo espaciamiento que los dientes que en las mandíbulas de Majungasaurus, y son del mismo tamaño que los dientes de Majungasaurus, y contienen muescas más pequeñas constantes con las endentaduras en esos dientes. Como Majungasaurus es el único gran terópodo conocido del área, la más simple explicación es que comían individuos de su misma especie.[18] Sugerencias como esta en el Triásico Coelophysis han sido recientemente descartadas, dejando Majungasaurus como el único terópodo no aviano con sospechas de canibalismo,[19] aunque hay cierta evidencia de que el canibalismo pudo haber ocurrido en otras especie también.[20]
Es desconocido si Majungasaurus cazara activamente su propia clase o solamente comiera las carcasa muertas.[18] Sin embargo, algunos investigadores han observado eso moderno Dragón de Komodo se matan entre si al competir por el acceso a las reses muertas. Los dragones entonces procederá a comer las piezas aprovechables de los restos de sus rivales, que pueden sugerir comportamiento similar en Majungasaurus y otros terópodos.[20]
Sistema respiratorio
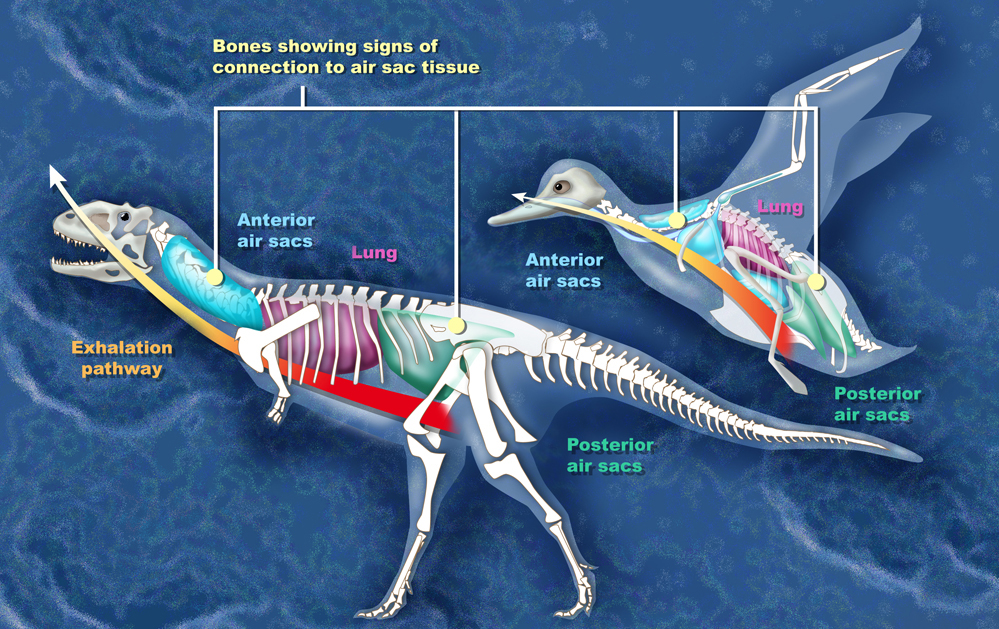
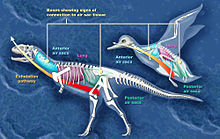 Comparación entre los sacos aéreos de Majungasaurus y un ave
Comparación entre los sacos aéreos de Majungasaurus y un aveLos científicos han reconstruido el sistema respiratorio de Majungasaurus basándose en la serie magníficamente preservada de vértebras (UA 8678) recobrados de la Formación Maevarano. La mayor parte de estas vértebras y algunas de las costillas contuvieron las cavidades (forámenes neumáticos) que pudo ser resultado de la infiltración de de sacos aéreos, similares a los del sistema respiratorio de las aves. En las aves, las vértebras del cuello y las costillas están ahuecadas por sacos de aire cervical, las vértebras de la espalda se conectan directamente con el pulmón, y las del lomo y sacras por los sacos de aires abdominales. Similares características de las vértebras del Majungasaurus implicarían la presencia de sacos aéreos. Estos sacos aéreos pudieron haber permitido una forma básica del estilo de respiración 'ventilación por flujo' donde el camino del aire que atraviesa los pulmones es unidireccional, de modo que el aire inhalado rico en oxígeno nunca se mezcla con el aire exhalado cargado con dióxido de carbono. Este método de respiración es muy complejo, pero a la vez altamente eficiente.[21]
El reconocimiento de forámenes respiratorios en Majungasaurus, además de proporcionar una comprensión de su biología respiratoria, también tiene implicaciones a mayor escala para la biología evolutiva. La separación entre la línea de los ceratosaurianos, la cuál llevó a Majungasaurus, y la línea de los tetanuros, a la que pertenecen las aves, habría ocurrido muy temprano en la historia de los terópodos. El sistema respiratorio aviar, presente en ambas líneas, debe por lo tanto haberse desarrollado antes de la división, y mucho antes de la evolución de las mismas aves. Esto proporciona evidencia adicional al origen dinosauriano de las aves.[21]
Cerebro y oído interno
Tomografía computada, del cráneo completo de Majungasaurus (FMNH PR 2100) permitió una reconstrucción grosera de su cerebro y la estructura del oído interno. Superficialmente, el cerebro era pequeño con respecto al tamaño del cuerpo, pero de de manera similar a muchos otros terópodos no celurosaurianos, con una forma muy conservadora más cercana a las de los cocodrilianos modernos que a los pájaros. Una diferencia entre Majungasaurus y otros terópodos es el pequeño flóculo, una región del cerebelo que ayuda a coordinar los movimientos de los ojos con los de la cabeza. Esto sugiere que Majungasaurus y otros abelisáuridos como Indosaurus, que también tendrían un pequeño flóculo, no confiaban en los movimientos principales rápidos de la cabeza para ver y para capturar su presa.[2]
Las inferencias sobre comportamiento se pueden hacer también del examen del oído interno. Los canales semicirculares del oído interno ayudaban en el equilibrio, y el canal semicircular lateral que es generalmente paralelo a la tierra cuando el animal tiene su cabeza en una postura de la alarma. Cuando la cabeza del Majungasaurus se gira de modo que su canal lateral sea paralelo a la tierra, la cabeza estaba completamente cerca de la horizontal. Esto contrasta con la mayoría de los otros terópodos, donde la cabeza estaba más fuertemente inclinada hacia el suelo cuando estaba en posición de alerta. El canal lateral era significativamente más largo wn Majungasaurus que en su pariente más basal Ceratosaurus, indicación de una mayor sensibilidad a los movimientos de lado a lado de la cabeza.[2]
Patologías
Un reporte de 2007 describe patologías en los huesos deMajungasaurus. Los científicos examinaron los restos por lo menos de 21 individuos y descubrieron cuatro con patologías claramente.[22] Mientras que las patologías se habían estudiado en grandes tetanuros como alosáuridos y tiranosáuridos,[23] esta es la primera vez que son estudiadas en un abelisáurido. No se encontró ninguna herida en elementos del cráneo, en contraste con los tiranosáuridos donde son comunes las mordeduras faciales a veces espantosas. Uno de los especimenes posee una de las falanges del pie, que aparentemente se habría quebrado y postriormente sanado.[22]
La mayoría de las lesiones de ven en las vértebras. Por ejemplo, una vértebra dorsal de un animal juvenil mostró exostosis en su superficie inferior. Este crecimiento probablemente se origino por la osificación de un cartílago o un tendón durante el desarrollo, pero la causa de la osificación no está determinada. La hipervitaminosis A y espondilosis son causas más probables, y el osteoma (tumor benigno de hueso) es posible pero muy raro. Otro espécimen, una pequeña vértebra caudal, también fue encontrado con un crecimiento anormal, este vez en el extremo de su proceso espinoso, que proyecta hacia dorsal de las vértebras, permitiendo la incerción del músculo[22] Crecimientos similares han sido encontrados en vértebras de Allosaurus[24] y Masiakasaurus, probablemente por la osificación del ligamento que corre por los los procesos espinosos (ligamento interespinal) a a los largo de sus extremos (ligamento supraespinal).[22]
La patología más seria descubierta estaba en una serie de cinco vértebras grandes de la cola. Las primeras dos vértebras mostraron solamente anormalidades de menor importancia a excepción de un surco grande que se extendía a lo largo del lado izquierdo de ambos huesos. Sin embargo, las tres vértebras siguientes estaban fundidas totalmente juntas en muchos diversos puntos, formando una masa de huesudo sólido. No hay rastros de ninguna otra vértebra después de la quinta de la serie, indicando que la cola terminó allí prematuramente. Del tamaño de las últimas vértebras, los científicos juzgaron que cerca de diez vértebras habían sido perdidas. Una explicación para esta patología es un severo traumatismo dando por resultado la pérdida de la extremidad de la cola, seguida por una osteomielitis (infección) de las vértebras ultimas restantes. Alternativamente, la infección pudo haber acaecido primero y haber llevado al final de la cola en convertirse en tejido necrótico y cayéndose. Éste es el primer ejemplo del truncamiento de la cola conocido en un dinosaurio de un terópodo no aviar.[22]
Paleoecología
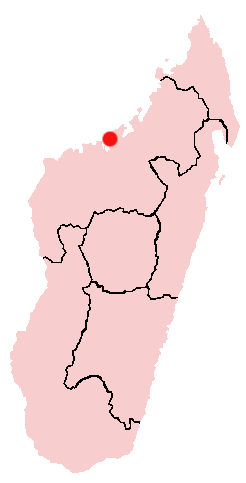
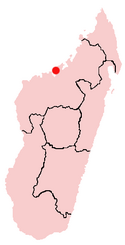 Todos los fósiles de Majungasaurus han sido encontrados en la Provincia de Mahajanga de Madagascar, a unos 50 kilometros al suroeste de la capital provincial de Mahajanga (marcada con un punto rojo en el mapa).
Todos los fósiles de Majungasaurus han sido encontrados en la Provincia de Mahajanga de Madagascar, a unos 50 kilometros al suroeste de la capital provincial de Mahajanga (marcada con un punto rojo en el mapa).Todos los especimenes de Majungasaurus han sido recobrados de la Formación Maevarano en la Provincia de Mahajanga noroeste de Madagascar. La mayoria de estos, incluyendo los más completos, provienen del Miembro Anembalemba de dicha formación, aunque dientes de Majungasaurus se han encontrado en la base de el Miembro Masorobe y en la superficie del Miembro Miadana. Mientras que estos sedimentos no se han fechado radiometricamente, la evidencia de bioestratigrafía y paleomagnetismo sugiere que fueran depositados durante Mastrichtiano, que duró entre los 70 a 65 millónes de años. Los dientes de Majungasaurus se encuentran justo encima del final del Mastrichtiano, cuando todos los dinosaurios no-aviares se extinguieron. [25]
Entonces como ahora, Madagascar era una isla, que se había separado a partir del Subcontinente indio hacia menos de 20 millones de años. Estaba viajando hacia el norte pero todavía se encontraba entre 10a 15 grados más al sur de lo que esta hoy. El clima prevaleciente era semiárido, con una fuerte estacionalidad en el régimen de lluvias y la temperatura. Majungasaurus habitó una costera llanura aluvional cortada por muchos río de lecho arenoso.[25] La fuerte evidencia geológica sugiere la ocurrencia de periódicas inundaciones de lodo a través de estos canales al principio de la estación húmeda, enterrando animales muertos durante la estación seca precedente y facilitando su preservación excepcional como fósiles.[26] Los niveles del mar en el área se elevaron a través del Mastrichtiano, y continuarían haciéndolo en el Paleoceno, por lo que Majungasaurus puede haber en vagado ambientes costeros como marismas también. La vecina Formación Berivotra representa el contemporáneo ambiente marino.[25]
Junto a Majungasaurus, los taxónes fósiles de Maevarano incluyen peces, ranas, lagartos, serpientes,[25] siete especies distintas de cocodrilomorfos,[27] cinco o seis de mamíferos,[27] y varias de aves,[25] entre ellas Vorona[28] , así como el posible dromeosáurido volador Rahonavis,[29] [30] en noasáurido Masiakasaurus[31] y dos saurópodos titanosaurianos, incluyendo el Rapetosaurus.[32] Majungasaurus eran por mucho el carnívoro más grande y probablemente el depredador dominante en tierra, aunque grandes cocodrilomorfos como Mahajangasuchus y Trematochampsa pudo ser competencia a la orilla del agua.[25]
Referencias
- ↑ a b c d e f g h i j Krause, David W.; Sampson, Scott D.; Carrano, Matthew T.; & O'Connor, Patrick M. (2007). «Overview of the history of discovery, taxonomy, phylogeny, and biogeography of Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar». En Sampson, Scott D.; & Krause, David W. (eds.). Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar. Society of Vertebrate Paleontology Memoir 8. pp. 1–20. http://www.bioone.org/perlserv/?request=get-abstract&doi=10.1671%2F0272-4634%282007%2927%5B1%3AOOTHOD%5D2.0.CO%3B2.
- ↑ a b c d e f g h i Sampson, Scott D.; & Witmer, Lawrence M. (2007). «Cranofacial anatomy of Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar». En Sampson, Scott D.; & Krause, David W. (eds.). Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar. Society of Vertebrate Paleontology Memoir 8. pp. 32–102. http://www.bioone.org/perlserv/?request=get-abstract&doi=10.1671%2F0272-4634%282007%2927%5B32%3ACAOMCT%5D2.0.CO%3B2.
- ↑ Smith, Joshua B. (2007). «Dental morphology and variation in Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar». En Sampson, Scott D.; & Krause, David W. (eds.). Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar. Society of Vertebrate Paleontology Memoir 8. pp. 103–126. http://www.bioone.org/perlserv/?request=get-abstract&doi=10.1671%2F0272-4634%282007%2927%5B103%3ADMAVIM%5D2.0.CO%3B2.
- ↑ O'Connor, Patrick M. (2007). «The postcranial axial skeleton of Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar». En Sampson, Scott D.; & Krause, David W. (eds.). Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar. Society of Vertebrate Paleontology Memoir 8. pp. 127–162. http://www.bioone.org/perlserv/?request=get-abstract&doi=10.1671%2F0272-4634%282007%2927%5B127%3ATPASOM%5D2.0.CO%3B2.
- ↑ Carrano, Matthew T. (2007). «The appendicular skeleton of Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar». En Sampson, Scott D.; & Krause, David W. (eds.). Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar. Society of Vertebrate Paleontology Memoir 8. pp. 163–179. http://www.bioone.org/perlserv/?request=get-abstract&doi=10.1671%2F0272-4634%282007%2927%5B163%3ATASOMC%5D2.0.CO%3B2.
- ↑ Depéret, Charles. (1896). «Note sur les Dinosauriens Sauropodes et Théropodes du Crétacé supérieur de Madagascar» (en francés). Bulletin de la Société Géologique de France 21: pp. 176–194.
- ↑ Depéret, Charles; & Savornin, Justin. (1928). «La faune de Reptiles et de Poissons albiens de Timimoun (Sahara algérien)». Bulletin de la Societé Géologique de France 27: pp. 257–265.
- ↑ Lavocat, René (1955). «Sur une portion de mandibule de Théropode provenant du Crétacé supérieur de Madagascar». Bulletin du Muséum National d’Histoire Naturelle 27: pp. 256–259.
- ↑ Sues, Hans-Dieter; & Taquet, Phillipe. (1979). «A pachycephalosaurid dinosaur from Madagascar and a Laurasia−Gondwanaland connection in the Cretaceous». Nature 279 (5714): pp. 633–635. doi:.
- ↑ Sampson, Scott D.; Krause, David W.; Dodson, Peter; & Forster, Catherine A. (1996). «The premaxilla of Majungasaurus (Dinosauria: Theropoda), with implications for Gondwanan paleobiogeography». Journal of Vertebrate Paleontology 16 (4): pp. 601–605. http://www.vertpaleo.org/publications/jvp/contents-16-4.cfm.
- ↑ Sampson, Scott D.; Witmer, Lawrence M.; Forster, Catherine A.; Krause, David W.; O'Connor, Patrick M.; Dodson, Peter; & Ravoavy, Florent. (1998). «Predatory dinosaur remains from Madagascar: implications for the Cretaceous biogeography of Gondwana». Science 280 (5346): pp. 1048–1081. doi:.
- ↑ Sampson, Scott D.; & Krause, David W. (eds.). (2007). Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar. Society of Vertebrate Paleontology Memoir 8. p. 184pp. http://www.bioone.org/perlserv/?request=get-toc&issn=0272-4634&volumen=27&issue=sp8.
- ↑ a b Sereno, Paul C. (2007). «New dinosaurs link southern landmasses in the Mid-Cretaceous». Proceedings of the Royal Society of London B: Biological Sciences 271 (1546): pp. 1325–1330. doi:.
- ↑ a b c Tykoski, Ronald B.; & Rowe, Timothy. (2004). «Ceratosauria». En Weishampel, David B.; Dodson, Peter; & Osmólska, Halszka (eds.). The Dinosauria (Second Edition edición). Berkeley: University of California Press. pp. 47–70. ISBN 0-520-24209-2.
- ↑ Coria, Rodolfo A.; Chiappe, Luis M.; & Dingus, Lowell. (2002). «A new close relative of Carnotaurus sastrei Bonaparte 1985 (Theropoda: Abelisauridae) from the Late Cretaceous of Patagonia» (subscription required). Journal of Vertebrate Paleontology 22 (2): pp. 460–465. doi:. http://www.bioone.org/perlserv/?request=get-document&doi=10.1671%2F0272-4634%282002%29022%5B0460%3AANCROC%5D2.0.CO%3B2.
- ↑ Rayfield, Emily; Norman, David B.; Horner, Celeste C.; Horner, John R.; Smith, Paula M.; Thomason, Jeffrey J.; & Upchurch, Paul. (2001). «Cranial design and function in a large theropod dinosaur». Nature 409 (6823): pp. 1033–1037. doi:.
- ↑ Van Valkenburgh, Blaire; & Molnar, Ralph E. (2002). «Dinosaurian and mammalian predators compared». Paleobiology 28 (4): pp. 527–543. doi:. http://paleobiol.geoscienceworld.org/cgi/content/abstract/28/4/527.
- ↑ a b c Rogers, Raymond R.; Krause, David W.; & Curry Rogers, Kristina. (2007). «Cannibalism in the Madagascan dinosaur Majungatholus atopus». Nature 422 (6931): pp. 515–518. doi:.
- ↑ Nesbitt, Sterling J.; Turner, Alan H.; Erickson, Gregory M.; & Norell, Mark A. (2006). «Prey choice and cannibalistic behavior in the theropod Coelophysis». Biology Letters 2 (4): pp. 611–614. doi:.
- ↑ a b Roach, Brian T.; & Brinkman, Daniel T. (2007). «A reevaluation of cooperative pack hunting and gregariousness in Deinonychus antirrhopus and other non-avian theropod dinosaurs». Bulletin of the Peabody Museum of Natural History 48 (1): pp. 103–138. doi:. http://web.archive.org/web/20070810172000/http://www.peabody.yale.edu/scipubs/abstracts/abs_b48-1b.html.
- ↑ a b O'Connor, Patrick M.; & Claessens, Leon P.A.M. (2006). «Basic avian pulmonary design and flow-through ventilation in non-avian theropod dinosaurs». Nature 436 (7048): pp. 253–256. doi:.
- ↑ a b c d e Farke, Andrew A.; & O'Connor, Patrick M. (2007). «Pathology in Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar». En Sampson, Scott D.; & Krause, David W. (eds.). Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar. Society of Vertebrate Paleontology Memoir 8. pp. 180–184. http://www.bioone.org/perlserv/?request=get-abstract&doi=10.1671%2F0272-4634%282007%2927%5B180%3APIMCTA%5D2.0.CO%3B2.
- ↑ Rothschild, Bruce; & Tanke, Darren H. (2005). «Theropod paleopathology: state-of-the-art review». En Carpenter, K. (ed.). The Carnivorous Dinosaurs. Bloomington: Indiana University Press. pp. 351–365. ISBN 978-0253345394.
- ↑ Hanna, Rebecca R. (2002). «Multiple injury and infection in a sub-adult theropod dinosaur Allosaurus fragilis with comparisons to allosaur pathology in the Cleveland-Lloyd Dinosaur Quarry collection». Journal of Vertebrate Paleontology 22 (1): pp. 76–90. doi:. http://www.vertpaleo.org/publications/jvp/22-076-090.cfm.
- ↑ a b c d e f Rogers, Raymond R.; Krause, David W.; Curry Rogers, Kristina; Rasoamiaramanana, Armand H.; & Rahantarisoa, Lydia. (2007). «Paleoenvironment and Paleoecology of Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar». En Sampson, Scott D.; & Krause, David W. (eds.). Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar. Society of Vertebrate Paleontology Memoir 8. pp. 21–31. http://www.bioone.org/perlserv/?request=get-abstract&doi=10.1671%2F0272-4634%282007%2927%5B21%3APAPOMC%5D2.0.CO%3B2.
- ↑ Rogers, Raymond R. (2005). «Fine-grained debris flows and extraordinary vertebrate burials in the Late Cretaceous of Madagascar». Geology 33 (4): pp. 297–300. doi:.
- ↑ a b Krause, David W.; O'Connor, Patrick M.; Curry Rogers, Kristina; Sampson, Scott D.; Buckley, Gregory A.; & Rogers, Raymond R. (2006). «Late Cretaceous terrestrial vertebrates from Madagascar: implications for Latin American biogeography». Annals of the Missouri Botanical Gardens 93 (2): pp. 178–208. doi:. http://www.mbgpress.info/index.php?task=id&id=11002.
- ↑ Forster, Catherine A.; Chiappe, Luis M.; Krause, David W.; & Sampson, Scott D. (1996). «The first Cretaceous bird from Madagascar». Nature 382 (6591): pp. 532–534. doi:.
- ↑ Forster, Catherine; Sampson, Scott D.; Chiappe, Luis M.; & Krause, David W. (1998). «The theropod ancestry of birds: new evidence from the Late Cretaceous of Madagascar». Science 279 (5358): pp. 1915–1919. doi:.
- ↑ Makovicky, Peter J.; Apesteguía, Sebastian; & Agnolín, Federico L. (2005). «The earliest dromaeosaurid theropod from South America». Nature 437 (7061): pp. 1007–1011. doi:.
- ↑ Sampson, Scott D.; Carrano, Matthew T.; & Forster, Catherine A. (2001). «A bizarre predatory dinosaur from the Late Cretaceous of Madagascar». Nature 409 (6819): pp. 504–506. doi:.
- ↑ Curry Rogers, Kristina; & Forster, Catherine A. (2001). «The last of the dinosaur titans: a new sauropod from Madagascar». Nature 412 (6846): pp. 530–534. doi:.
Enlaces externos
 Portal:Dinosaurios. Contenido relacionado con Dinosaurios.
Portal:Dinosaurios. Contenido relacionado con Dinosaurios. Wikimedia Commons alberga contenido multimedia sobre Majungasaurus. Commons
Wikimedia Commons alberga contenido multimedia sobre Majungasaurus. Commons- Majungasaurus en Dinodata (en inglés, se requiere registrarse en forma gratuita)
- Majungasaurus dentro de Abelisauridae en The Thescelsosaurus (en inglés)
- Significado y pronunciación del nombre (en inglés)
- Majungasaurus en The Theropod Database (en inglés)
Categorías:- Fósiles
- Abelisauroidea
- Dinosaurios del Cretácico
- Dinosaurios de la India y Madagascar
- M. crenatissimus (tipo)




Wikimedia foundation. 2010.